
Abstract
Connective tissue growth factor (CTGF, also named CCN2) plays an important role in the development of tubulointerstitial fibrosis, which most critically determines the progression to end-stage renal failure in autosomal-dominant polycystic kidney disease (ADPKD), the most common genetically caused renal disease. We determined CTGF expression in a well-characterized animal model of human ADPKD, the PKD/Mhm (cy/+) rat. Kidneys of 12 weeks old (cy/+) as well as (+/+) non-affected rats were analyzed for CTGF RNA and protein expression by RT-PCR, Northern and Western blot analyses, in situ hybridization, and IHC. Besides the established expression of CTGF in glomerular cells in kidneys of wild-type (+/+) animals, in (cy/+) rats, CTGF mRNA and protein were robustly expressed in interstitial, stellate-shaped cells, located in a scattered pattern underlying the cystic epithelium and in focal areas of advanced tubulointerstitial remodeling. Renal CTGF mRNA and protein expression levels were significantly higher in (cy/+) rats compared with their (+/+) littermates. Detection of CTGF expression in cells adjacent to cystic epithelium and in areas of marked fibrosis suggests a role in the local response to cyst development and indicates that CTGF may be a relevant factor contributing to tubulointerstitial fibrosis in polycystic kidney disease.
Introduction
Autosomal-dominant polycystic kidney disease (ADPKD) is the most frequent hereditary renal disease (with a prevalence of about 1:800) and is a common cause of end-stage renal failure. ADPKD is characterized by mutations in the PKD1 or the PKD2 gene. 1 With the exception of pharmacological blockade of the vasopressin (V2) receptor, randomized control studies have failed to introduce promising therapeutic strategies for preventing cyst growth and renal failure in man. To date, routine treatment of ADPKD is mainly restricted to controlling hypertension preferably by angiotensin-converting-enzyme (ACE)-inhibitors or angiotensin II receptor blockers and prevention or therapy of infections. 2
During the progression of ADPKD, renal function is maintained for a long time, and its decline and subsequent loss is mainly driven by fibrotic changes, which increase during the advanced stages of the disease. 3 So far, measures to decrease the progression of these early fibrotic changes have not been adequately included in the treatment of ADPKD. These early alterations are not easy to recognize, but begin assumingly years before fibrosis becomes evident by diagnostic imaging techniques.
A well-characterized genetic animal model resembling human ADPKD is the Hannover Sprague Dawley (cy/+) rat 4 of which a colony has been inbred later on to the PKD/Mhm (cy/+) substrain (polycystic kidney disease, Mannheim, inbred strains of rats).5,6 These animals bear a missense mutation in the Anks6 gene, which determines the development of renal cysts.5,7 Although homozygous animals die soon after birth, exhibiting massive renal enlargements, their heterozygous littermates develop renal cysts and renal failure over a period of several months. 4 In this model, renal cysts arise predominantly from proximal tubules. 8 Analogous to the human disease, cyst development and particularly interstitial fibrosis are the determining factors leading to end-stage renal failure. In this ADPKD rat model, various stages of cyst development and fibrosis can be observed and examined simultaneously in the same kidney of an adult animal at an age of 3 months. In some cortical areas, cystic changes and fibrotic scarring are already far advanced, while in other areas, more moderate changes are detectable. 9
Although genes triggering the disease in the human and in animal models have been identified,10,11 the molecular pathways leading to cyst enlargement and especially interstitial fibrosis are still not completely understood. Connective tissue growth factor (CTGF), recently renamed to CCN2, is a 38-kD cysteine-rich peptide that belongs to the CCN family of matricellular proteins.12,13 Increased CTGF mRNA expression has been shown in the human kidney in a wide variety of renal diseases associated with glomerulosclerosis or tubulointerstitial fibrosis, such as diabetic nephropathy, cresentic glomerulonephritis, and IgA nephropathy. 14 CTGF interacts with and binds to diverse groups of multi-ligand receptors and participates in early and late stages of fibrosis.13,15 Most recent data hint for a role of CTGF in fibrosis-associated renal lymphangiogenesis by interaction with vascular endothelial growth factor - C (VEGF-C). 16
In many cases, as in induction of the synthesis of extracellular matrix components, fibroblast proliferation, and wound repair, CTGF acts together with transforming growth factor TGF-β1.17 –19
Although TGFβ1 and CTGF are assumed to be key players in the progression of renal fibrosis, the involvement of CTGF in the course of ADPKD has not been demonstrated. CTGF is induced by angiotensin II (ANG II) in renal fibroblasts. 20 ANG II can stimulate CTGF expression by TGF-β-dependent 21 as well as TGF-β-independent pathways. 22 Because components of the renin-angiotensin-aldosteron system (RAAS) able to produce active ANG II, such as renin and angiotensinogen, are elevated in human ADPKD 23 and in various animal models, one may speculate that in polycystic kidney disease, local production of ANG II may also be involved in the regulation of CTGF expression and subsequent fibrosis.
However, only very limited data about CTGF exist regarding cyst formation and matrix deposition in ADPKD.24 –26 In the Pkd1nl,nl mutant mouse, increased CTGF gene expression can be found, which is probably the result of activated Hippo signaling pathways. 26 In the present study, using non-radioactive in situ hybridization and IHC, we could demonstrate distinct CTGF mRNA and protein expression in areas of interstitial fibrosis and in fibroblast-like cells underlying cystically transformed tubules in the PKD/Mhm (cy/+) rat model resembling human ADPKD.
Materials and Methods
Animals
All procedures performed on animals were carried out in accordance with the German animal welfare act and were approved by the local authorities (ref. number G78/07 and I 15/10). Twelve-week-old male rats of the PKD/Mhm (cy/+) substrain were used for the experiments. This inbred substrain originates from the Han:SPRD (cy/+) rat. 27 Male (cy/+) as well as unaffected (+/+) rats were anaesthetized by ketamin hydrochloride (5.8 mg/100 g) and xylazine hydrochloride (0.39 mg/100 g) and sacrificed by retrograde perfusion through the distal abdominal aorta with 3% paraformaldehyde at a pressure level of 180 mm Hg for 3 min. The kidneys were removed, sliced, and embedded in paraffin. In a parallel set of experiments, the fixation step was omitted and kidneys were removed and processed for quantitative RT-PCR, Northern and Western blotting. The cystic phenotype of each kidney was visually verified by H&E staining. All slides were examined by a Leica DM RB microscope equipped with a Leica DFC 450 camera (Leica; Wetzlar, Germany) for documentation.
Probes
A 303 bp rat CTGF cDNA fragment was amplified by RT-PCR from total rat kidney RNA using internal primers of the published sequence (containing nucleotides 1252–1554 of rat CTGF Accession: NM_022266.2; Primers: FW: ATGGAGACATGGCGTAAAGC, RV: CACTCTTCCAGGAGGCTCAC). This fragment does not share any significant homologies to other transcripts in the rat including the other members of the CCN family such as CCN3. The PCR fragment was cloned into a pCRII-TOPO vector (Invitrogen; Carlsbad, California). Sense and antisense CTGF cRNA probes were generated using an RNA digoxigenin labeling kit (GE-Healthcare; Mannheim, Germany). P32-labeled rat specific CTGF and 18S cDNA probes were generated using a random hexamer primer and Klenow polymerase. 28
Northern Blot Analysis
Total RNA was isolated by homogenizing frozen kidneys in cold guanidine isothiocyanate buffer and a subsequent extraction with phenol chloroform. RNA samples were run on agarose gels and blotted onto nylon membranes according to standard protocols. After transfer to a Biodyne A nylon membrane (Pall Gelman Laboratory; Ann Arbor, MI), the filters were probed with a P32-labeled rat specific CTGF or 18S cDNA probes. CTGF values were normalized to the 18S RNA signal. Quantification was performed using ImageJ 1.46 (downloaded from http://imagej.nih.gov/ij/download.html).
Quantitative RT-PCR
RNA was reversely transcribed using ReverseAid reverse transcriptase (New England Biolabs; Frankfurt am Main, Germany) and subjected to quantitative PCR (qRT-PCR) using CTGF primers (FW: CTGACCTAGAGGAAAACATT, RV: AGAAAGCTCAAACTTGACAG) and Cybergreen mastermix (GE-Healthcare). Expression was calculated by the ΔΔCT method 29 using 18S RNA (FW-primer: CCTATCAACTTTCGATGGTA, RV-primer: TCCTGTATTGTTATTTTTCG) as reference.
Western Blot Analysis
The kidneys were crushed in liquid nitrogen and subsequently homogenized in ice-cold RIPA buffer containing 10 mmol/L EDTA, 5 mmol/L benzamidine, 10 mmol/L N-ethyl-maleimide, 1 mmol/L phenylmethyl sulfonyl fluoride. Forty µg of protein was loaded on a 12.5% SDS-PAGE gel and subsequently transferred onto a PVDF membrane (Merck-Millipore; Darmstadt, Germany). Nonspecific binding sites were blocked with 5% horse serum in TBS (pH 7.4) and incubated with a goat anti-human CTGF antibody, cross-reacting with rat and mouse, diluted 0.2 µg/ml (sc-14939; Santa Cruz Biotechnology Inc., CA) overnight at 4C. After washing, the membrane was incubated for 1 h with horseradish peroxidase-labeled donkey-anti-goat IgG, diluted 1:50,000 (Santa Cruz Biotechnology Inc.). The antibody was detected by the enhanced chemiluminescence detection kit (ECL, GE-Healthcare). Densitometric analysis was performed using ImageJ 1.46 (downloaded from: https://imagej.nih.gov/ij/download.html). A rat CTGF specific band was detected between 35 and 40 kDa.
In Situ Hybridization
Five µm sections of paraformaldehyde-fixed paraffin-embedded rat kidney sections were dewaxed, rehydrated, proteinase K-treated, dehydrated, and hybridized overnight at 42C using a hybridization mix containing digoxigenin-labeled riboprobe (500 ng/ml), 50% deionized formamide, 4× SSC, 10 µg/ml heparin, 25 µg/ml yeast t-RNA, and 0.2% blocking reagent (GE-Healthcare). Slides probed with antisense or sense rat digoxigenin-labeled CTGF cRNAs were then subjected to RNase A treatment (10 µg/ml, 30 min) and washing with 0.1 × SSC in 50% formamide at 45C for 30 min. Signal detection was carried out using an alkaline phosphatase-coupled anti-digoxigenin antibody (Dako GmbH; Hamburg, Germany) and NBT/BCIP (Sigma-Aldrich; Munich, Germany) as substrate for color development. Control hybridizations using sense cRNA were free of any specific staining.
IHC
After deparaffination and antigen retrieval by microwave treatment (10 min in 0.01 M citrate buffer pH 6), slides were blocked for 30 min with 20% fetal bovine serum diluted in PBS. The sections were then incubated with one or two primary antibodies (in double labeling experiments). The following antibodies were used: goat anti-human CTGF (final concentration: 10 µg/ml in PBS containing 5% BSA, sc-14939; Santa Cruz Biotechnology Inc.) recognizing human, rat, and mouse CTGF; mouse monoclonal anti-porcine vimentin (V9; final concentration 0.5 µg/ml, sc-6260; Santa Cruz Biotechnology Inc.) recognizing rat vimentin. For staining of podocytes, double IHC experiments were performed with anti-CTGF antibody and a mouse monoclonal antibody to synaptopodin, 30 recognizing the rat protein, supplied as hybridoma supernatant, clone G1D4 (cat. no. 65194; Progen Biotechnik GmbH, Heidelberg, Germany). This was applied as ready-to-use solution (IgG1 content approximately 2–5 µg/ml).
After incubation of the sections with the primary antibodies for 1 h at room temperature and then overnight at 4C in a moist chamber, sections were washed three times with PBS and incubated with a Cy3-labeled donkey anti-goat antibody, diluted 1:200 (Jackson ImmunoResearch; Newmarket, Suffolk, UK) alone, or in combination with a Cy2-labeled rabbit anti-mouse antibody, diluted 1:100 (Jackson ImmunoResearch) for 1 hr in the dark, finally washed and mounted in Moviol. In all slides, nuclei were stained with 4′,6 diamidino-2-phenylindole dihydrochloride (DAPI, 1 µg/ml).
Statistical Analysis
The differences between the two experimental groups were determined by a Mann Whitney U test: p Values of less than 0.05 were considered to be statistically significant.
Results
To compare the expression of CTGF mRNA in the kidneys of 12-week-old affected heterozygous (cy/+) rats and their unaffected (+/+) littermates (five of each group), we isolated RNA from whole tissue homogenates and performed quantitative RT-PCR with rat CTGF specific primers. Analysis showed that (cy/+) animals exhibited a 2.2 ± 0.9-fold higher renal CTGF mRNA expression than their unaffected littermates (Fig. 1A). PCR data were verified and validated by performing Northern blot analysis of tissue isolates from different animals of each genotype. With this method, a 3.0 ± 0.76-fold higher renal CTGF mRNA expression was apparent in kidneys of cystic animals (Figs. 1B and C) when compared with wild-type animals. Accordingly, CTGF protein was also elevated in affected kidneys of PKD/Mhm (cy/+) rats. Western blots performed with protein extracts from whole kidney homogenates revealed a band between 35 and 40 kDa (Fig. 2B). Densitometric analysis displayed a 2.68 ± 1.2-fold increase in the renal CTGF protein expression adjusted to actin, in cystic kidneys versus non-cystic littermates (Fig. 2A). Western blot experiments using two other CTGF antibodies that were specific for rat and human CTGF (sc-25440; a rabbit anti-human CTGF and sc-101586; a mouse monoclonal anti-human CTGF, Santa Cruz Biotechnology Inc.) yielded comparable results (data not shown).
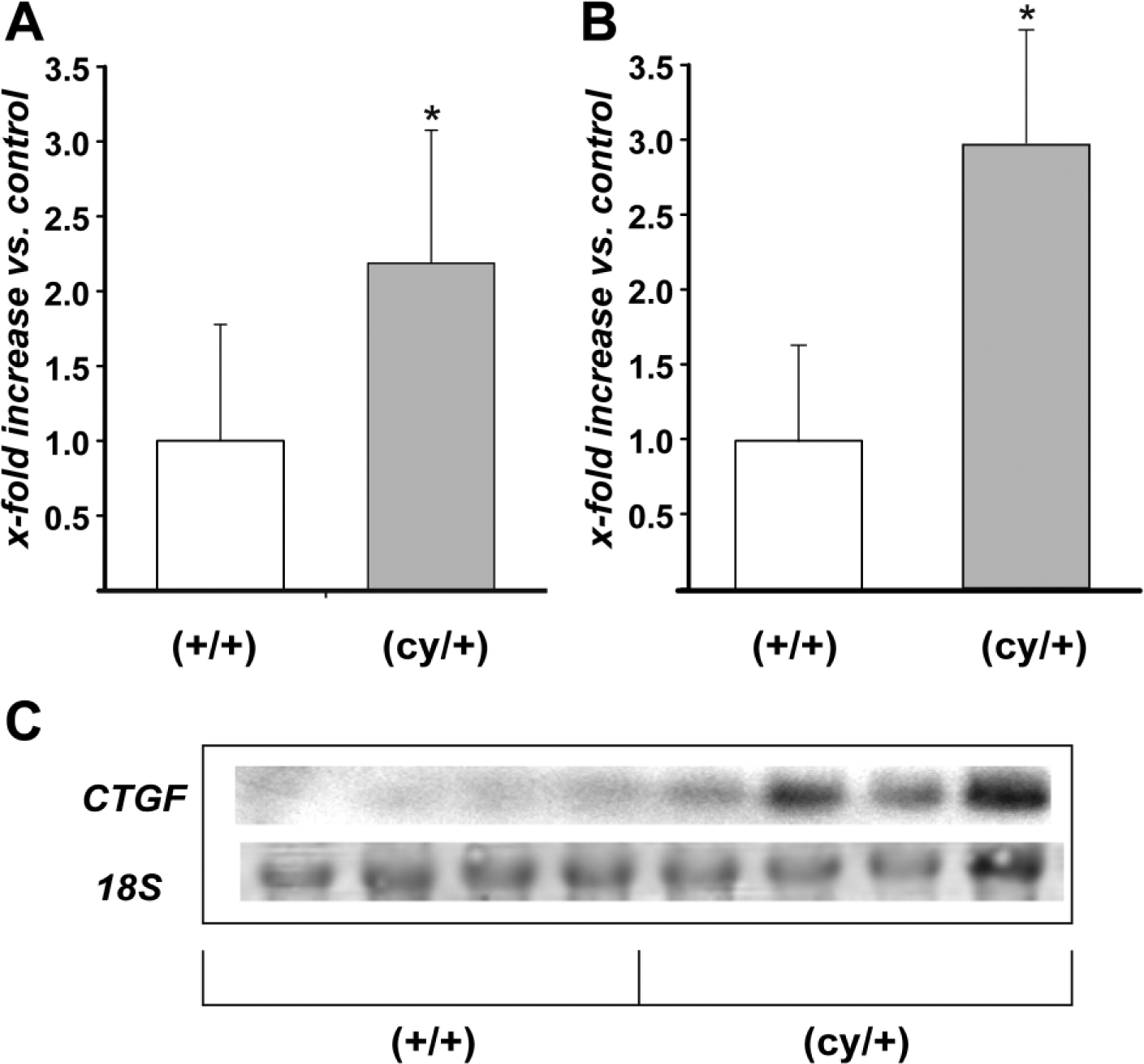
(A) Quantitative RT-PCR of CTGF in the kidneys of (cy/+) and control (+/+) rats, 5 animals per group. (B, C) Northern blot analysis of CTGF mRNA expression in (cy/+) and (+/+) kidneys, 4 animals per group (C). CTGF signal was normalized for the corresponding 18S RNA. (A, B) represents densitometric analysis. Data expressed as mean ± SD; *p<0.05. Abbreviation: CTGF, connective tissue growth factor.
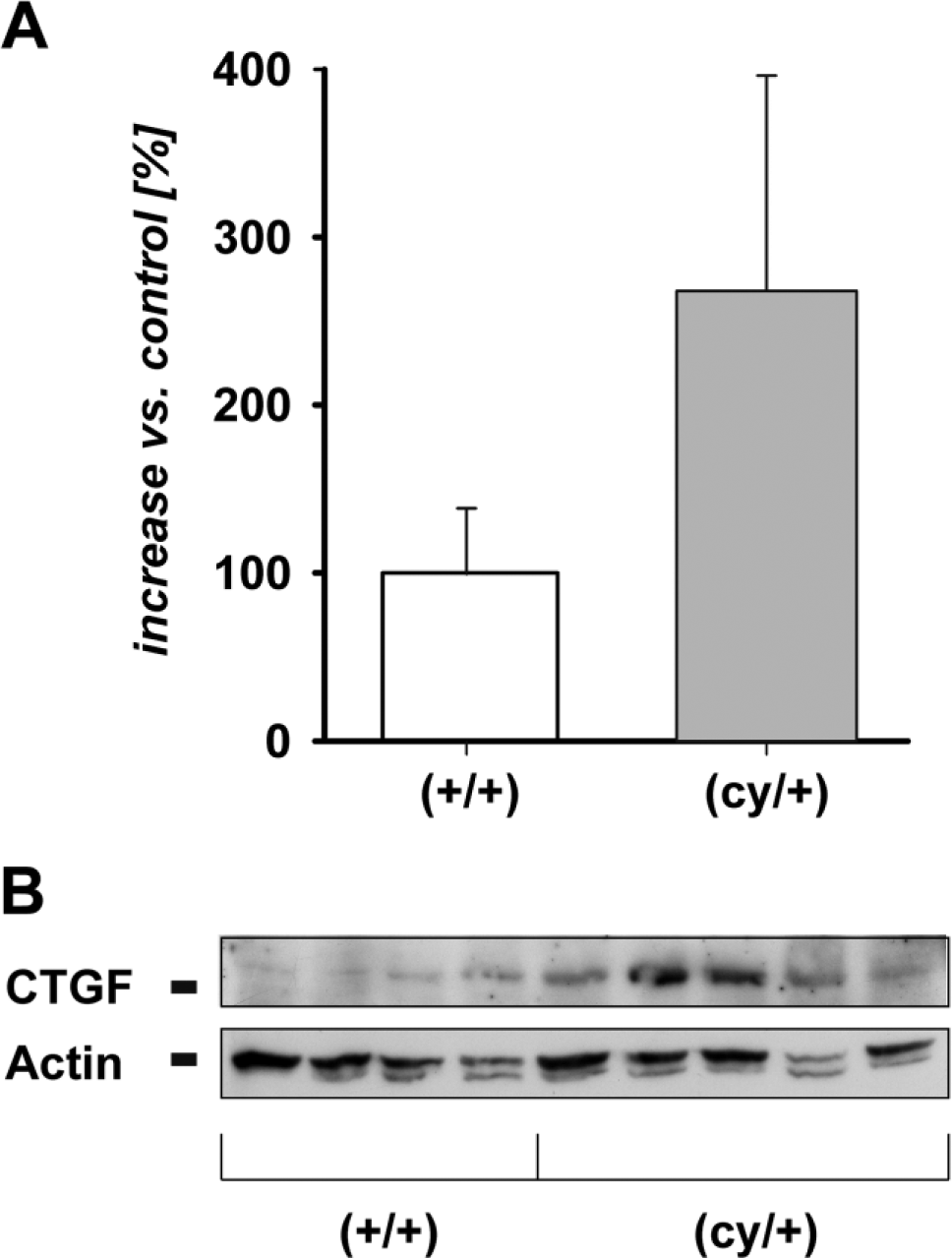
(A) Densitometric comparison represents mean ± SD from 5 (cy/+) and 4 (+/+) animals. (B) Western blot analysis of CTGF of 5 (cy/+) and 4 control (+/+) kidneys. 40 μg of total protein from whole kidney lysates was applied to each lane. β-Actin is used as an internal control; p<0.05. Abbreviation: CTGF, connective tissue growth factor.
To define the renal cells responsible for the observed increase in CTGF mRNA production, we performed in situ hybridization with tissue sections of 12-week-old (cy/+) and (+/+) PKD/Mhm rats (Fig. 3). Specificity of signals was verified by control hybridizations with DIG-labeled sense CTGF cRNA probes, which did not produce any signals (Fig. 3A). In unaffected (+/+) rat kidneys, the distribution of CTGF mRNA was, as expected, limited to podocytes of glomeruli, while almost no other cells in the renal cortex expressed CTGF. A different expression pattern of CTGF was observed in kidneys of (cy/+) rats, undergoing cystic transformation of the proximal tubules. In kidney cortex, robust CTGF mRNA expression was present in individual cells underlying the epithelium of many cystically transformed proximal tubules and in regions with fibrosis (Fig. 3C, overview). CTGF mRNA containing cells were found to be located isolated or in clusters encompassing cystic profiles, often in proximity to the epithelium of the cyst walls (Figs. 3D and E). Moreover, CTGF expression was detected in numerous cells located in areas of advanced interstitial fibrosis (Fig. 3F). Distinct interstitial localization of these CTGF-expressing cells indicate that they presumably represent fibroblast-like cells. In addition, CTGF transcripts were detected in flattened epithelia of a few dilated tubular profiles, which exhibited an atrophic appearance (Fig. 3G). The robust and exclusive cytoplasmic CTGF mRNA localization in podocytes and interstitial cells is highlighted in Figs. 3H and I.
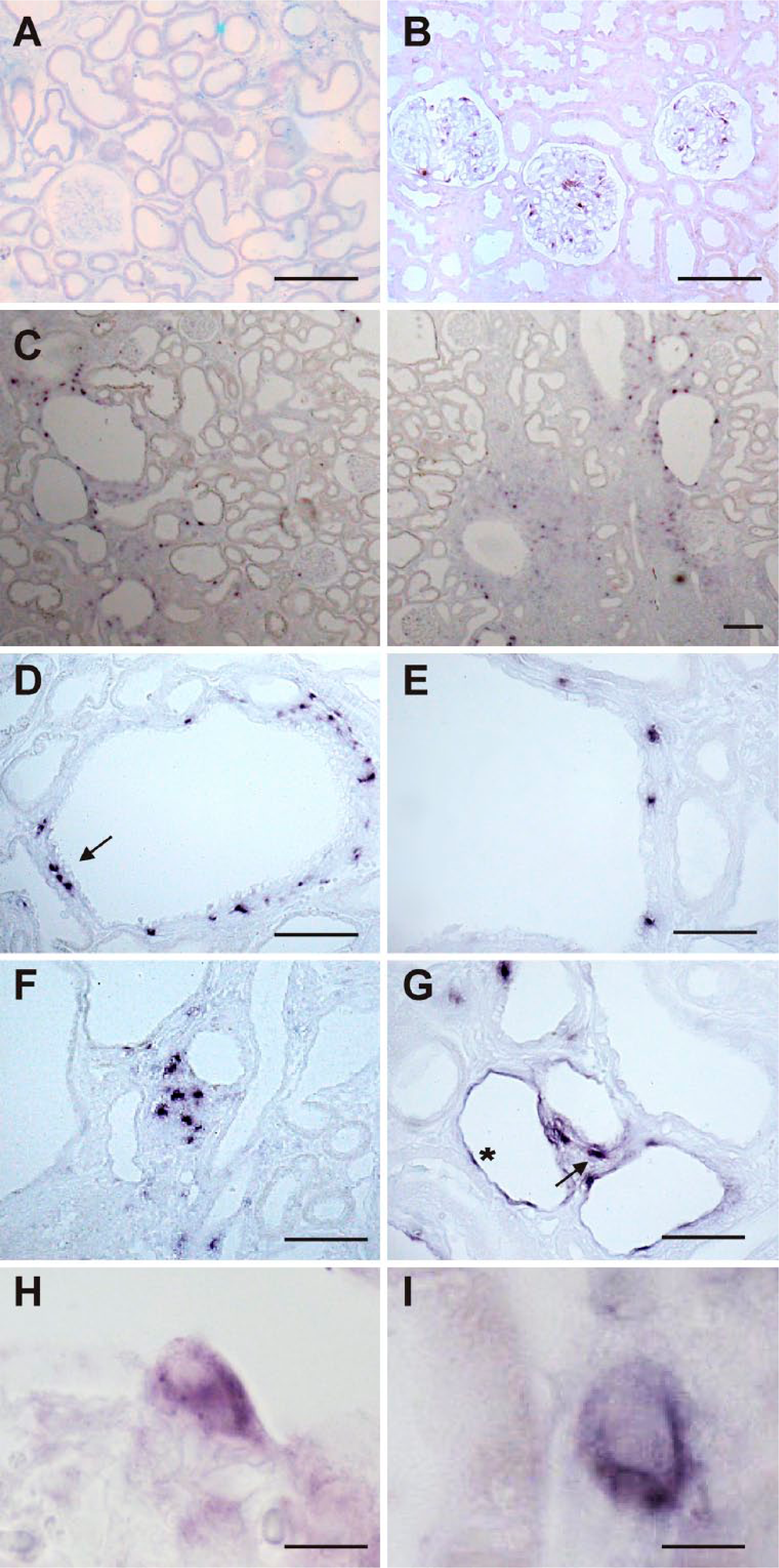
ISH for CTGF mRNA on adult PKD/Mhm rat kidneys cortex. (A, B) represent (+/+), (C–I) represent (cy/+) animals. (A) Hybridization with sense control probe did not show any signal in rat kidney cortex. (B) ISH with specific antisense probe shows labeling (dark blue) restricted to glomerular podocytes where CTGF is known to be expressed. (C) Overview of the inner cortex: In cystically transformed kidneys, multiple but distinct sites of CTGF mRNA expression can be detected (dark blue), preferentially in the vicinity of proximal cysts, but also in areas with fibrosis, seen in particular on the right. (D) Detailed view of a cyst from a (cy/+) kidney with multiple CTGF-expressing cells in the interstitial compartment surrounding the cyst, but not in the respective epithelia; a cluster of positive cells is marked with an arrow. The pericystic expression pattern is again highlighted in (E). (F) shows particular and focal interstitial expression of CTGF mRNA in an area of extensive matrix remodeling and cystic transformation. In (G), sporadic mRNA expression in epithelia of flattened dilated distal tubules (example is marked with an asterisk) can be identified, besides interstitial expression (example is marked with a small arrow). (H) High power magnification of a glomerulus demonstrating strong cytoplasmic CTGF mRNA expression in a podocyte protruding into the urinary space, the nucleus is devoid of a signal. (I) shows detailed view of an interstitial cell expressing CTGF mRNA, located adjacent to the tubular epithelium, which is shown on the left. Scale bars: (A, B, F, G) = 100 µm, (C, D, E) = 90 µm, (H) = 24 µm, (I) = 12 µm. Abbreviations: ISH, in situ hybridization; CTGF, connective tissue growth factor; PKD, polycystic kidney disease.
Kidney sections of (+/+) and (cy/+) rats immunohistochemically investigated with a CTGF antibody revealed a protein expression pattern consistent with that of the corresponding mRNA: As shown in Figs. 4A and B, renal CTGF protein expression in kidneys of unaffected rats was exclusively confined to glomerular podocytes, while in (cy/+) rats, CTGF was also strongly expressed in fibroblast-like cells underlying the epithelium of transformed proximal tubules. Areas around these tubules differ in the extent of extracellular matrix remodeling (Figs. 4C–E). As for the corresponding mRNA, CTGF protein was also detected in some epithelial cells within altered and flattened tubules, most likely of distal origin (Fig. 4F). High resolution magnification indicates an exclusive expression of CTGF in the cell body of podocytes, which can be discerned from the foot processes by double IHC with synaptopodin (Figs. 5A–C). High-power view indicates the stellate-shaped character of the CTGF-expressing cells (Fig. 6A). Moreover, the assumption that CTGF is synthesized by fibroblast-like cells in the vicinity of cysts was demonstrated by double-labeling experiments showing positive co-staining for vimentin, a non-exclusive widely used marker for fibroblasts (Figs. 6B–D). In this model, vimentin is, in addition, expressed in cells of dedifferentiating epithelia. 9
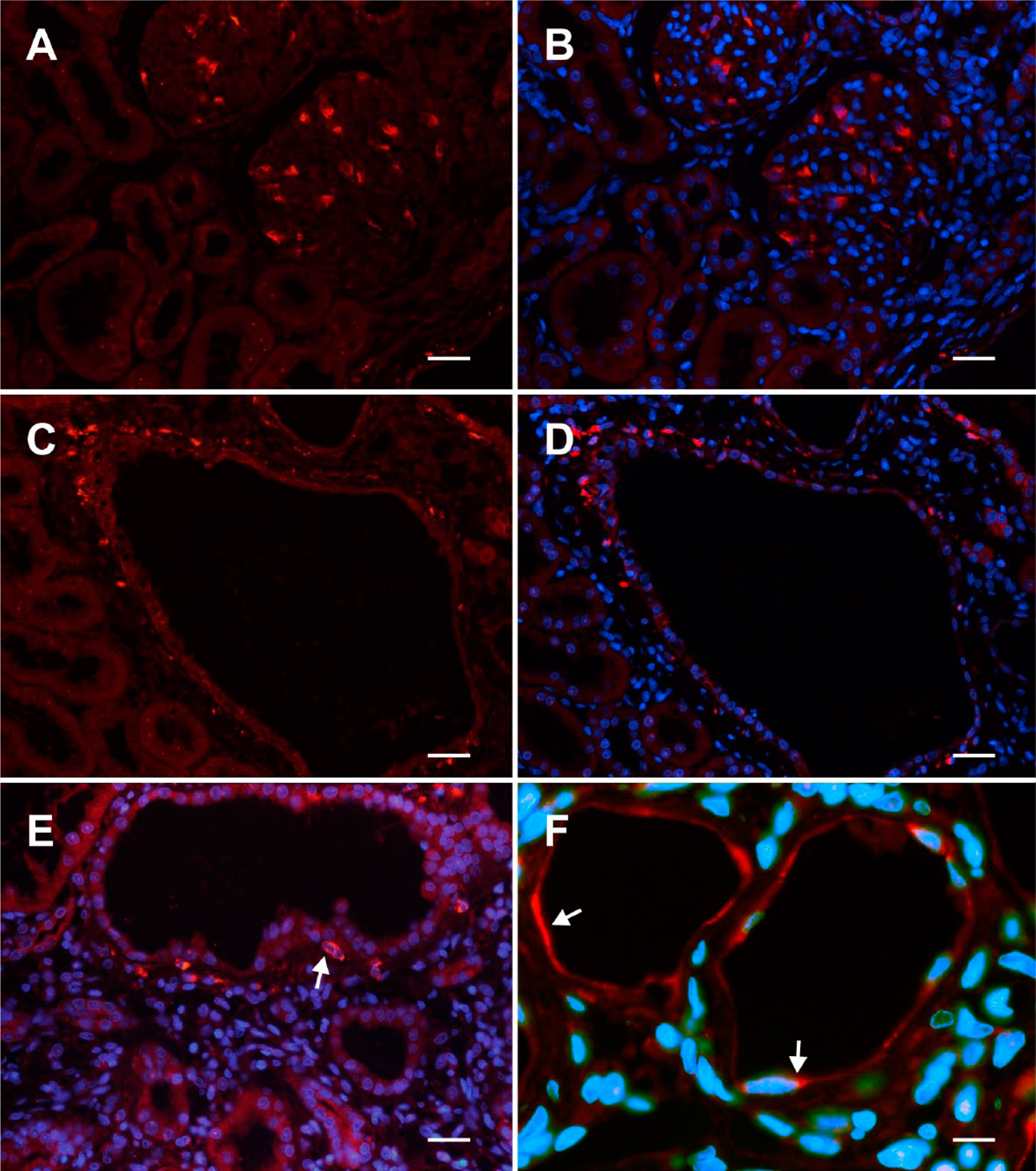
Immunofluorescent labeling (Cy3, red) of CTGF on adult PKD/Mhm rat kidney section from kidney cortex. (A, B) represent (+/+), (C–F) represent (cy/+) animals. In sections (B, D, E, F), nuclei are stained with DAPI fluorescence (in blue). (A, B) Two adjacent glomeruli exhibited a strong cytoplasmic staining of CTGF in multiple podocytes. Panel (C) represents an overview, where a large cyst is surrounded by CTGF-positive cells, particularly on the upper left. (E) Heavy immunofluorescent cytoplasmic CTGF staining in the vicinity of a cyst (example is marked with an arrow) and illustrated by nuclear counterstaining. (F) Two slightly dilated tubules with thinned epithelia show delicate CTGF immunoreactivity (examples are marked with arrows). Scale bars: (A, B) = 35 µm, (C, D) = 45 µm, (E) = 30 µm, (F) = 15 µm. Abbreviations: CTGF, connective tissue growth factor; PKD, polycystic kidney disease; DAPI, 4′,6 diamidino-2-phenylindole dihydrochloride.
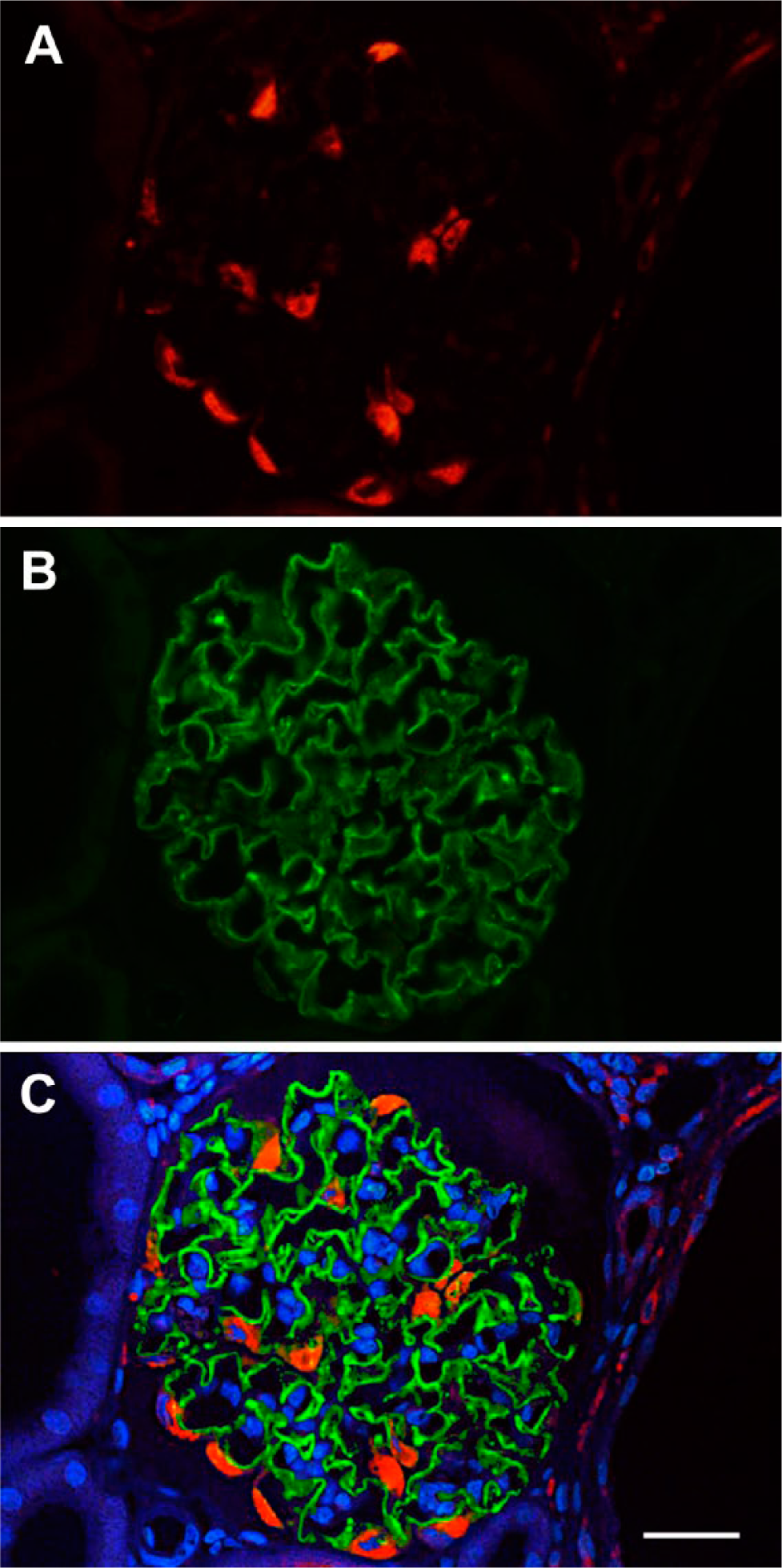
Double IHC with anti-CTGF (A, Cy3 red) and anti-synaptopodin (B, Cy2, green) of a glomerulus of a cy/+ rat in the vicinity of a greater cyst (on the right). Merged view and nuclear staining (DAPI, blue) is provided in (C). CTGF protein is exclusively expressed in the cell bodies of podocytes (A), which are clearly identifiable at the border to the urinary space. There is no overlap with synaptopodin-positive foot processes, which are directly connected. Scale bar = 37 µm. Abbreviations: CTGF, connective tissue growth factor; DAPI, 4′,6 diamidino-2-phenylindole dihydrochloride.
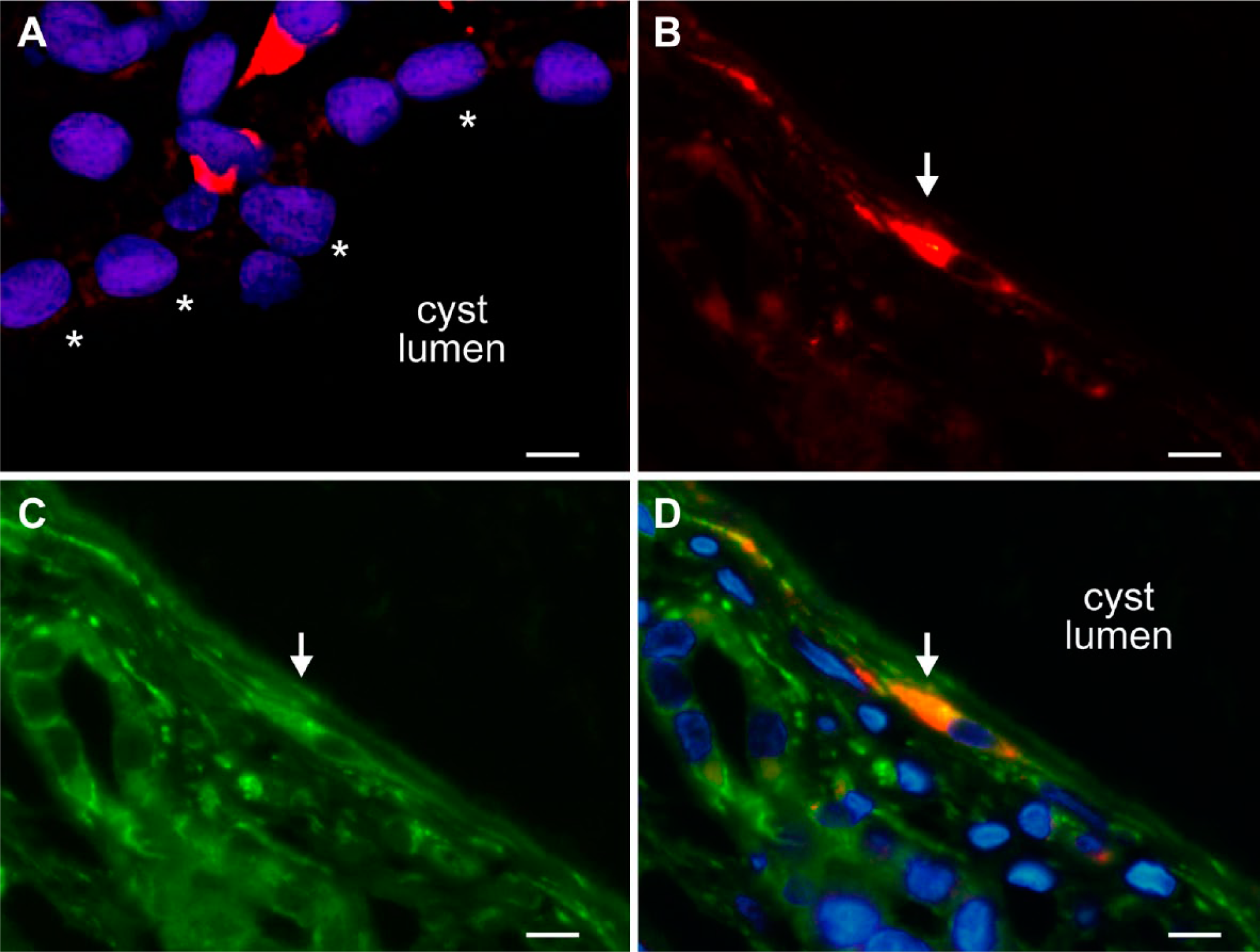
(A-D): Immunofluorescent labeling (Cy3, red) with anti-CTGF antibody and anti-vimentin antibody (Cy2, green) or nuclear staining (DAPI, blue) on (cy/+) kidneys. (A) A high-power magnification showing IHC with anti-CTGF antibody (Cy3, red) and nuclear staining with DAPI fluorescence (blue): Two interstitial cells express CTGF in the cytoplasm, exhibiting a stellate morphology (top of the figure). The cyst epithelial wall (showing normal-sized nuclei) is marked by asterisks. (B–D) displays a double labeling for CTGF and V9. An interstitial stellate-shaped cell (marked with arrow) expresses CTGF (B, Cy3, red), and vimentin (C, Cy2, green), both signals clearly overlap in the cytoplasm in the merged view in (D), which contains nuclear staining (blue). The cyst here is lined by an epithelium of extremely flattened morphology. Scale bars: (A) = 8 µm, (B–D) =10 µm. Abbreviations: CTGF, connective tissue growth factor; DAPI, 4′,6 diamidino-2-phenylindole dihydrochloride.
Discussion
CTGF is a pleiotrophic factor that exerts vital roles, for example, in wound healing and organ development, 31 but in renal pathophysiology, this molecule has attracted attention by its potential role in the contribution to renal fibrosis. 19 A recent study using an affymetrix expression chip revealed that CTGF was among the genes whose expression appeared to be increased in kidneys of PKD/Mhm (cy/+) rats when compared with their unaffected littermates. 24 Interestingly, another member of the CCN family, closely related to CTGF, CCN3 also called NOV was shown to be downregulated in that study. 24 Cystic kidney disease is not only characterized by proliferation and dedifferentiation processes within the tubular epithelium layer but also by an increasing remodeling of the inter-tubular extracellular matrix going along with an expanding tubulointerstitial deposition of extracellular matrix components. The extent of the resulting interstitial fibrosis is one of the key elements determining the loss of renal function in cystic kidney disease over time. During interstitial fibrosis, CTGF is synthesized by activated myofibroblasts in a possibly autocrine fashion as shown in the heart. 32 Presumably synthesized initially to facilitate tissue repair processes, an overshooting production of CTGF by these cells may contribute to the onset and the progression of renal fibrosis. In the PKD/Mhm (cy/+) rat model of human ADPKD, we observed fibroblast-like cells to be the main source of CTGF within the kidney. This is in accordance with previous analysis that demonstrated that CTGF is only expressed by resident cells in the diseased kidney. 33 In affected (cy/+) kidneys, these fibroblasts, mainly underlying tubular epithelium of various states of dedifferentiation, produce significant amounts of CTGF mRNA and protein. Based on the immunohistochemical and in situ hybridization data, it must be assumed that these cells are responsible for most of the overall increase in renal CTGF mRNA and protein, detected by quantitative RT-PCR, Northern and Western blotting. This excessive CTGF synthesis may enforce the changes in the extracellular matrix deposition, which can be observed during the progression of the renal disease in ADPKD, finally leading to a loss of renal function. 34 Our previous studies in this rat model have shown that specific matrix metalloproteinases (MMP) and their tissue inhibitors (TIMP) are expressed in the tubulointerstitial compartment in areas where tubular epithelium has undergone cystic transformation. 35 In these areas, we observed a pronounced CTGF expression. CTGF may also induce other factors involved in fibrotic processes, such as TIMPs, that inhibit matrix degrading enzymes and, thus, augment the expansion of the extracellular matrix as seen in a wound healing scenario. 36 Treatment with an mTOR inhibitor attenuated MMP expression and reduced extracellular matrix alterations. 37 The hypothesis of a close connection between increased CTGF expression, fibrosis (and conceivably even cyst formation) in our ADPKD model is supported by the observation that CTGF is only barely expressed in areas exhibiting no or only slight proximal tubular dedifferentiation and cyst formation.
On one hand, ADPKD is known to be associated with a pathologically activated RAAS. 23 The activity of the RAAS is one factor that can determine synthesis and release of CTGF. 21 Ang II is a key component of the RAAS and has been demonstrated to regulate CTGF expression via a TGF-β-dependent and TGF-β-independent pathways.38 –40 Although the interaction between CTGF and TGF-β is important, the role of TGF-β is, on the other hand, still uncertain in ADPKD because, for example, one study shows that TGF-β inhibits cystogenesis in human autosomal dominant polycystic kidney epithelial cells. 41 In contrast, rosiglitazone, an oral antidiabetic agent, inhibits TGF-β1 mediated fibrogenesis in ADPKD cyst-lining epithelial cells. 42 TGF-β isoforms may have different roles in promoting or diminishing fibrosis in the context of ADPKD, perhaps depending on their expression levels and site of origin. Our data demonstrate that only a small number of cells produce CTGF, but CTGF might also be secreted into the extracellular compartment, thereby reaching different structures within the tubulointerstitial area.
Another issue relates to the role of the CTGF transcript that has an exceptionally long 3′UTR, containing multiple interaction sites for known microRNAs as well as RNA stabilizing proteins. Because Ang II is known to mediate some of its effects on RNA expression via interaction with these factors, 43 at least a part of the observed increase in CTGF RNA may be mediated by RNA stabilization via the CTGF 3′UTR. Recently, Gao et al. describe miRNA-19b to enhance CTGF expression in cardiomyocytes. 44 A decline in renal CTGF production may account at least in part for supposed beneficial long-term effects of a RAAS blockade observed in human ADKPD patients,45,46 which could be due to antifibrotic actions. In this regard, recent data from murine transgenic cardiomyocytes and cardiac fibroblasts point to the role of aldosterone-induced pro-fibrotic signaling involving CTGF. 47 A reduced renal expression of CTGF has been proposed to be a key effect in a clinical study relating to the mechanisms of a RAAS blockade in diabetic nephropathy. 48
Interestingly, diabetic nephropathy is also characterized by severe alterations in the tubulointerstitial area. However, these changes are seen in the context of severe glomerular damage and concomitant proteinuria, whereas ADPKD exhibits classical characteristics of a pure chronic tubulointerstitial disease without prominent glomerular lesions. Contrasting with our findings in rat models for diabetic nephropathy, CTGF protein was detected de novo in proximal tubular cells, but not in the interstitium.49,50
Beside activation of the RAAS, hypoxic effects, caused by pressure of fluid filled cysts on the surrounding tissue and also on the capillary network may account for the observed increase in local CTGF. It has been demonstrated that hypoxia-inducible factor-1 (HIF-1) can induce CTGF transcription in fibroblasts. 51 Therefore, the observed expression CTGF in affected kidneys of PKD/Mhm rats may be the result of multiple upstream effectors.
There is almost no data supporting that CTGF may also participate in early processes of cyst formation. Different members of the CCN family display functional interaction with Wnt signaling pathways. CTGF can inhibit Wnt signaling, however, Wnt3A/β-catenin induces CTGF on a transcriptional level. 52 As a role of disturbed Wnt signaling in cyst development has been described whereby decreased Wnt/β-catenin activity has been found in some forms of cystic disease, an involvement of CTGF appears to be also reasonable in early cystogenesis. The exact effect of CTGF on the Wnt signaling pathway might depend on the cell types and microenvironment. CTGF binds to the Wnt co-receptor LDL receptor-related protein 6 (LRP6) in a distinct manner than it interacts with bone morphogenic protein (BMP) or TGF-β. 53 Interestingly, new data obtained from pericytes show that differently cleaved domains of CTGF along with the Wnt signaling may play substantial roles in fibrosis. 54 Primarily, interventions in the CTGF pathway may have the potential to reduce excessive fibrosis in ADPKD patients and, thus, retard the loss of renal function.
The present data, corroborated by different biochemical and histochemical techniques, strongly demonstrate an association between upregulated expression of CTGF in polycystic kidney disease and tubulointerstitial changes during cyst formation. Inhibition of CTGF expression or signaling should be further on evaluated as a potential target for an antifibrotic option in this particular disease.
Footnotes
Acknowledgements
The excellent technical support of Sandra Fay is greatly acknowledged.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
SG and NO conceived the study, analyzed the data, and wrote the manuscript. YH, BK, and SCH performed animal and experimental work. NG, IAH, MG-S, and HG participated in data reviewing and results interpreting. All authors have read and approved the final manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported (only material costs for experiments) by the Ellinore and Herbert Vogel-Foundation, Germany (N.O., S.G.), and was supported by a grant from the National Institute of Health (NIH), Bethesda, USA (5RO1DK100482) (SCH).