
Abstract
Intratympanic gentamicin (ITG) has been used to treat refractory Ménière’s disease. Disequilibrium after ITG was still a challenge for some patients, and the underlying mechanism is poorly understood. Our previous study demonstrated that gentamicin distributed in the bilateral vestibular efferent neurons (VEN) after ITG; however, does it lead to VEN damage and cause further disequilibrium in patients following ITG? In this study, we observed severe damaged gentamicin-positive neurons of VEN and severe fractured myelin layer plates around neural fibers when viewed under transmission electron microscopy at day 3 after ITG. At day 30, neurons of VEN presented with relatively normal structures. Compared with the control group, the total number of choline acetyltransferase (CHAT) immunolabeling neurons in bilateral VEN showed a significant decrease both at day 3 and day 30. However, there was no significant difference in the total number of CHAT immunolabeling neurons between day 3 and day 30. It indicates that gentamicin is not only retrogradely transported into bilateral VEN, but also results in the degeneration of VEN after ITG. These findings may be related to patients’ disequilibrium symptom after ITG. Furthermore, we speculate that VEN may play a role in vestibular compensation.
Introduction
Intractable Ménière’s disease (MD), characterized by spontaneous attacks of vertigo, fluctuating sensorineural hearing loss, tinnitus, and aural fullness, has plagued otolaryngologists for years. Since the intratympanic application of aminoglycoside was first reported in the treatment of refractory MD by Schuknecht, 1 intratympanic low or high doses of gentamicin injection gradually became a feasible and effective clinical strategy in vertigo control. In recent years, the reported vertigo control rate ranged from 80.7% to 89.5% using intratympanic gentamicin (ITG).2–5 However, the potential adverse effects induced by cochleotoxicity and vestibulotoxicity of gentamicin were also noticed in clinical application. Except for hearing loss, it was reported that 63% of patients with MD complained of a variety of disequilibrium after being treated with high-dose ITG. 6 Hillman et al. demonstrated that the median time of disequilibrium lasted up to 4 months. 7
Hair cells in vestibular end-organs, rather than non-sensory cells, showed a primary distribution of gentamicin when ITG was applied. 8 Furthermore, significant vestibular neuroepithelial damages were reported after administering ITG in animal studies.9–11 The mechanism of vertigo control following ITG in MD patients is that gentamicin damages vestibular neuroepithelium and decreases vestibular function to some extent. Therefore, we assumed that the post-treatment disequilibrium may be related to the unilateral peripheral vestibular damage. Compared with ITG, which induces the incomplete decrease of peripheral vestibular function, the vestibular neurotomy leads to complete vestibular damage. However, it is reported that a similar rate and severity of post-treatment disequilibrium was identified in patients with either vestibular nerve section or with ITG. 7 Unilateral peripheral vestibular damage may not fully explain the mechanism of disequilibrium after ITG, especially the persistent disequilibrium. We then hypothesized that the central vestibular degeneration following ITG may also contribute to the persistent disequilibrium.
Our previous study demonstrated that intense gentamicin immunofluorescence accumulated in the bilateral vestibular efferent neurons (VEN) for at least 1 month; however, no gentamicin was identified in the neurons of vestibular nuclear complex. 12 It suggested that gentamicin might serve as a retrograde tracer traveling from the vestibular efferent terminals to VEN along vestibular efferent fibers. Although it is clear that the application of ITG leads to damages of vestibular hair cells and nerve endings,9,10,13,14 how gentamicin impacts VEN remains unclear.
In this study, we applied transmission electron microscopy (TEM) and fluorescent IHC with antibodies to choline acetyltransferase (CHAT) to determine the short-term and long-term effects of gentamicin on VEN qualitatively and quantitatively.
Materials and Methods
Animals
A total of 30 adult male and female albino guinea pigs weighing between 250 g and 300 g were used. The animals were divided into 6 groups (Table 1), including group a1, a2, and a3 (4 animals in each group) for TEM observation, and group b1, b2, and b3 (6 animals in each group) for immunofluorescence analysis. Animals in group a1 and b1 underwent intratympanic saline administration and were sacrificed at day 3 as the control group. Animals in group a2 and b2 were sacrificed at day 3 following intratympanic gentamicin injection of 60 mg/ml. Animals in group a3 and b3 were sacrificed at day 30 following ITG injection of 60 mg/ml. Animals in group a1 and a3 were sacrificed for TEM observation following retrograde tracing.
The Particular Information of Each Group.
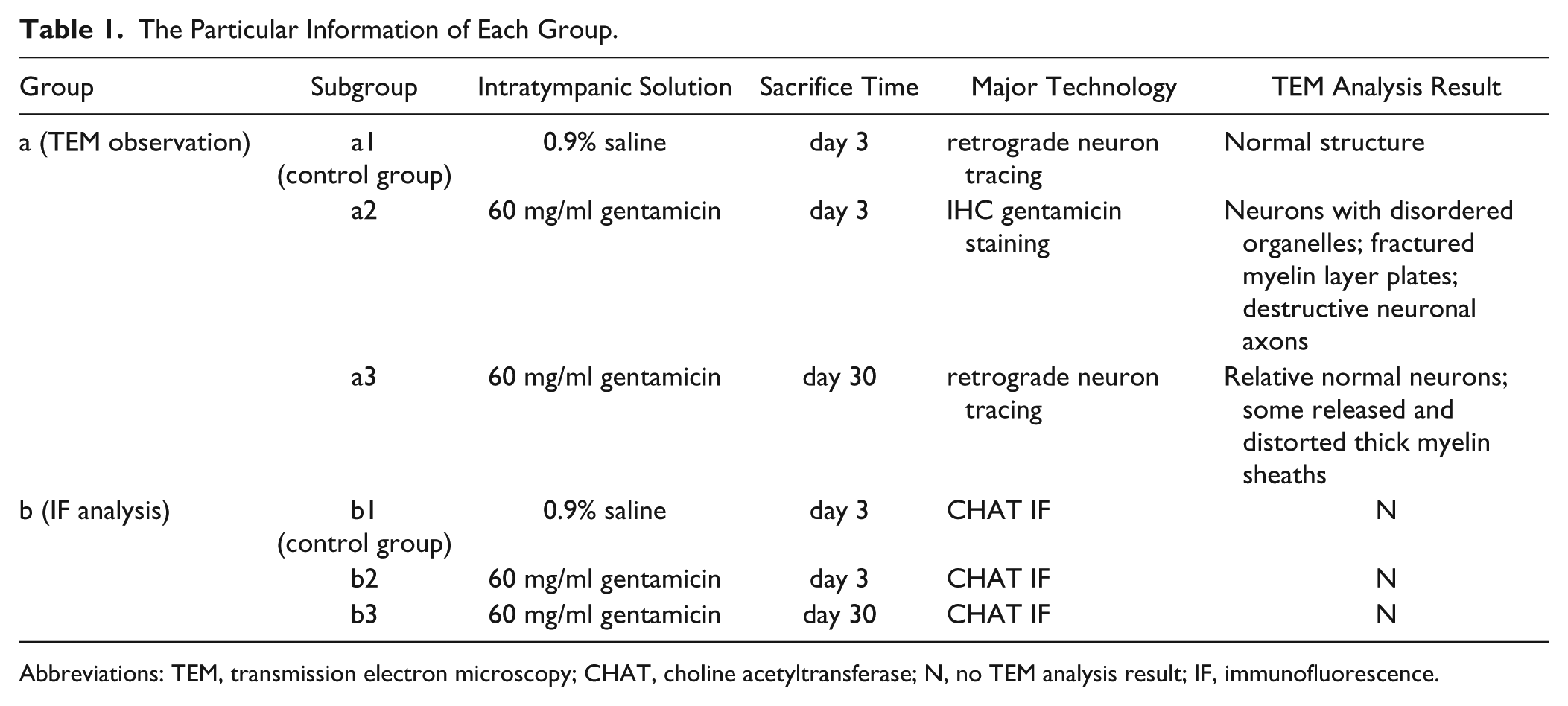
Abbreviations: TEM, transmission electron microscopy; CHAT, choline acetyltransferase; N, no TEM analysis result; IF, immunofluorescence.
All experimental protocols followed the guidelines of the Ethical Board of Eye & Ear Nose and Throat Hospital, Fudan University, and were approved by the Committee of Care and Use of Animals, Shanghai, China.
ITG Administration
All the animals, without visible otitis media, were anesthetized with intramuscular ketamine (40 mg/kg) and xylazine (10 mg/kg). The external auditory canal was sterilized with 75% ethanol solution. Gentamicin (product # G3632; Sigma Co.; Saint Louis, MO) was diluted into a concentration of 60 mg/ml by dissolution in 0.9% saline. The gentamicin solution (100–120 µl) was injected into the tympanic cavity until the middle-ear space was full after puncturing the tympanic membrane using a 100 µl Hamilton microsyringe. The animal’s head was held stationary with the treated ear and nose turned toward the ceiling to ensure that the injected solution contacted with the round window and the oval window for at least 20 min. For the control group, 0.9% saline solution was administered following the same protocol. The right ear of all animals was defined to be the treated ear.
Tissue Preparation and Immunohistochemical Gentamicin Staining
Animals in group a2 (used for TEM observation) were perfused at day 3 through the left ventricle transcardially with 250 ml of saline solution and 250 ml of fixative solution (4% paraformaldehyde and 0.125% glutaraldehyde in 0.1 M phosphate buffer, pH = 7.4) at room temperature. Then, the brain stem was removed and fixed overnight at 4C in the same fixative solution. Cross-sections (50 μm) containing VEN were obtained using a vibratome (Leica; Wetzlar, Germany).
After washing in 0.01 M PBS (pH = 7.2–7.4) for 10 min 3 times, the floating-tissue sections were blocked in 10% donkey serum for 1 hr at room temperature and then incubated in mouse anti-gentamicin monoclonal antibodies (1:500 in PBS; QED Biosciences; San Diego, CA) overnight at 4C. Afterward, they were incubated in goat anti-mouse IgG antibody, horseradish peroxidase (HRP) conjugate (1:500 in PBS; Sigma) for 30 min at room temperature and reacted with 0.025% 3,3′-diaminobenzidine (DAB) tetrahydrochloride reaction (Sigma) with 0.075% hydrogen peroxide for 5 to 10 min until the neurons labeled. Floating sections were moved into the fixative solution for sample preparation of TEM observation.
Retrograde Neuron Tracing
Animals in groups a1 and a3 (used for TEM observation) were fully anesthetized with the aforementioned protocol. They were placed in a right recumbent position. After being sterilized with 75% ethanol solution, a C-shaped retro-auricular skin incision was made. The muscles attached to the right bulla were removed to expose the bulla. Then, the bulla was opened to access the stapedial footplate and the oval window. The retrograde tracer, biotinylated dextran amine (BDA; 3000 MW; Molecular Probes; Eugene, OR), at a concentration of 10% in 0.01 M phosphate, was slowly injected into the right oval window with a Hamilton microsyringe (2 µl). The total amount of retrograde tracer used was 0.5 to 0.8 µl. Then, the oval window was blocked by muscles. The wound was treated with Aureomycin ointment after suturing to prevent infection.
After a survival period of 7 days, the animals were fully anesthetized and euthanized. The processes of tissue preparation were the same as mentioned previously. The tissue floating sections were incubated in Avidin-HRP (1:4000 in PBS; Sigma) overnight at 4C after washing in 0.01 M PBS for 10 min 3 times and then reacted with 0.025% DAB working solution with 0.075% hydrogen peroxide for 5–10 min until the neurons were present. Sections were removed into the fixative solution for sample preparation of TEM observation.
TEM
The floating sections were washed in double distilled water (ddH2O) for 10 min 3 times and fixed in 1% osmium tetroxide for 1 hr. Then, the sections were dehydrated in a series of ascending graded ethanol (50%, 70%, 90%, and 100%) for 5 min, respectively, and propylene epoxide for 10 min and then embedded in araldite and polymerized for 12 hr at 37C, 12 hr at 45C, and 48 hr at 60C. As the neurons of VEN had been stained, the targeted areas were removed from the sections. The ultrathin sections (100 μm) with the target neurons were obtained using an ultramicrotome (Leica) and stained with uranyl acetate and Sato lead stain. They were examined under the TEM (JEOL; Tokyo, Japan).
Tissue Preparation and CHAT Immunofluorescence
Animals in group b1, b2, and b3 (used for immunofluorescence analysis) were perfused through the left ventricle transcardially with 250 ml saline solution and fixative solution (4% paraformaldehyde in 0.1 M phosphate buffer, pH = 7.4) at room temperature. Then, the brain stem was removed and fixed overnight at 4C in 4% paraformaldehyde. Tissues were immersed in 20% sucrose solution for approximately 24 hr until they sank to the bottom at 4C, followed by 30% sucrose solution. Afterward, tissues were rapidly frozen with optimal cutting temperature (OCT; Leica) and cut into 30 μm cross-sections using a cryostat (Leica). Every 4 consecutive floating sections were put together.
After washing in 0.01 M PBS for 10 min 3 times, the floating-tissue sections were blocked in 10% donkey serum with 0.2% Triton X-100 for 1 hr at room temperature. Sections were incubated with goat anti-CHAT antibody (1:100 in 5% donkey serum with 0.1% Triton X-100; Merck Millipore; Darmstadt, Germany) for 24 hr at 4C. Then, they were secondarily labeled with Alexa Fluor-488-conjugated donkey anti-goat antibodies (1:1000 in PBS containing 5% donkey serum and 0.1% Triton X-100) for 1 hr at 37C. DAPI (4′,6-diamidino-2-phenylindole; 1:500 in PBS; Invitrogen, Waltham, MA) was used to label nuclei for 15 min at 37C. Washing steps in 0.01 M PBS for 10 min 3 times were performed between each incubating step. Sections were mounted on glass slides using mounting medium. Positive fluorescent neurons were observed and counted under confocal laser scanning microscopy (TCS SP5; Leica). In order to show the target neurons and the location of neurons, the high magnification images were spliced by using Adobe Illustrator CC 2017 and adjusted the brightness and contrast by using Photoshop CC 2017.
One floating-sectioned brain slice was selected from every 4 consecutive 30 µM sections containing the VEN. The total unbiased cells with CHAT-positive cell body and visible neuronal nuclei in VEN were counted within these selected sections.
Statistical Analysis
One-way ANOVA was used and followed by the least-significant difference (LSD) test to find the differences among group b1, b2, and b3. Comparison of cell counts between the regions of interest on either side of the midline for each group was performed using the paired-sample Student’s t-test. Statistics of positive neurons were analyzed using SPSS v16.0 (SPSS, Inc.; Chicago, IL). A p<0.05 was defined as statistically significant.
Results
Identification of VEN Under Light Microscope
VENs in group a2 were identified by immunostaining of gentamicin (Fig. 1), while VENs in group a1 and a3 were identified by immunostaining of BDA (Fig. 2). These VENs were mainly located in the region of lateral vestibular efferent neurons (L-VEN), which is dorsolateral to the genu of the facial nerve (g7), ventral and medial to the medial vestibular nucleus (MVN), and in close proximity to the subependymal granular layer of the fourth ventricle (4V).12,15–18 Neurons were not always labeled in the medial vestibular efferent neurons (M-VEN), which is ventral to the genu on either side. The neuronal cell bodies of VEN in group a2 (sacrificed at day 3 without retrograde tracing of BDA after ITG) were dramatically labeled with gentamicin (Fig. 1). Meanwhile, DAB particles labeling the retrograde tracer BDA in group a1 and group a3 (sacrificed at day 30 after ITG; Fig. 2) appeared not only in neuronal cell bodies but also in their axons and dendrites.
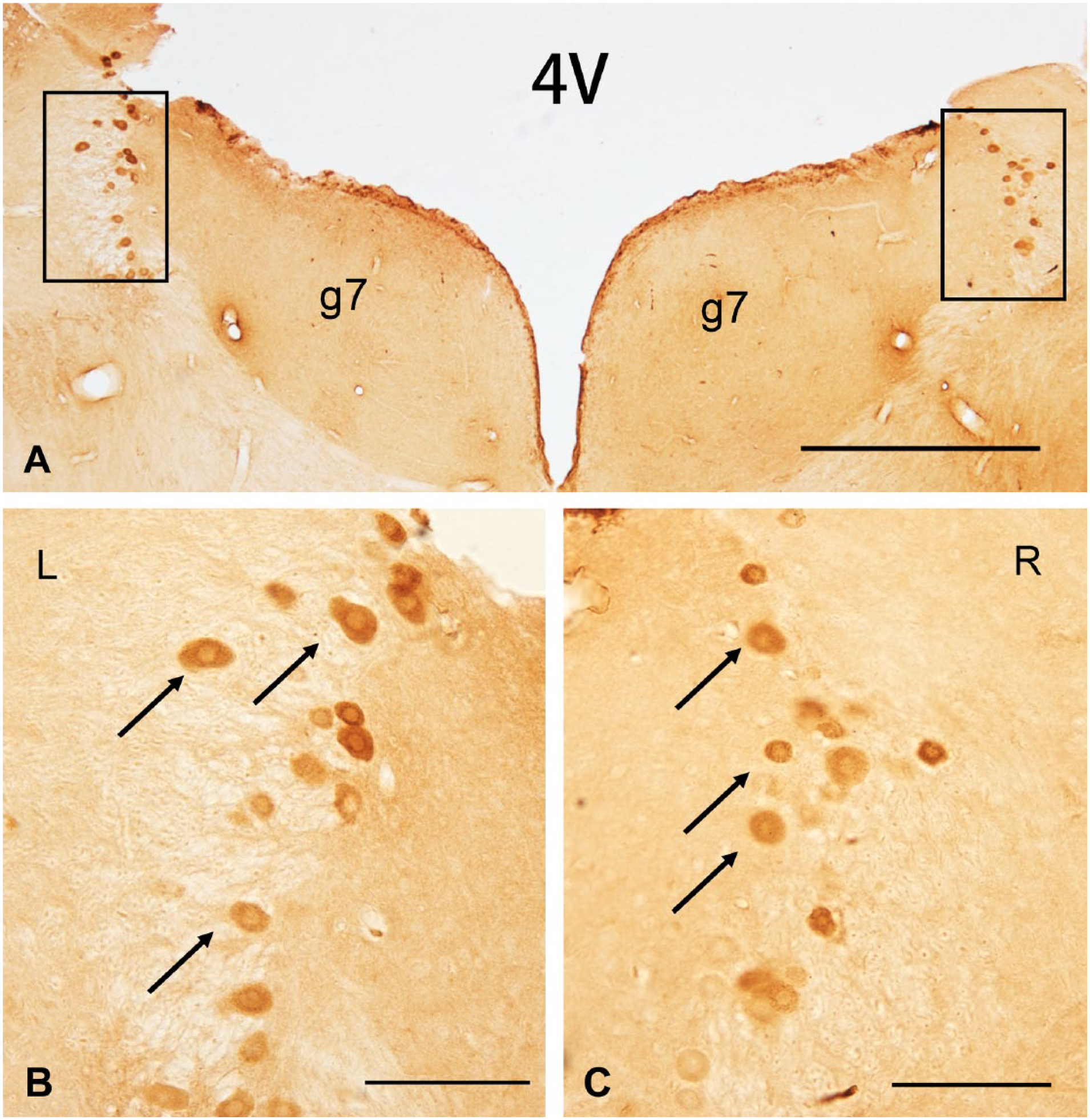
Cross-section of the brain stem stained with gentamicin at day 3 following ITG. Positive gentamicin immunostaining was presented in bilateral VEN and gentamicin particles diffusely distributed in the cytoplasm of neurons at day 3 following unilateral intratympanic injection of 60 mg/ml gentamicin. Arrows indicate examples of neuronal cell bodies. Scale bar = 500 μm in image A; 100 μm in image B and C. Abbreviations: 4V, the fourth ventricle; g7, the genu of facial nerve; ITG, intratympanic gentamicin; VEN, vestibular efferent neurons.
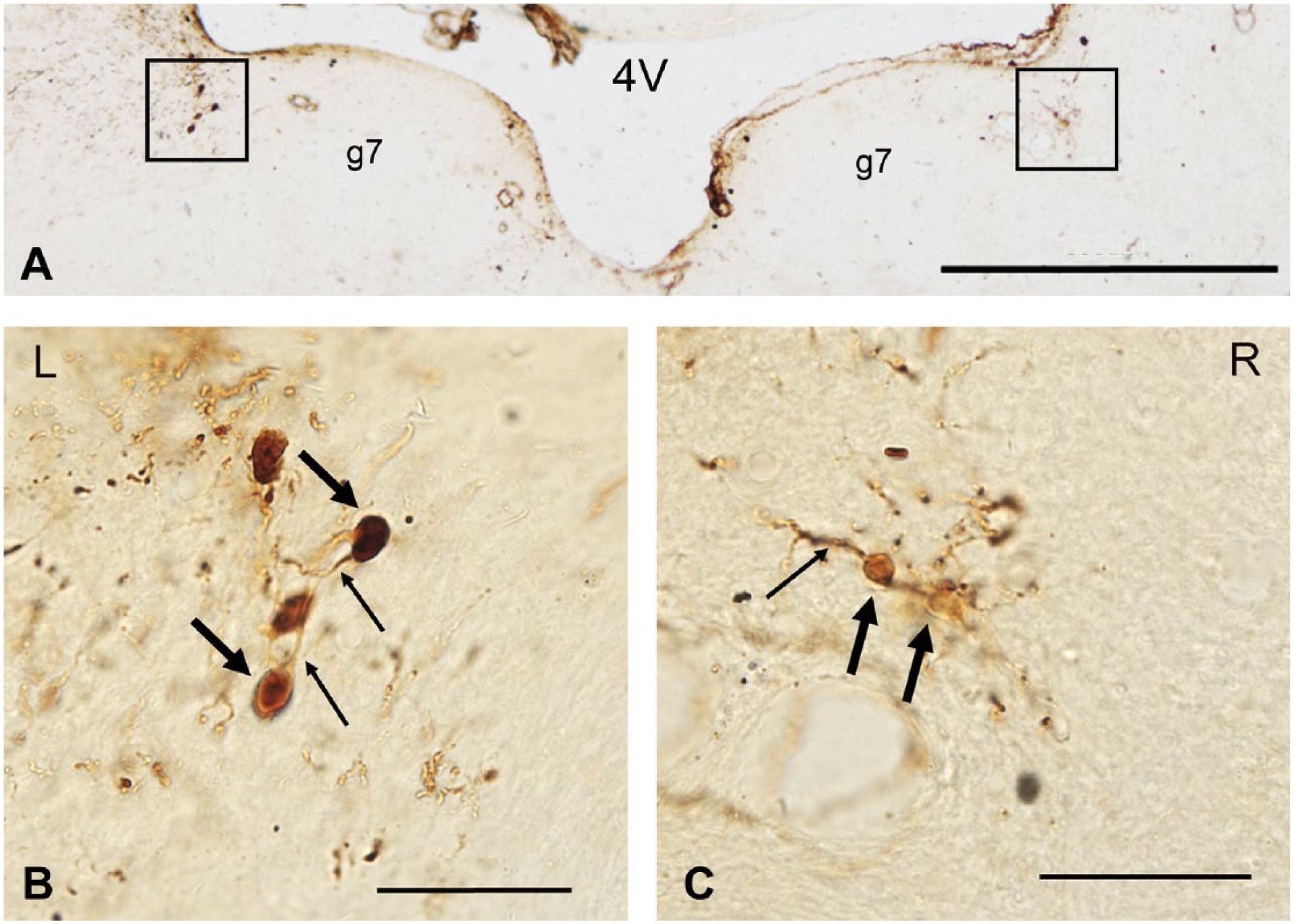
Cross-section of the brain stem stained with DAB labeling the retrograde tracer BDA at day 30 following ITG. BDA particles diffusely distributed in neuronal cell bodies, neuronal axons, and dendrites at day 30 following unilateral intratympanic injection of 60 mg/ml gentamicin. Thin arrows indicate examples of neuronal cell bodies. Thick arrows indicate examples of neuronal axons and dendrites. Scale bar = 500 μm in image A; 50 μm in image B and C. Abbreviations: 4V, the fourth ventricle; g7, the facial nerve; DAB, 3,3′-diaminobenzidine; BDA, biotinylated dextran amine; ITG, intratympanic gentamicin.
Ultrastructural Changes of VEN After ITG
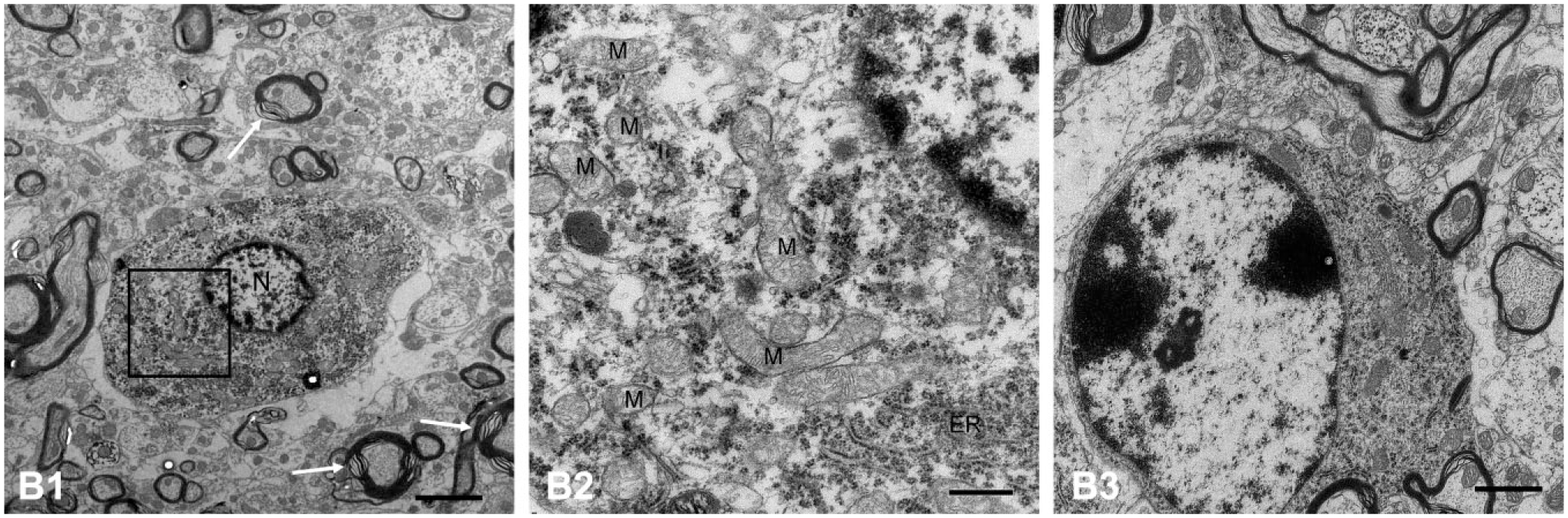
A series of ultrastructural degenerations were observed in the VEN after unilateral single ITG in the short and long term under electron microscopy. In group a2 (sacrificed at day 3 after ITG), the cytoplasm of neurons showed a diffused distribution of gentamicin-dense particles. The neurons exhibited disordered arrangement of organelles, swollen mitochondria (M), and dilated endoplasmic reticulum (ER; Fig. 3A2). The swollen mitochondria lost the matrix density, which indicated the outer membrane of mitochondria was damaged. In addition, the components of mitochondria, such as mitochondrial cristae, had disappeared and were replaced by gentamicin-dense particles (Fig. 3A2). The nerve fibers, especially those with thicker myelin layer plates around neurons, presented markedly fractured myelin layer plates and destructive neuronal axons. A large number of vacuolated nerve fibers was also detected (Fig. 3A1). Neuroglial cells presented with discontinuous plasma membranes, marginalized chromatin, disordered cytoskeletons, and scanty organelles (Fig. 3A3).
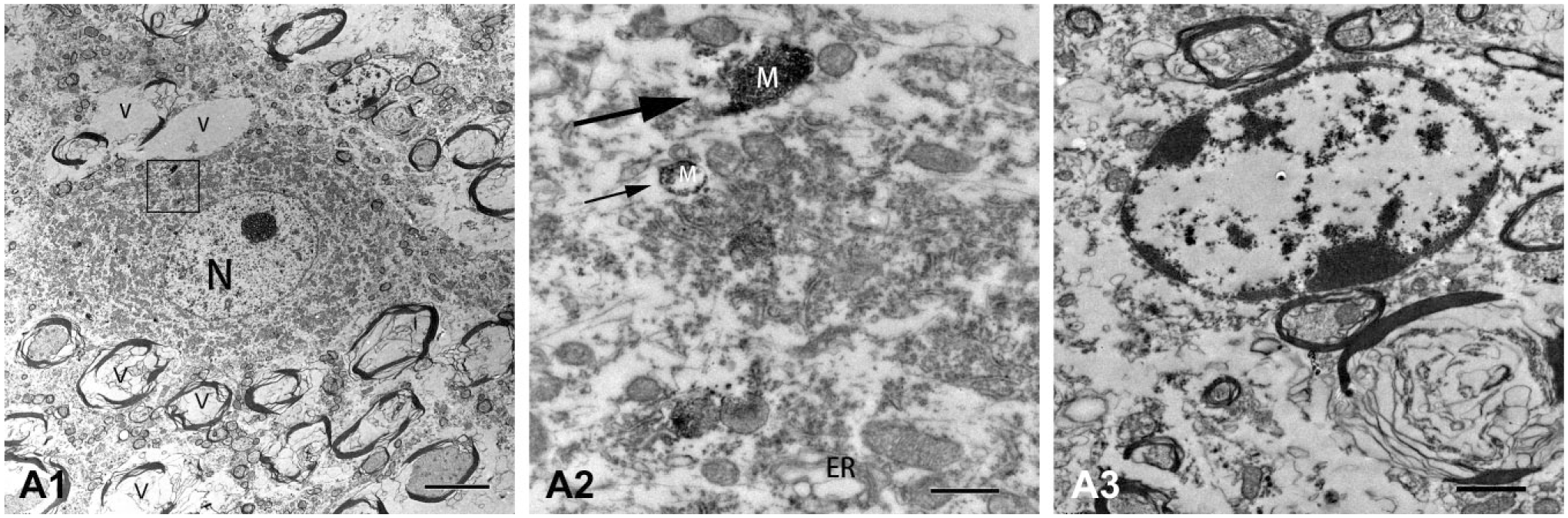
Electron micrographs of VEN in guinea pigs sacrificed at day 3 following ITG. (A1) A diffused distribution of gentamicin particles was presented in the cytoplasm of neurons at day 3 after unilateral intratympanic injection of 60 mg/ml gentamicin. The nerve fibers showed markedly fractured myelin layer plates and destructive neuronal axons. A large number of vacuolated nerve fibers (V) was also detected. (A2) The neurons exhibited disordered arrangement of organelles, swollen mitochondria (M; thin arrow), and dilated ER. In some mitochondria, mitochondrial cristae disappeared and were replaced by gentamicin particles (thick arrow). (A3) Neuroglial cells showed serious damage of discontinuous plasma membranes, disordered cytoskeletons, and scanty organelles. Scale bar = 5 μm in image A1; 0.5 μm in image A2; 1 μm in image A3. Abbreviations: VEN, vestibular efferent neuron; ITG, intratympanic gentamicin; ER, endoplasmic reticulum.
In group a3 (sacrificed at day 30 after ITG), neuronal cytoplasm was diffusedly filled with high electron–dense BDA particles (Fig. 4B1). However, these neurons of group a3 presented with nearly normal structures. Normal organelles (including normal mitochondria with cristae, well-developed ER, and lysosomes) were observed in neuronal cytoplasm (Fig. 4B2). Neuroglial cells were in a healthy condition with intact cell membranes, nuclei, and rich cytoplasmic organelles (Fig. 4B3). Although most myelin sheaths were normal with concentric myelin layer plates, myelin layer plates of some thick myelin sheaths were released and distorted (Fig. 4B1).
Electron micrographs of VEN in guinea pigs sacrificed at day 3 following ITG. (B1, B2) BDA particles were presented in the cytoplasm of neurons and were distributed densely around the membranes of cell nucleus (N) at day 30 after unilateral intratympanic injection of 60 mg/ml gentamicin. Most myelin sheaths were normal with concentric myelin layer plates. Myelin layer plates of some thick myelin sheaths were released and distorted (arrows). (B2) The neuron was relatively normal with normal mitochondria (M), and well-developed ER. (B3) Neuroglial cells exhibited a healthy condition with intact cell membranes, nuclei, and rich cytoplasmic organelles. Scale bar = 2 μm in image B1; 0.5 μm in image B2; 1 μm in image B3. Abbreviations: VEN, vestibular efferent neuron; BDA, biotinylated dextran amine; ITG, intratympanic gentamicin; ER, endoplasmic reticulum.
In the control group (group a1), the neurons presenting with high electron density of BDA particles were located diffusely in neuronal cytoplasm. The neurons and all components of the VEN were healthy. The neurons had large round nuclei, mitochondria with numerous cristae, well-developed ER and Golgi complexes, and lysosomes (Fig. 5C1). Myelin sheaths and neuroglial cells around the neurons were also healthy (Fig. 5C2).
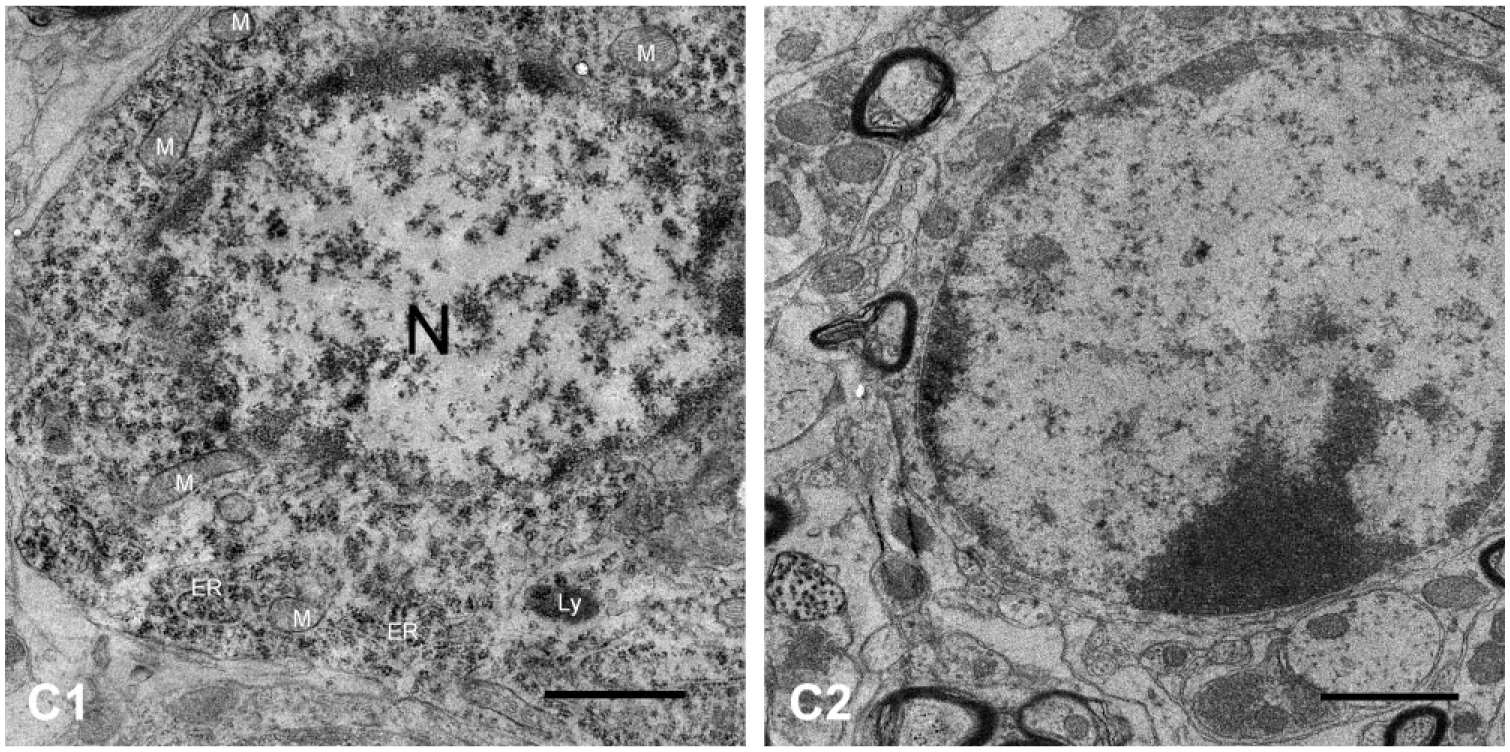
Electron micrographs of VEN in the control group. (C1) The neuron, which contained large round nuclei, mitochondria with numerous cristae, well-developed ER, and some Ly, presented a distribution of BDA particles in the neuronal cytoplasm and densely around the membranes of nucleus (N). (C2) Myelin sheaths and neuroglial cells were also healthy. Scale bar = 1 μm. Abbreviations: VEN, vestibular efferent neuron; ER, endoplasmic reticulum; Ly, lysosome; BDA, biotinylated dextran amine; ER, endoplasmic reticulum.
Identification of VEN With CHAT Immunofluorescence
We applied immunofluorescence with anti-CHAT antibody to identify the location of VEN. The CHAT-positive neurons, which were located in the region dorsolateral to g7, ventral and medial to MVN, and adjacent to the fourth ventricle, were counted and analyzed (Fig. 6).
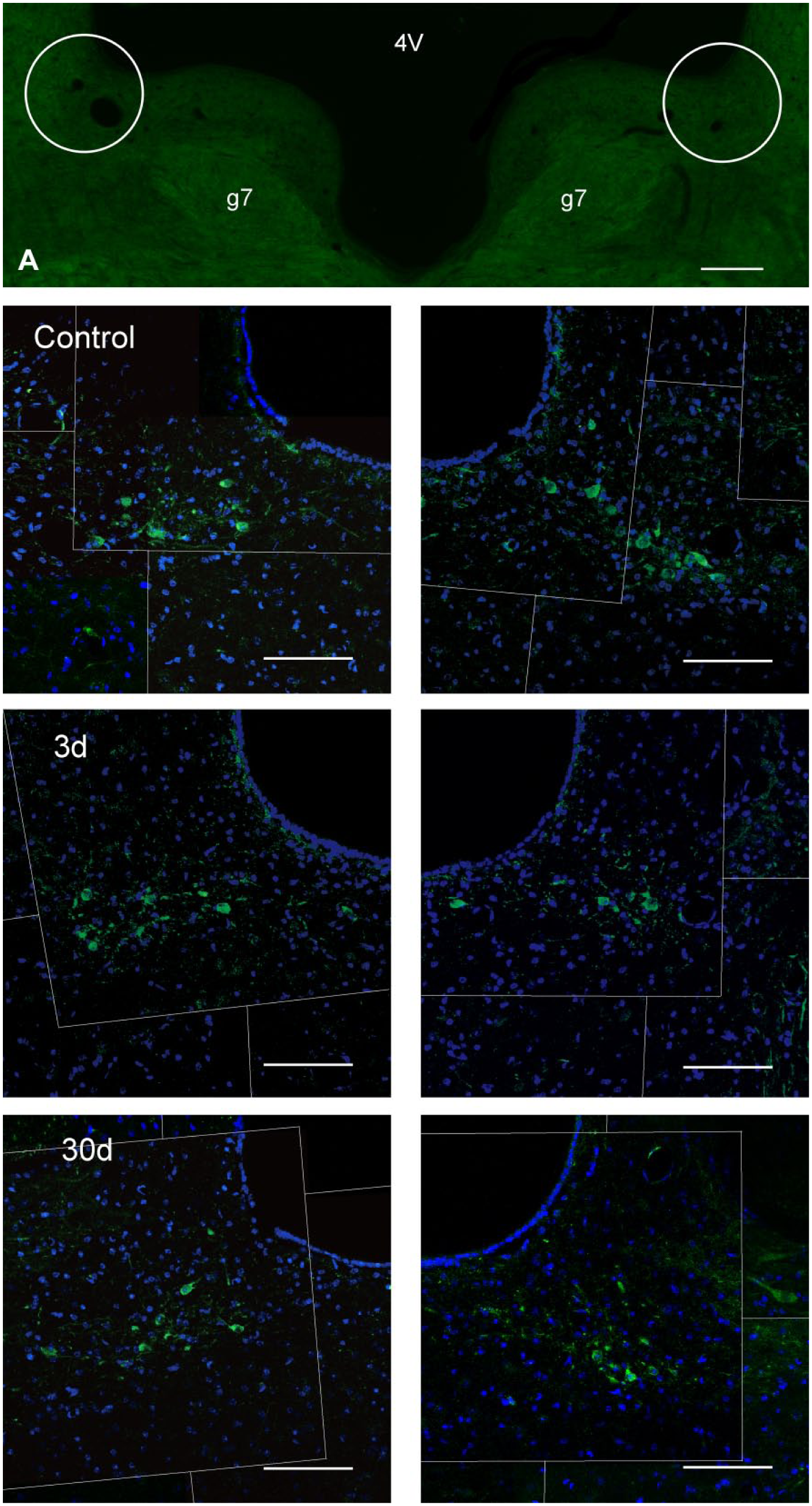
Distribution of CHAT-positive neurons (green) in bilateral L-VEN in different groups. (A) The location of bilateral L-VEN was presented in the region dorsolateral to g7 and adjacent to the fourth ventricle (4V). Control: Positive CHAT immunolabeling showed in bilateral L-VEN after intratympanic saline. 3d: Bilateral L-VEN were immunolabeled with CHAT at day 3 after unilateral intratympanic injection of 60 mg/ml gentamicin. 30d: Bilateral L-VEN were immunolabeled with CHAT at day 30 after unilateral intratympanic injection of 60 mg/ml gentamicin. Scale bar = 2 mm in image A; 100 μm in other images. Abbreviations: 4V, the fourth ventricle; g7, the facial nerve; 3d, at day 3; 30d, at day 30; CHAT, choline acetyltransferase; L-VEN, lateral vestibular efferent neuron.
Statistical Analysis of VEN Number After ITG
The mean number of CHAT-positive VEN in every group was calculated (Table 2). The total number of CHAT immunolabeling neurons in bilateral VEN showed a significant decrease at day 3 (group b2; p=0.000) and day 30 (group b3; p=0.000) following ITG compared with group b1 (control group) by using one-way ANOVA (F = 21.09, p=0.000) and LSD test (Fig. 7). However, no significant difference was observed in the total number of CHAT immunolabeling neurons in bilateral VEN between day 3 and day 30 (p=0.967). The same trend of the number of CHAT-positive neurons between day 3 and day 30 was also detected in the ipsilateral and contralateral VEN, respectively. There was no statistical difference in numbers of CHAT-positive neurons between the ipsilateral and contralateral VEN within control group, group b2, or group b3 by using the paired-sample Student’s t-test (p>0.05).
The Number of CHAT-positive Neurons in VEN.
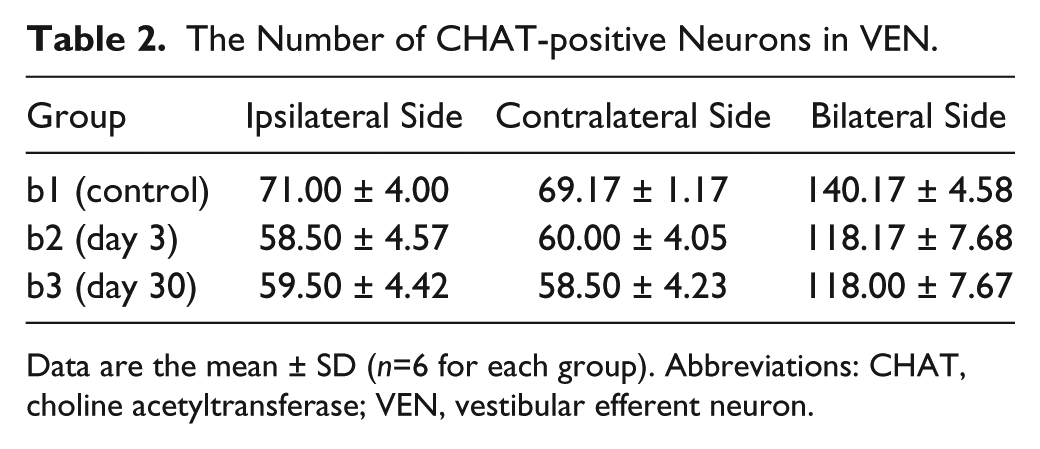
Data are the mean ± SD (n=6 for each group). Abbreviations: CHAT, choline acetyltransferase; VEN, vestibular efferent neuron.
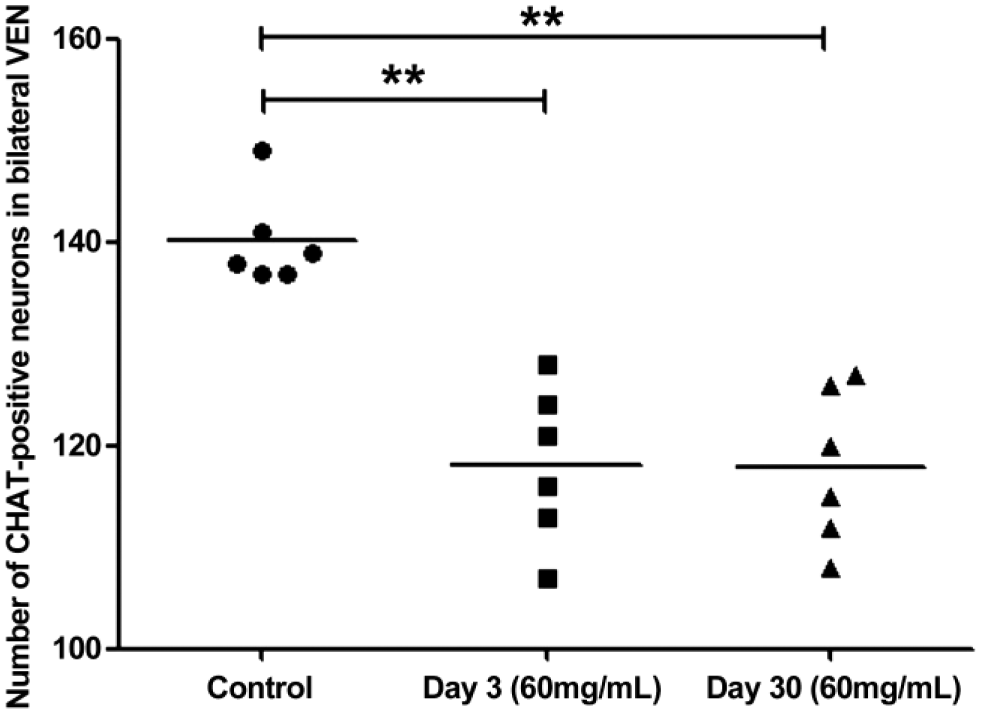
The number of CHAT-positive neurons in bilateral VEN at different time-points post-injection. Significantly decreased numbers of CHAT immunolabeling neurons were observed at day 3 (LSD: p<0.01) and day 30 (LSD: p<0.01) following unilateral intratympanic injection of 60 mg/ml gentamicin. No significant difference in number of CHAT immunolabeling neurons in VEN was observed between day 3 (group b2) and day 30 (group b3) following unilateral intratympanic injection of 60 mg/ml gentamicin (LSD: p>0.05). Abbreviations: CHAT, choline acetyltransferase; VEN, vestibular efferent neuron; LSD, least-significant difference. **p<0.01.
Discussion
In the present study, we explored the short- and long-term effects of gentamicin on VEN by using TEM observation and immunofluorescence analysis following unilateral intratympanic 60 mg/ml gentamicin injection. The results suggest that gentamicin could not only be retrogradely transported into bilateral vestibular efferent neuronal cytoplasm but also cause significant damages to the ultrastructure of VENs and a quantitative loss of the CHAT-positive neurons in the VEN in a short period of time after unilateral injection. Nerve fibers and neuroglial cells around the neurons also demonstrated destructive changes at day 3. However, the residual neurons in VEN showed a relatively normal condition at day 30 following ITG with an equal quantity of neurons compared with neurons at day 3. These quantitative and qualitative changes of the VEN following ITG in the short and long term may be associated with the persistent disequilibrium complication and may suggest some role of VEN in vestibular compensation.
In the group a2 (short term), the animals were sacrificed for TEM observation, and the location of DAB particles indicated where gentamicin was located. Intense gentamicin immunolabeling in the cytoplasm of bilateral VEN, which is consistent with the result in our previous study, 12 was observed at day 3 after unilateral ITG. Therefore, gentamicin served as a retrograde tracer to precisely identify VEN under light microscopy and then to determine the ultrastructural changes of the identified neurons under TEM. Although gentamicin-positive neurons could be observed in VEN by using immunofluorescent staining at day 30 after unilateral ITG in our previous study, 12 gentamicin-positive neurons could not be identified directly by immunohistochemical staining of gentamicin at day 30 after unilateral ITG because of the lower sensibility of DAB staining compared with immunofluorescence. Therefore, retrograde neuron tracing was applied in the long-term group. Bilateral VEN were successfully pinpointed by injecting the retrograde tracer BDA into the oval window. VEN consists of two groups. L-VEN is a larger group that is located dorsolateral to g7, ventral and medial to MVN, and adjacent to 4V; and M-VEN is located ventral to the genu. 18 Neurons in the L-VEN are much more prevalent than neurons in M-VEN and are easier identified by retrograde neuron tracing. In addition, the M-VEN ventral to the genu was reported to be negative to some markers, such as CHAT, acetylcholinesterase (AChE), and calcitonin gene-related peptide (CGRP). 19 CHAT is known as being expressed in VEN.15,20,21 In this study, antibody to CHAT was not only used to confirm the location of VEN but also to quantitatively analyze the neurons of VEN after unilateral ITG. Thus, we primarily focused on the L-VEN in this study.
The morphological changes and decreased number of VEN were demonstrated at day 3 after ITG. Two possible explanations need to be taken into account. One is de-efferentation induced by the indirect impact of the degeneration of vestibular neuroepithelium following ITG. However, previous studies reported that there was no loss of vestibular ganglion cells up to 12 weeks after labyrinthectomy.22,23 In addition, the impact of de-efferentation should not appear in such a short period of time. Another reason may be the direct impact of gentamicin on the neurons in VEN. The nerve fibers of VEN directly innervate to bilateral vestibular hair cells and synapse on type II hair cells, on bouton afferents innervating type II hair cells, and on calyx and dimorphic afferents of type I hair cells.24,25 Gentamicin may transport to the VEN promoted by damaging the vestibular hair cells and the nerve terminals. The mechanisms of how gentamicin induces the degeneration of neurons in VEN may be similar to that of vestibular hair cell degeneration following gentamicin accumulation. It is known that mitochondria are the source of energy and the mediators of cell death. Furthermore, it is reported that aminoglycoside antibiotics are ototoxic to sensory hair cells and cochlea nucleus by influencing mitochondrial energy metabolism and increasing reactive oxygen species (ROS) production, which affected mitochondria themselves, other organelles, and various pathways, and ultimately induced apoptosis.26–29 The electron micrographs showed that besides diffused distribution of gentamicin in the cytoplasm of the neurons, gentamicin was found markedly in swelling or completely damaged mitochondria at day 3 after ITG. Specific distribution of gentamicin in neurons and the correlated degeneration of neurons in VEN provided strong evidences to support the direct toxicity of gentamicin on the neurons in VEN. The neurotoxic effect of gentamicin on VEN may be originally caused by the direct toxicity on mitochondria in the neurons. Ultrastructural changes of the nerve fibers presenting markedly fractured myelin layer plates, destroyed neuronal axons, and vacuolation also may be associated with the direct toxicity of gentamicin.
At day 30 following ITG, we observed that neurons of VEN were in a relatively normal morphological ultrastructure under TEM. Furthermore, most myelin sheaths were healthy with concentric myelin layer plates. However, the count of neurons in VEN between day 3 and day 30 following ITG showed no significant difference. We hypothesize that some incompletely damaged neurons may gradually restore to relative normality. This restoration may be related to the self-recovery abilities of neurons and myelin sheath, as well as the support and assistance of surrounding neuroglial cells including astrocytes, oligodendrocytes, and microglias. 30 The ultrastructural recovery of neuroglial cells at day 30 after ITG suggested that they could also repair themselves or proliferate. However, neurons can only be repaired, but are not able to proliferate. Intratympanic high-dose gentamicin injection was applied in this study to confirm initially whether gentamicin could have an effect on neurons in VEN in the short term and long term. A follow-up study to compare effects of different doses of gentamicin on VEN should be conducted.
Although the number of neurons in VEN is small, a single efferent fiber arising from efferent neurons makes contact with major afferents and hair cells in the vestibular end organs. 25 VEN also received afferent fibers from neurons in MVN, and this suggested that VEN, MVN, and the vestibular periphery formed a short signal circuit. 31 It has been shown that VEN could mediate vestibular afferent discharge and exert inhibitory/excitatory modulatory control over afferents or type II hair cells.32–35 The wide neural connection and electrophysiological control of VEN may contribute to vestibular compensation. The findings, which demonstrated a collateral projection from bilateral VEN to cerebellar flocculus and ventral paraflocculus, also suggested a potential role of VEN in vestibular plasticity. 36 In addition, a recent study using alpha9 knockout mice showed that loss of the vestibular efferent system markedly diminished vestibulo-ocular reflex (VOR) adaptation, similarly indicating the crucial role of VEN in vestibular adaptation and compensation. 37 Therefore, the results of this study showing acutely decreased and markedly damaged bilateral VEN at day 3 following unilateral intratympanic 60 mg/ml gentamicin suggested the vestibular efferent activity was reduced, and then may lead to the reduction of vestibular compensation.
Clinically, vestibulotoxicity of gentamicin was primarily used to relieve the symptom of vertigo in MD patients. However, some patients complained of disequilibrium either in the short term or long term after ITG. Compensation in central vestibular pathways may play an important role in vestibular functional recovery after an acute damage of vestibular function. The results of this study showed, in the short term, that disequilibrium might be associated with the damaged and decreased neurons in VEN that displayed a reduced ability of modulating vestibular discharge and a reduced vestibular compensation. The morphology of the VEN at day 30 following ITG showed a relatively normal situation, whereas the number of neurons in VEN at day 30 was similar to that at day 3. Therefore, the persistent disequilibrium symptom may be associated with the loss of neurons in VEN. However, the alteration of neuronal function in VEN after ITG remains unclear. Further studies to explore the electrophysiological function of VEN should be also conducted.
Footnotes
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
All authors have contributed to this article as follows: YZ and CD designed the research and revised it critically for important intellectual content; QW performed the acquisition for the work, analysis of the data, and wrote the manuscript; YK and LP carried out some transmission electron microscopy steps; and all authors have read and approved the manuscript as submitted.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (No. 81300826 [YZ] and 81570917 [CD]).