
Abstract
The probiotic mixture VSL#3 attenuates colitis in patients with Inflammatory Bowel Disease (IBD) and in animal models of this condition, but the mechanisms involved are incompletely understood. VSL#3 alters macrophage morphology and secretory profile in vitro in a polarization-dependent manner. We examined the effect of VSL#3 on macrophages in acute trinitrobenzene sulfonic acid-induced colitis. Rats were randomized to normal, colitis, or colitis+VSL#3 groups. After sacrifice, the colons were evaluated for macroscopic and microscopic damage. Serum cytokine levels were measured, and microbiome analysis undertaken. Total and M1 colonic macrophages, and total and proliferating hepatic macrophages were assessed by double immunofluorescence staining. Colitis+VSL#3 rats had lower macroscopic damage, with less microscopic damage in the proximal colon, compared with colitis alone. Colitis significantly increased colonic macrophage infiltration, which was significantly reduced by VSL#3 treatment. VSL#3 did not decrease the colitis-induced surge of colonic M1 macrophages or hepatic macrophages. VSL#3 reduced colitis-induced serum cytokine levels, and induced restoration of colonic transcript levels for pro-inflammatory, anti-inflammatory, and barrier proteins to, or past, normal levels. Fecal bacteria distribution changed between groups. In summary, the probiotic VSL#3 reduces colitis severity, colonic macrophage infiltration, and serum cytokine levels, but does not dampen the pro-inflammatory phenotype of M1 macrophages.
Introduction
Inflammatory bowel disease (IBD) is a chronic and debilitating condition of the intestines that is becoming increasingly common in the United States and around the world. 1 Although the exact cause remains elusive, microbial and immune interactions are involved in the pathogenesis of IBD. 2 Crohn’s disease and ulcerative colitis, the two main forms of IBD, frequently manifest in the colon, the site in the body with the highest microflora concentration. 3 Dysbiosis, or altered gut microflora, is common among IBD patients4,5 and can cause colitis development in rodents. 6 Targeting this dysbiosis with probiotics, living microbes with beneficial qualities, has proven to be effective both in animal models of IBD and in clinical trials with IBD patients and is actively being investigated as a therapeutic strategy for patients with IBD.
Uncovering the mechanism of action for the different probiotics that have shown promise for treating IBD is of great interest. Hayashi and colleagues have recently reported that the probiotic Clostridium butyricum reduces the intensity of colitis in mice treated with dextran sodium sulfate (DSS) by triggering the secretion of IL-10 in colonic macrophages. 7 The macrophage is one of the most numerous immune cells in the colon,8,9 which, in turn, is one of the most macrophage-rich organs. 10 Colonic macrophages are remarkably phagocytic, and they routinely phagocytose bacteria that translocate across the epithelial barrier.11-15 However, they do not mount an inflammatory response against these microbes16,17 and, instead, respond with an anti-inflammatory profile.11,12,14,18 Furthermore, macrophages in the colon promote regulatory T-cell expansion.19,20 Macrophages can be classified as pro-inflammatory M1 or anti-inflammatory M2 macrophages. 21 Therefore, under homeostatic conditions, colonic macrophages can be regarded as M2 macrophages. 22 Nevertheless, M1 macrophages accumulate in the colon of animals undergoing experimental colitis and of patients with IBD.12,15,22-26 This dual role of macrophages in the colon makes them interesting therapeutic targets. 22
The effect that the probiotic mixture VSL#3, which has shown promise in treating patients with IBD,27,28 has on colonic macrophages is incompletely understood. It has been suggested that the probiotic mixture VSL#3 exerts its anticolitic effects by reducing M1 macrophages in the inflamed colon. 29 We have previously demonstrated in an in vitro model that VSL#3 not only alters the morphology of polarized (M1/M2) and unpolarized macrophages but also modifies the secretion of cytokines and chemokines in these cells in a polarization-dependent manner. 30 In general, the pro-inflammatory phenotype of M1 macrophages was relatively unaffected, whereas M2 and unpolarized macrophages were shifted toward a more balanced phenotype, in which an augmentation of their anti-inflammatory/pro-healing phenotype was accompanied by an increase in their pro-inflammatory potential. To better understand how colonic macrophages respond to this probiotic, the aim of the present study was to examine the effect that treatment with VSL#3 has on macrophages in acute trinitrobenzene sulfonic acid (TNBS)-induced colitis in rats. Consistent with our in vitro results, we found that VSL#3 treatment does not reduce colonic M1 macrophages in this model of colitis, but, instead, decreases macrophage infiltration into the colon. In addition to our studies with the acute colitis model, we also examined macrophage numbers in the colon of rats undergoing a chronic TNBS-colitis. 31 In contrast to our findings with acute colitis, neither chronic colitis alone nor chronic colitis with VSL#3 treatment affected colonic macrophage numbers when compared with controls in the chronic colitis model.
Materials and Methods
Animals
All animal studies were approved by the Institutional Animal Care and Use Committee (IACUC) at Ponce Health Sciences University. Male Sprague-Dawley rats of approximately 7 weeks of age and weighing approximately 200g were randomized to a normal group (n=5), colitis group (n=13), or colitis with probiotic treatment (colitis+VSL#3) group (n=13). Animals were housed individually in a room maintained at 23C with the light/dark cycle alternating every 12 hr. Bedding was autoclaved prior to use, and rats were fed with standard laboratory chow ad libitum. Weight was monitored daily.
Treatment With VSL#3 and Colitis Induction
The probiotic formulation VSL#3 was generously provided by Sigma Tau Pharmaceuticals (Gaithersburg, MD). Based on previous studies, 31 rats in the colitis+VSL#3 group received 10 billion colony forming units (CFU) of the probiotic, dissolved in 150 mL of drinking water and prepared daily, starting 1 week before colitis induction and continuing until the time of sacrifice (Fig. 1). Rats in the normal and colitis groups received regular drinking water during this period. Fluid consumption was monitored daily. Colitis was induced by intracolonic administration of the haptenating agent TNBS, as has been described previously.32-34 Briefly, rats were lightly anesthetized with ether, and a plastic catheter was inserted 8 cm proximal to the anus, reaching the transition between the proximal and middle colon. Through the catheter, 0.5 mL of 60 mg/mL TNBS (Fluka) in 50% ethanol were instilled into the colon of each rat, and the rats were held in an inverted position to minimize expulsion of the TNBS. Animals were sacrificed 72 hr later by overdose with pentobarbital. Normal rats were left untreated. Ethanol was not administered to the normal rats because of two reasons: first, the ethanol serves as a barrier disruptor and can, therefore, cause inflammation by itself, as has been demonstrated by several groups when this model was originally developed and also in follow-up studies; and second, the goal was to compare the effects of the probiotic on the colitis and to either induce colitis (colitis and colitis+VSL#3 groups) or not induce colitis (normal group), rather than to control for the barrier disruptor that enables the TNBS to penetrate the epithelial barrier and exert its role as a haptenating agent. Two rats in the colitis group and one rat in the colitis+VSL#3 group died before completion of the 72-hr period. Two rats from the colitis group leaked a substantial amount of TNBS and were excluded from further studies on the basis of insufficient weight loss and macroscopic damage. Methodology for rats undergoing chronic colitis has been described previously. 31 Briefly, colitis was induced in the same manner as employed for the acute colitis protocol described herein. Afterward, rats were allowed to heal for a period of 6 weeks, subjected to colitis reactivation with daily intravenous administration of TNBS for 3 days, and further treated with 2 weekly intravenous administrations of TNBS for 10 weeks. VSL#3 treatment of chronic colitis rats, as for acute colitis rats, began 1 week before colitis induction and continued until time of sacrifice, with VSL#3 administered ad libitum dissolved in drinking water.
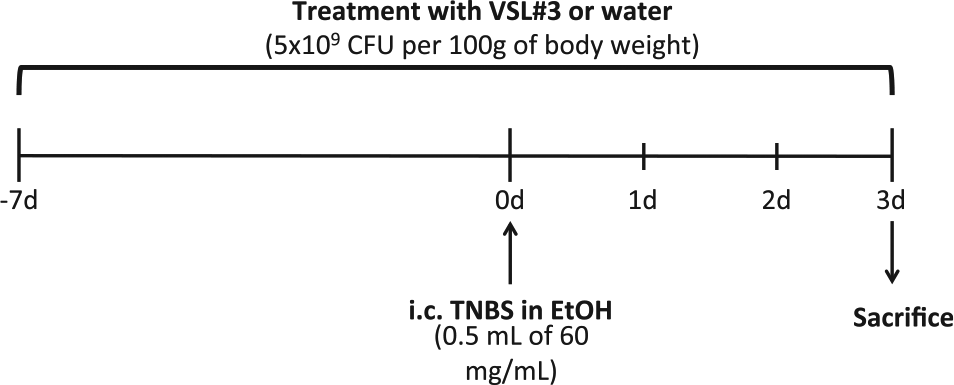
Probiotic treatment and colitis induction protocol. Abbreviation: TNBS, trinitrobenzene sulfonic acid; EtOH, ethanol.
Sample Collection
At the time of sacrifice, blood was collected by cardiac puncture, and the colon was resected. The colon from each rat was opened longitudinally, assessed for macroscopic damage, and sectioned into proximal, mid, and distal thirds. Each third was subsequently bisected longitudinally into equal parts, one for histological analysis and one for molecular analysis. Samples of liver, spleen, and terminal ileum were also collected for future studies. Tissue samples for histological analyses were fixed in formalin and embedded in paraffin as previously described, 37 whereas tissue samples for molecular analysis were snap frozen in liquid nitrogen and stored at −80C.
Macroscopic and Microscopic Damage Scoring
Macroscopic and microscopic damage for each colon was assessed using previously described scoring systems.31,35,36 The macroscopic damage scoring system evaluates colonic thickness (mm) and the presence of adhesions (none to major, 0–2), diarrhea (none or present, 0–1), and ulceration (none to extensive, 0–10). The values for each of these four criteria are added to obtain the total macroscopic damage score. The microscopic damage score evaluates preservation of mucosal architecture (preserved to severe loss, 0–3), location of inflammatory cell infiltration (none, 0; in muscularis mucosa, 1; in lamina propria, 2; in serosa, 3), thickness of the muscularis propria (<½ of mucosal thickness, 0; ½–¾ of mucosal thickness, 1; muscle = mucosal thickness, 2; muscle > mucosal thickness, 3; all muscle, 4), depletion of goblet cells (absent or present, 0–1), and presence of crypt abscess (absent or present, 0–1). As with the macroscopic damage score, the sum of the values for these five criteria yields the total microscopic damage score. For each rat, a microscopic damage score was obtained for the proximal, mid, and distal thirds based on the most severe findings present within each third.
Serum Cytokine Measurement
Serum from blood samples collected at time of sacrifice was used to measure systemic cytokine and chemokine levels. The rat cytokine/chemokine magnetic multiplex panel (RECYTMAG-65K, EMD Millipore) was used as per the manufacturer’s instructions to measure levels of the following cytokines (minimum detectable concentration, inter assay % coefficient of variation, % spike recovery): EGF (0.3 pg/mL, 6.3%, 88%), eotaxin (3.2 pg/mL, 9.9%, 95.3%), fractalkine (0.7 pg/mL, 6.4%, 93.4%), G-CSF (1.8 pg/mL, 11.5%, 99.3%), GM-CSF (6.8 pg/mL, 7.4% 93.0%), GRO/KC (19.7 pg/mL, 7.7%, 97.9%), IFN-γ (6.2 pg/mL 12.4%, 89.2%), IL-1α (4.2 pg/mL, 4.8%, 85.2%), IL-1β (2.8 pg/mL, 11.3%, 94.0%), IL-2 (5.4 pg/mL, 13.3%, 97.2%), IL-4 (3.1 pg/mL, 10.7%, 97.9%), IL-5 (7.4 pg/mL, 22.7%, 85.8%), IL-6 (30.7 pg/mL, 12.7%, 93.9%), IL-10 (2.7 pg/mL, 9.0%, 99.3%), IL-12p70 (3.3 pg/mL, 7.8%, 92.6%), IL-13 (2.4 pg/mL, 8.4%, 89.8%), IL-17A (2.3 pg/mL, 9.2%, 89.8%), IL-18 (6.2 pg/mL, 11.1%, 97.2%), IP-10 (1.4 pg/mL, 12.2%, 103.9%), leptin (10.2 pg/mL, 14.3%, 101.6%), LIX (20.9 pg/mL, 24.8%, 130.6%), MCP-1 (9.0 pg/mL, 9.2%, 80.2%), MIP-1α (0.8 pg/mL, 9.3%, 92.3%), MIP2 (11.3 pg/mL, 7.7%, 96.1%), RANTES (1.3 pg/mL, 17.1%, 95.7%), TNF-α (1.9 pg/mL, 10.8%, 86.0%), and VEGF (2.6 pg/mL, 6.0%, 92.1%). Each sample was run in duplicate.
Colonic Messenger Ribonucleic Acid (mRNA) Extraction and Real-Time Polymerase Chain Reaction (PCR)
Colonic samples for molecular analyses were collected at the time of sacrifice, snap frozen in liquid nitrogen, and stored at −80C. Thirty mg of proximal colon tissue were weighed and transferred to mRNAse free microcentrifuge tubes filled with beads and homogenizing solution. Tubes were transferred to a Bullet blender (Advance) and homogenized for 5 min. Nucleic acids and proteins were extracted from homogenized samples using the AllPrep deoxyribonucleic acid (DNA)/RNA/Protein Mini Kit (80004, Qiagen). The concentration and quality of extracted RNA was determined with a Nanodrop 2000 (Thermoforma). Then, 1 µg from the mRNA was used to generate complementary DNA with the iScript cDNA synthesis kit3 (1708891, Biorad). Real-time PCR was performed using iQ SYBR Green Supermix (1708882, Biorad) and the following primers (from Qiagen unless otherwise indicated): beta-actin, interleukin-1b (Il1b), claudin-5, chemokine c-c motif ligand 2 (Ccl2), occludin, interleukin-10 (Il10, IDT), interleukin-12 (Il12, IDT), and nitric oxide synthase 2 (Nos2, IDT). Data are reported as fold change using the 2−ΔCT equation.
Fecal Metagenomics and Microbiome Analysis
Fecal samples were collected after opening the colon under sterile conditions, transferred to sterile tubes, snap frozen, and stored at −80C. DNA was extracted from 100 mg of each fecal sample with the QIAamp DNA Stool Minikit (51504, Qiagen), and DNA concentration was determined with a Nanodrop 2000 (Thermoforma). The V1-V3 ribosomal region was amplified by PCR and the resulting product was analyzed via agarose gel. Samples were then sequenced with Illumina MiSeq, which provides the representative sequences of each Operational Taxonomic Unit (OTU). An OTU table was created to assess bacterial taxonomy and determine percent of bacterial composition. Closed reference OTU was used in all cases except Odoribacteraceae at the family level and Ruminococcus at the genus levels, for which open reference OTU was used (indicated by brackets).
Macrophage Staining and Quantification
Total and M1 colonic macrophages were stained by sequential double immunofluorescence for CD68 (MCA341B, AbD Serotec) and inducible nitric oxide synthase (iNOS; sc-7271, Santa Cruz), as previously described. 37 For each rat, two representative high power fields (HPF, 0.523 mm2) of mucosa, submucosa, and areas of mucosa or submucosa with positive staining for iNOS were photographed. Total macrophages (CD68+ cells) and M1 macrophages (CD68+iNOS+) were quantified using the cell counter plug-in on ImageJ v1.48i (NIH).
For tissue from our previously reported chronic colitis model, 31 tissues were stained for CD68 (AbD Serotec), and mucosal macrophages were quantified with ImageJ in 25 HPFs taken from a total of 3 normal age-matched rats, 39 HPFs taken from a total of 10 chronic colitis rats, and 38 HPFs taken from a total of 9 chronic colitis+VSL#3 rats.
Hepatic macrophages and proliferating cells were stained by sequential double immunofluorescence for CD68 (AbD Serotec) and Ki67 (550609, BD Pharmingen), as previously described. 37 Two representative HPFs of hepatic parenchyma surrounding central veins and of parenchyma surrounding portal areas were photographed for each rat. Total macrophages (CD68+ cells), proliferating cells (Ki67+ cells), and CD68+Ki67+ cells were quantified using ImageJ.
Statistical Analysis
All data are presented as mean ± standard error of the mean and were analyzed statistically with Prism v6.0h (GraphPad Software, Inc.). The three groups were compared via one-way analysis of variance with Tukey’s post hoc test.
Results
Treatment With VSL#3 Attenuates Colitis
Colitis induction caused a significant decrease in body weight in both the colitis and colitis+VSL#3 groups (p<0.001), in which rats lost nearly 20% of their precolitis body weight over the 72-hr postcolitis period, when compared with normal rats, which gained approximately 10% of their body weight during this same period (Fig. 2A). VSL#3 treatment did not have an effect on this weight loss. Average fluid intake per day of the protocol did not vary significantly or substantially between the three groups (Fig. 2B), with rats in the VSL#3-treated group consuming an average of 31.77 ± 2.69 106 CFU per gram of body weight each day. Rats in the colitis and colitis+VSL#3 groups had significantly higher macroscopic damage in the colon when compared with normal controls (p<0.001; Fig. 2C). Treatment with VSL#3 significantly reduced macroscopic damage in the colitic rats (p<0.05).
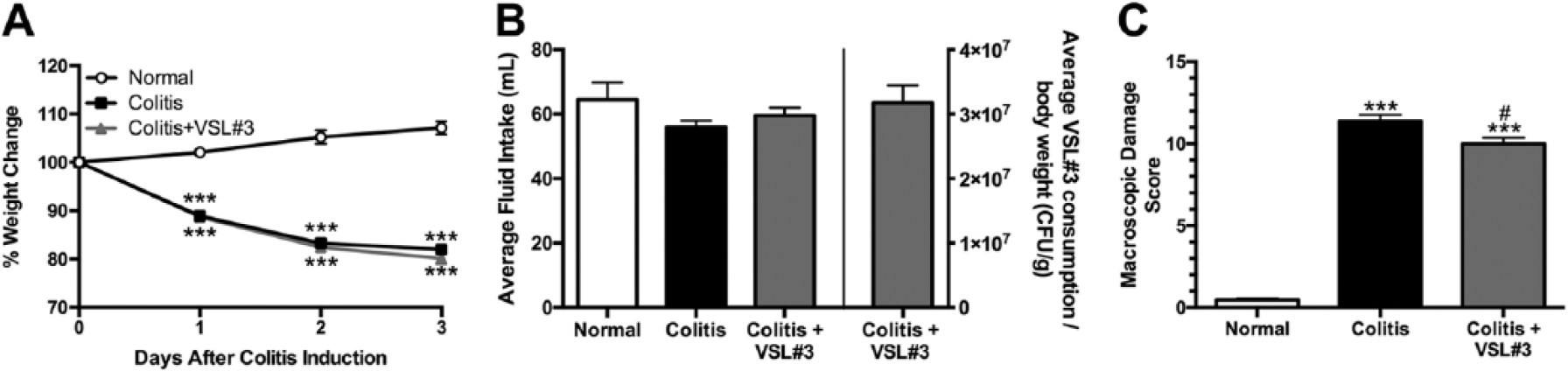
Macroscopic damage is reduced in rats treated with VSL#3. (A) weight change expressed as a percentage of body weight on the day colitis was induced for rats in colitis and colitis+VSL#3 groups. (B) average of daily fluid intake for all groups and of daily VSL#3 consumption per gram of body weight for the colitis+VSL#3 group. (C) Macroscopic damage scores. ***p<0.001 versus normal group, #p<0.05 versus colitis group. n=5–12 rats per group.
As expected, colitis induction produced increased microscopic damage in the proximal, mid, and distal colon when compared with normal. In the proximal colon, colitis and colitis+VSL#3 rats exhibited higher microscopic damage scores than normal rats (p<0.001 and p<0.05, respectively), and VSL#3 treatment significantly reduced microscopic damage (p<0.01; Fig. 3A). In the mid and distal colon, higher microscopic damage scores were observed for rats in the colitis group and the colitis+VSL#3 group when compared with the normal group (p<0.001), but these damage scores were not affected by VSL#3 treatment (Fig. 3B and C). Representative micrographs of histological findings in proximal colon are shown in Fig. 3D to F.
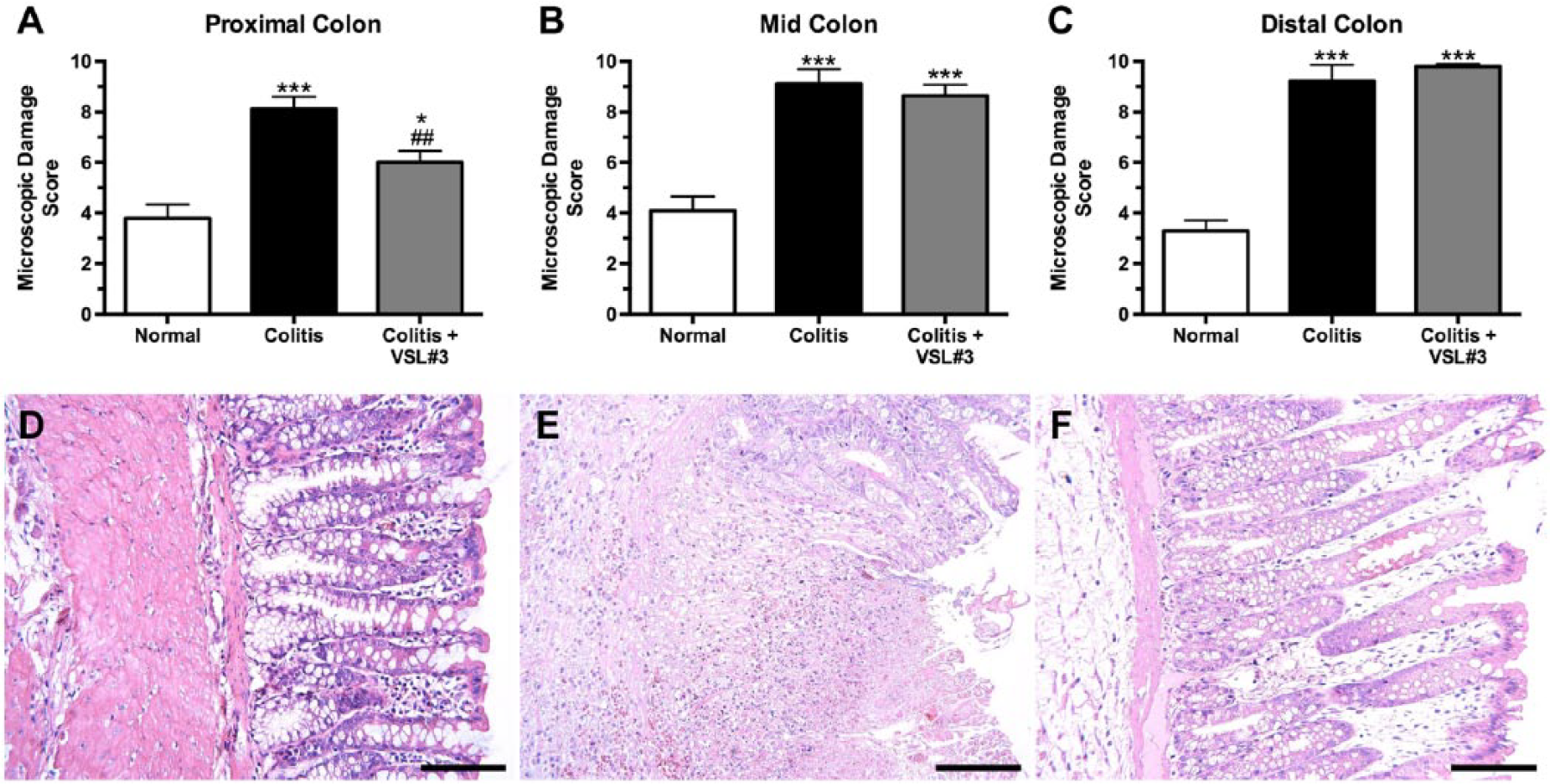
Microscopic damage in the proximal colon is decreased in rats treated with VSL#3. Microscopic damage in the proximal (A), mid (B), and distal (C) colon. Representative micrographs of histological findings in proximal colon from normal (D), colitis (E), and colitis+VSL#3 (F) rats. Hematoxylin and eosin staining. Scale bars = 100 µm. *p<0.05, ***p<0.001 versus normal group, ##p<0.01 versus colitis group. n=5–12 rats per group.
Administration of VSL#3 Reduces Serum Cytokine and Chemokine Levels
Serum cytokine and chemokine levels were measured to determine whether VSL#3 treatment altered the systemic inflammatory profile in colitic rats (Fig. 4). In general, treatment with VSL#3 had a tendency to bring cytokine and chemokine levels toward the baseline levels observed in normal rats. Colitis treatment alone significantly increased levels of EGF, GRO/KC, IL-5, IL-12p70, IL-13, IL-17A, IL-18, MCP-1/CCL-2, MIP-1α, TNF-α, and VEGF when compared with normal controls. Levels of GRO/KC, MCP-1/CCL-2, and VEGF were also significantly elevated in VSL#3-treated rats when compared with normal controls, but to a lesser degree than rats with colitis alone. VSL#3 treatment significantly reduced levels of EGF, eotaxin, IL-2, and VEGF when compared with colitis alone. Levels of leptin were significantly lower in colitis and colitis+VSL#3 rats when compared with normal controls.
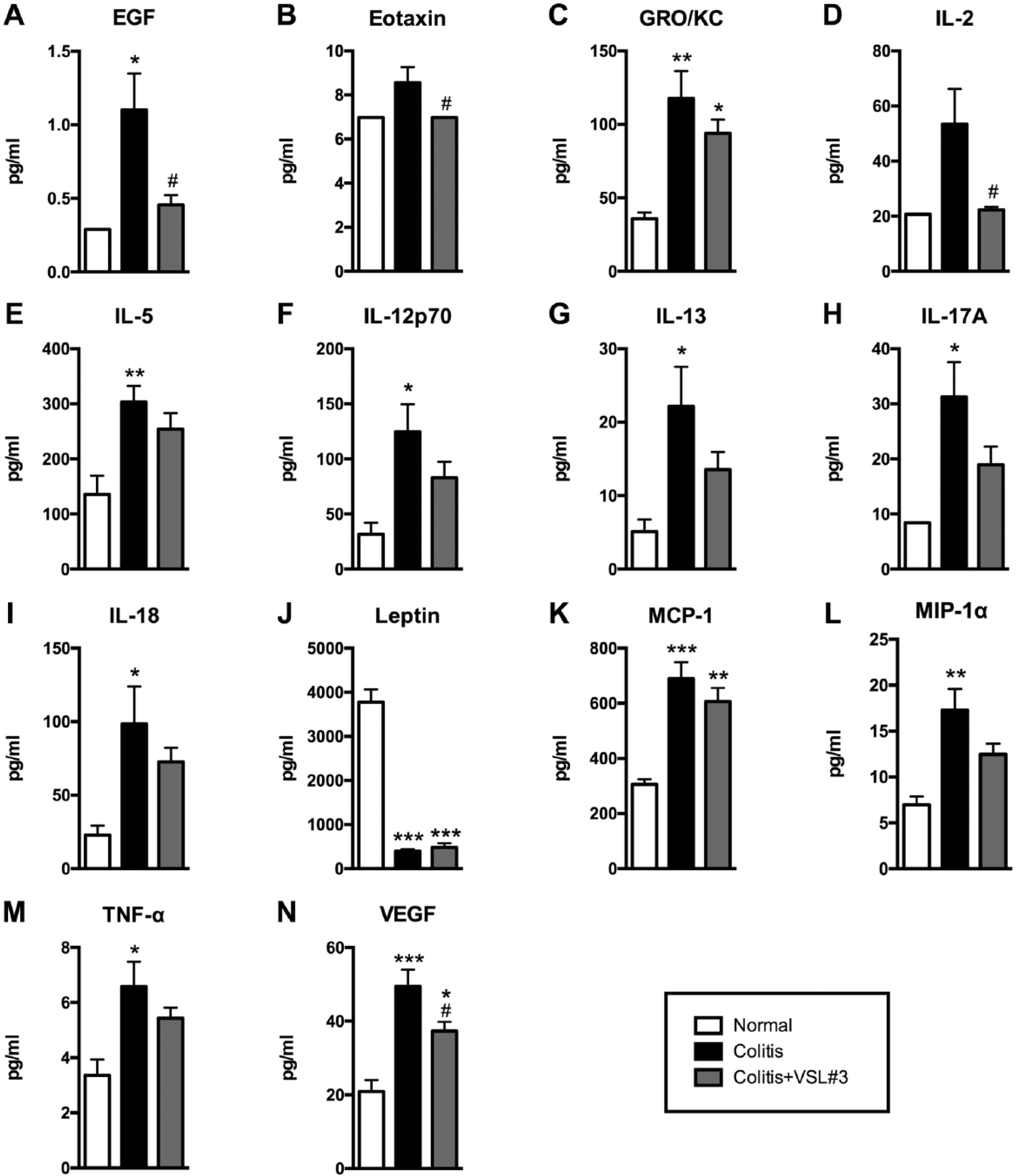
Serum cytokine and chemokine levels are modulated by treatment with VSL#3. Levels of EGF (A), eotaxin (B), GRO/KC (C), IL-2 (D), IL-5 (E), IL-12p70 (F), IL-13 (G), IL-17A (H), IL-18 (I), leptin (J), MCP-1 (K), MIP-1α (L), TNF-α (M), VEGF (N) in serum from normal, colitis, and colitis+VSL#3 rats, as determined by multiplex bead array. *p<0.05, **p<0.01, ***p<0.001 versus normal group, #p<0.05 versus colitis group. n=5–12 rats per group.
VSL#3 Modulates Transcript Levels of Colonic Cytokines and Tight Junction Proteins
Molecular analyses of proximal colon tissue was undertaken to assess whether VSL#3 also modulates the local inflammatory milieu and epithelial barrier integrity in colitic rats, particularly in the colonic segment in which VSL#3 had a significant effect (Fig. 5). The only statistically significant change observed was for relative Il12 expression, which was significantly increased in the colitis+VSL#3 group when compared with the colitis group (Fig. 5B). Nevertheless, the following statistically nonsignificant changes were observed relative to normal transcript levels: colitis induction decreased transcript levels for all the pro-inflammatory factors measured (Il1b, Il12, Nos2, Ccl2/Mcp-1), the anti-inflammatory cytokine Il10, and the tight junction proteins Occludin and Claudin-5; whereas VSL#3 treatment increased transcript levels for all analytes except Il1b.
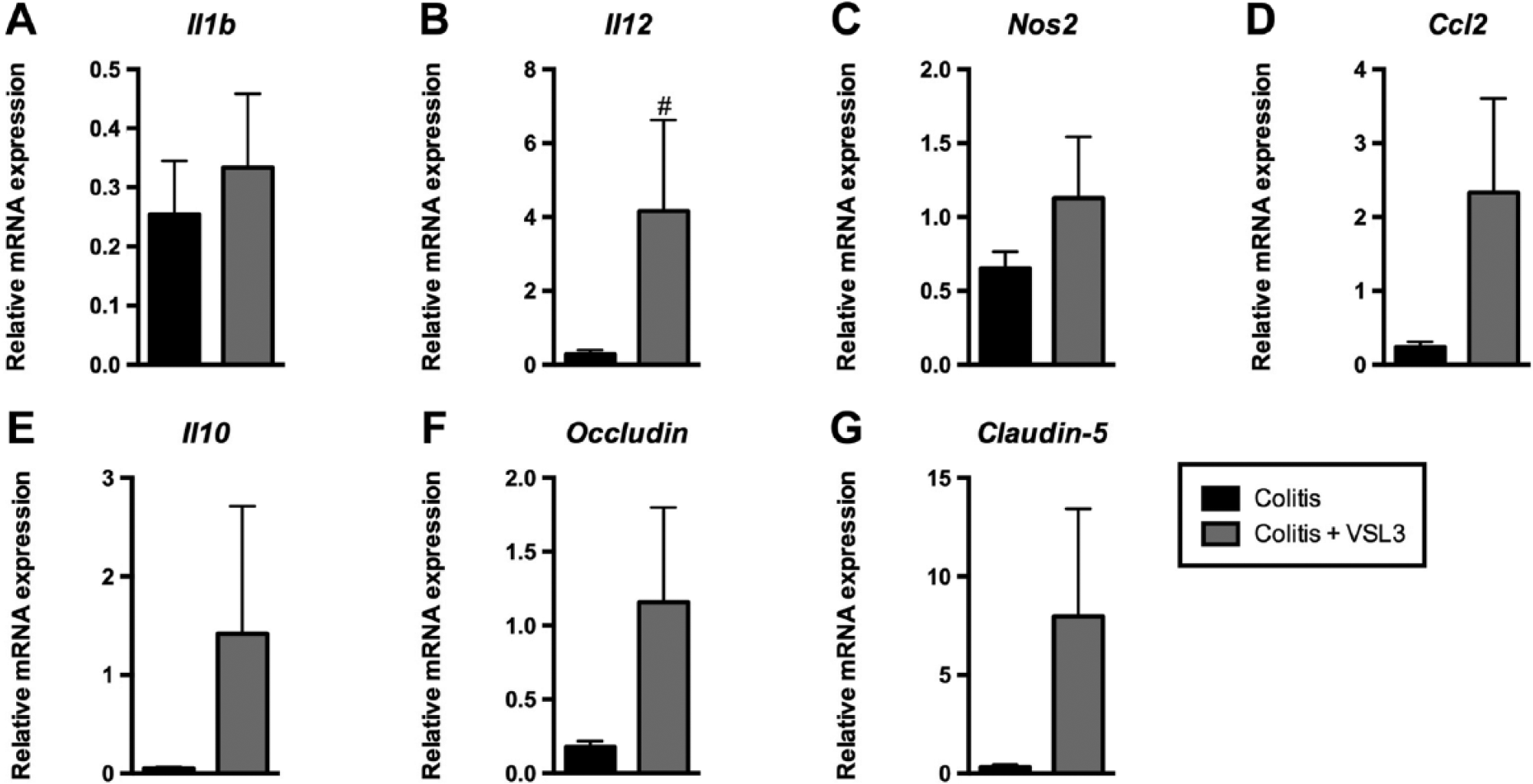
Colonic transcript levels for cytokines and tight junction proteins are modulated by treatment with VSL#3. Fold change in mRNA expression of Il1b (A), Il12 (B), Nos2 (C), Ccl2 (D), Il10 (E), Occludin (F), and Claudin-5 (G) as compared with beta-actin in proximal colonic tissue lysates from normal, colitis, and colitis+VSL#3 rats, as determined by real-time PCR and using the 2−ΔCT equation. Data normalized to normal rats. #p<0.05 versus colitis group. n=5–6 rats per group. Abbreviations: PCR, polymerase chain reaction; Il1b, interleukin-1b; Il12, interleukin-12; Nos2, nitric oxide synthase 2; Ccl2, chemokine c-c motif ligand 2; Il10, interleukin-10.
Treatment With VSL#3 Alters the Composition of Fecal Microflora
Previous studies have shown that VSL#3 treatment leads to changes in the intestinal flora both in patients with IBD and in several animal models of IBD. However, we wanted to assess whether VSL#3 treatment starting a week before, and continuing for 3 days after, colitis onset could alter the composition of the fecal microflora in our animals. Metagenomic analyses of the fecal microflora demonstrated marked differences in the percent distribution of bacteria contributing to fecal microflora from phylum to genus taxonomic levels (Fig. 6). Individual comparisons revealed the following: colitis, with or without VSL#3 treatment, caused statistically significant increases in Porphyromonodacea and [Odoribacteraceae] at the family level (p<0.01) and Parabacteroides and Butyricimonas at the genus level (p<0.01) and decreases in Firmicutes at the phylum level (p<0.01), Clostridia at the class level (p<0.001), Clostridiales at the order level (p<0.001), Clostridaceae and Lachnospiraceae at the family level (p<0.01), and SMB53 (p<0.01), Coprococcus (p<0.05), and [Ruminococcus] (p<0.001) at the genus level when compared with normal. Colitis without VSL#3 treatment caused statistically significant (p<0.05) increases in Bacteroidetes at the phylum level, Bacteroidia and Erysipelotrichi at the class level, Bacteroidales and Erysipelotrichales at the order level, Erysipelotrichaceae at the family level, and Allobaculum at the genus level when compared with normal. VSL#3 treatment in the setting of colitis caused statistically significant increases in Bacteroidaceae at the family level and Bacteroides at the genus level when compared with normal (p<0.05) and in Clostridium at the genus level when compared with normal and colitis alone (p<0.05), and decreases in Prevotellaceae at the family level and Prevotella at the genus level when compared with colitis alone (p<0.01). VSL#3 treatment also increased Verrucomicrobia at the phylum level, Verrucomicrobiae at class level, Verrucomicrobiales at the order level, Verrucomicrobiaceae at the family level, and Akkermansia at the genus level when compared with normal (p=0.057) and colitis alone (p=0.054).
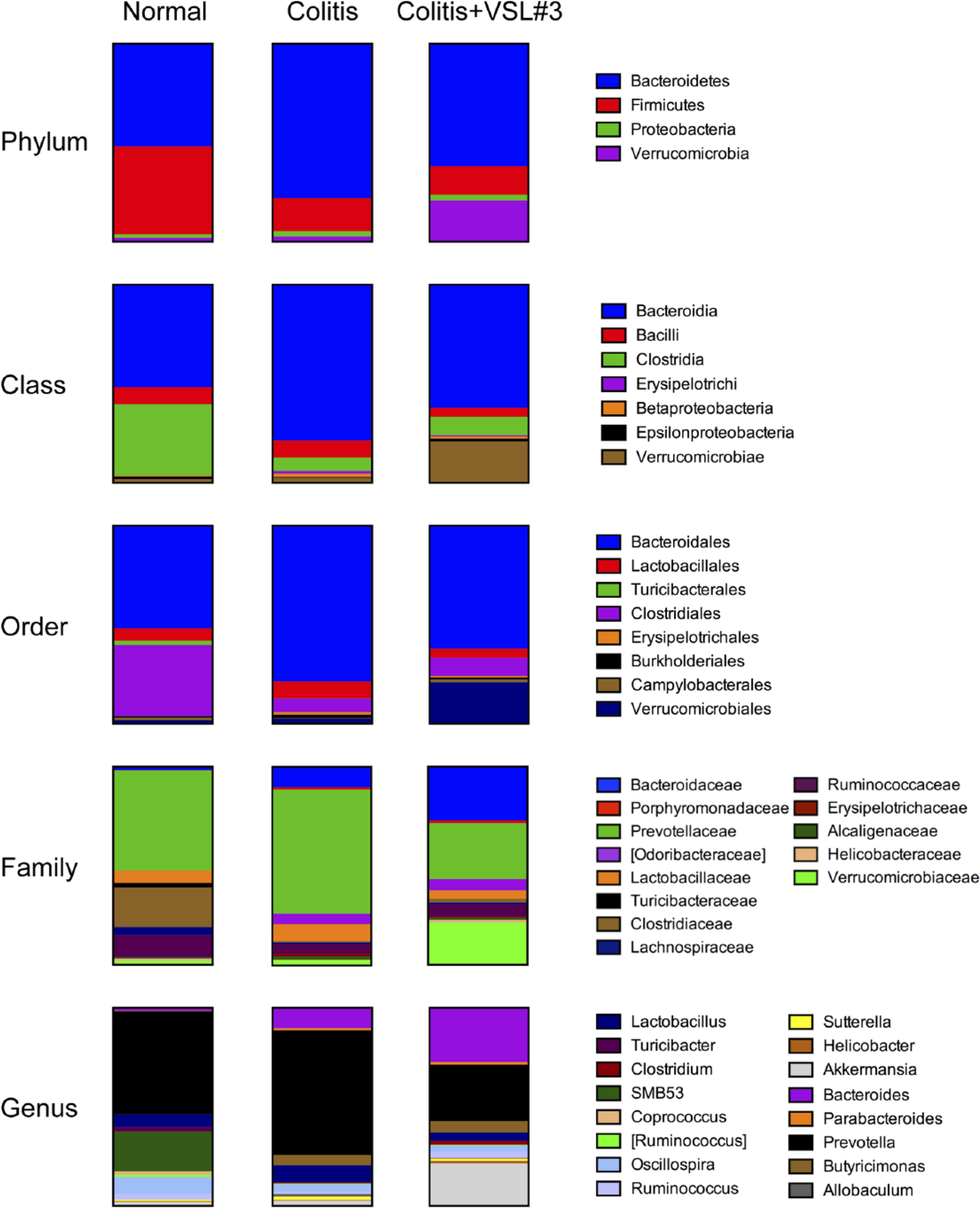
Fecal microflora composition is altered by treatment with VSL#3. Percent microflora composition at the phylum, class, order, family, and genus levels, as determined by fecal metagenomics. n=5–6 rats per group.
VSL#3 Decreases Macrophage Infiltration in Acute, but Not Chronic, Colitis
In light of the protective effect observed for VSL#3 on microscopic damage in this colonic segment, macrophage phenotype and infiltration analyses focused on the proximal colon. M1 macrophages (iNOS+CD68+ cells), as identified by double immunofluorescence, were mostly observed within mucosal and submucosal areas that stained positively for iNOS and were scarce elsewhere. Within these areas of mucosa and submucosa, the percentage of macrophages that were of the M1 phenotype was significantly increased in the colitis and colitis+VSL#3 groups when compared with normal (p<0.05), but was not affected by VSL#3 treatment (Fig. 7A). The number of M1 macrophages was significantly increased in the colitis group when compared with normal controls (p<0.001; Fig. 7B), and this was not significantly decreased by VSL#3. The total number of macrophages (CD68+ cells) within these iNOS-positive areas was comparable between the normal, colitis, and colitis+VSL#3 groups (Fig. 7C). In the normal group, iNOS staining was present in the submucosa (Fig. 7D). In the colitis and colitis+VSL#3 groups, iNOS was also present in the submucosa underlying undamaged/intact mucosa as well as in mucosal and submucosal areas of inflammation and necrosis (Fig. 7E and F).
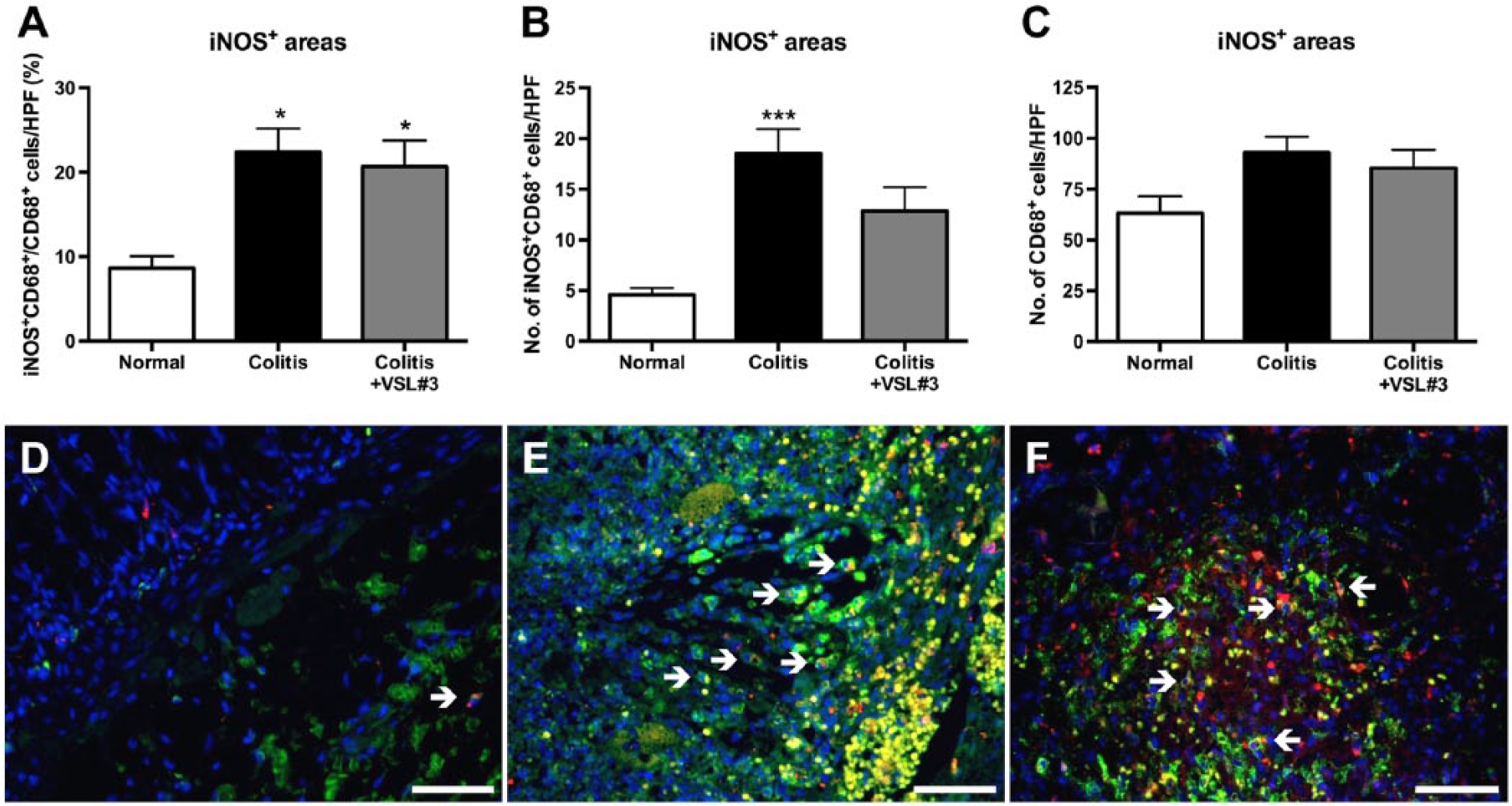
Treatment with VSL#3 does not reduce the percentage of M1 macrophages. Sections of proximal colon were stained by double immunofluorescence for CD68 and inducible nitric oxide synthase (iNOS). (A) percentage of CD68+ cells (macrophages) that are iNOS+CD68+ (M1 macrophages, white arrows in D-E) per high power field (HPF=0.523 mm2) of mucosa or submucosa staining positively for iNOS. (B) number of iNOS+CD68+ cells per HPF of mucosa or submucosa staining positively for iNOS. (C) number of CD68+ cells per HPF of mucosa or submucosa staining positively for iNOS. Representative micrographs of CD68 (red), iNOS (green), and 4’,6-diamidino-2-phenylindole (DAPI, blue) in colonic tissue from normal (D), colitis (E), and colitis+VSL#3 (F) rats. Scale bars = 50 µm. *p<0.05, ***p<0.001 versus normal group. n=10–20 HPFs per group.
Treatment with VSL#3 significantly reduced the mucosal and submucosal macrophage infiltration resulting from the induction of colitis. Mucosal macrophage numbers were significantly increased in the colitis group when compared with the normal group (p<0.001) and significantly reduced in the colitis+VSL#3 group (p<0.01) when compared with the colitis group (Fig. 8A, C–E). The number of submucosal macrophages was significantly higher in colitis rats when compared with normal rats (p<0.001; Fig. 8B, F–H). Submucosal macrophage numbers were significantly lower in the colitis+VSL#3 group than in the colitis group (p<0.01), but these numbers were still significantly increased in the colitis+VSL#3 group when compared with normal rats (p<0.01).
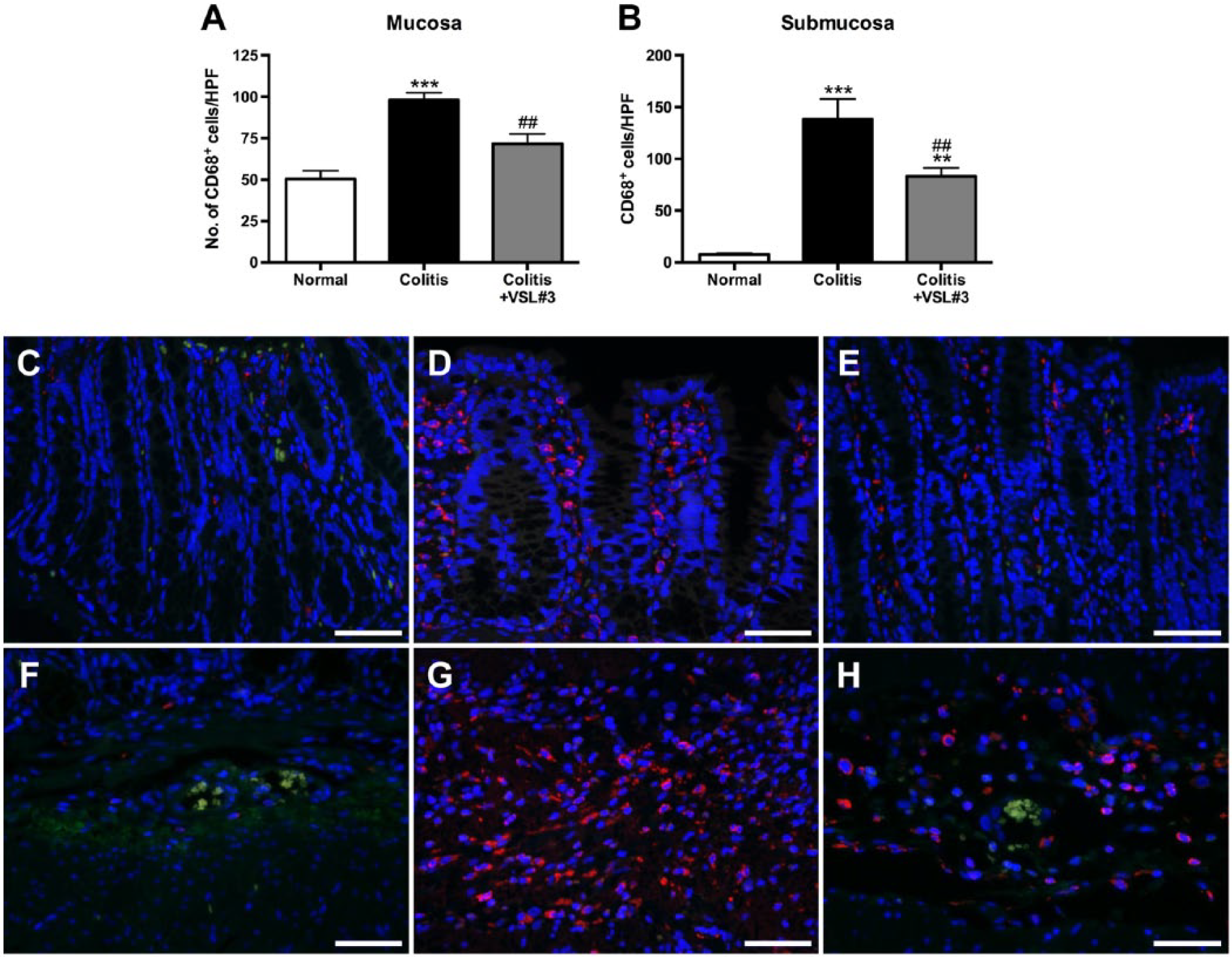
Treatment with VSL#3 decreases mucosal and submucosal macrophage infiltration. Sections of proximal colon were stained by double immunofluorescence for CD68 and inducible nitric oxide synthase (iNOS). Note the paucity of iNOS+ cells. Number of CD68+ cells per high power field (HPF=0.523 mm2) of mucosa (A) or submucosa (B). Representative micrographs of CD68 (red), iNOS (green), and 4’,6-diamidino-2-phenylindole (DAPI, blue) in mucosa from normal (C), colitis (D), and colitis+VSL#3 (E) rats and in submucosa from normal (F), colitis (G), and colitis+VSL#3 (H) rats. Scale bars = 50 µm. **p<0.01, *** p<0.001 versus normal group, ##p<0.01 versus colitis group. n=10–24 HPFs per group per area.
Given the VSL#3-induced reduction in mucosal and submucosal macrophages in our acute colitis model, we asked whether this probiotic also mod-ulated macrophage infiltration in our previously reported chronic colitis model (Appleyard et al., 2011). In colonic tissue from our previously reported chronic colitis model, macrophage numbers did not differ significantly between age-matched normal rats (65.48 ± 4.90 cells per HPF), chronic colitis rats (59.97 ± 3.19 cells per HPF), and chronic colitis+VSL#3 rats (63.47 ± 3.58 cells per HPF; Fig. 9A). Differences were not observed when data were stratified by histopathological diagnosis (Fig. 9B), as macrophage numbers in tissue from chronic colitis rats and chronic colitis+VSL#3 rats contained similar numbers of macrophages in areas of IBD (70.26 ± 6.33 and 63.42 ± 3.45 cells per HPF, respectively), dysplasia (53.24 ± 2.85 and 71.20 ± 7.32 cells per HPF, respectively), and cancer (77.60 ± 21.38 and 52 ± 15.00 cells per HPF).
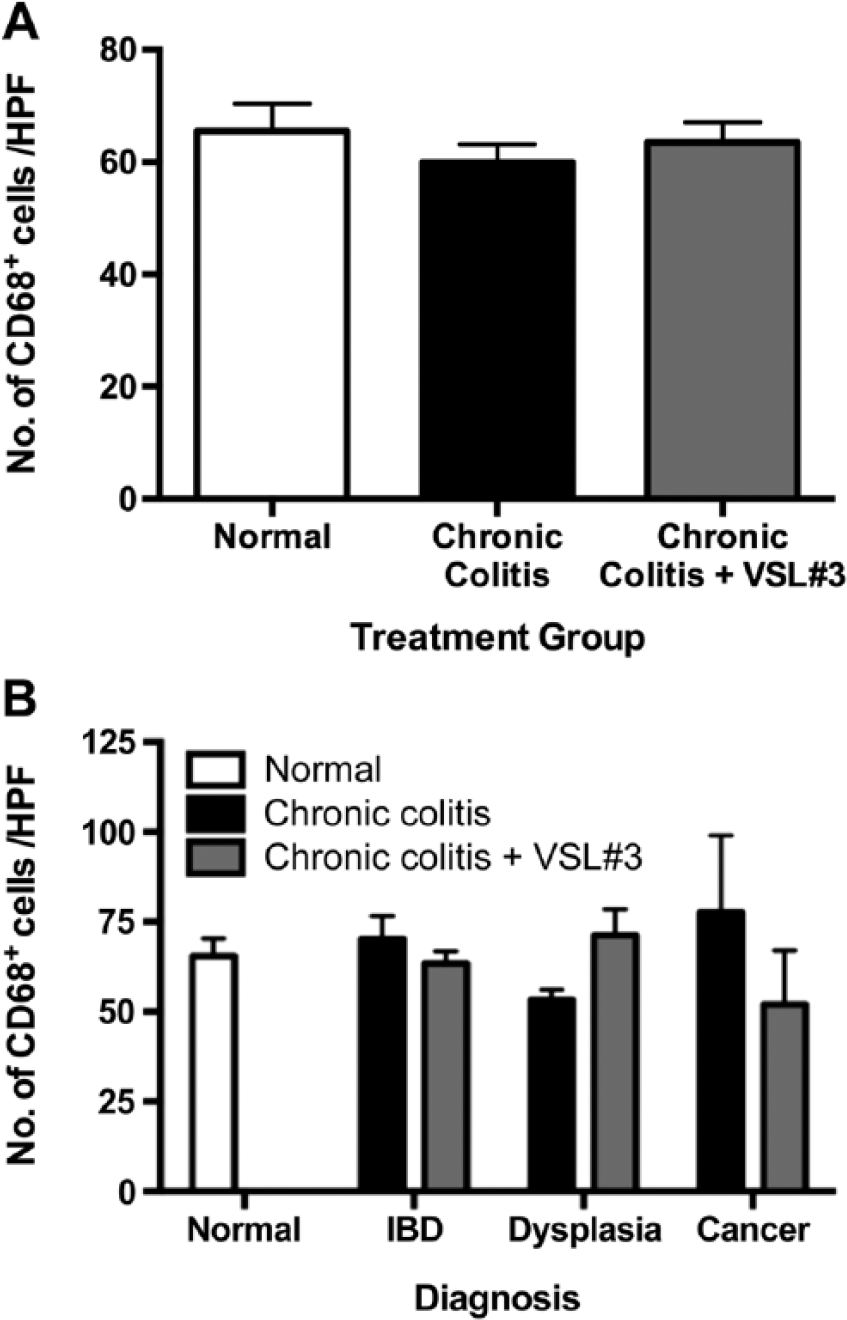
Chronic colitis alone or in combination with VSL#3 treatment does not increase mucosal macrophage numbers. Colonic sections were stained by immunofluorescence for CD68. (A) number of CD68+ cells per high power field (HPF=0.523 mm2) of mucosa for each treatment group (n=25–39 HPFs per group). (B) number of CD68+ cells per HPF (HPF=0.523 mm2) of mucosa for each treatment group stratified by histopathological diagnosis (n=26 normal HPFs, 19–20 IBD HPFs, 17–20 dysplasia HPFs, and 2–5 cancer HPFs). Abbreviation: IBD, inflammatory bowel disease.
Liver Macrophage Accumulation Is Not Affected by VSL#3 Treatment
Macrophages have recently been shown to accumulate in the liver in several models of colitis. 38 We, therefore, asked whether this accumulation could be attenuated by treatment with VSL#3 given the inhibitory effect VSL#3 had on colonic macrophage infiltration. Macrophage numbers in hepatic parenchyma surrounding central veins and portal areas significantly increased in colitis and colitis+VSL#3 rats in comparison with normal rats (p<0.01 and p<0.05, respectively; Fig. 10A). Positivity for the proliferative marker Ki67 was used to determine whether the macrophage accumulation was a result of increased local macrophage proliferation. The percentage of proliferating cells attributable to macrophages was increased in colitis and colitis+VSL#3 groups in hepatic parenchyma surrounding central veins, but not portal areas, when compared with normal (p<0.05; Fig. 10B). However, the vast majority of macrophages in all groups were not actively proliferating (not shown).
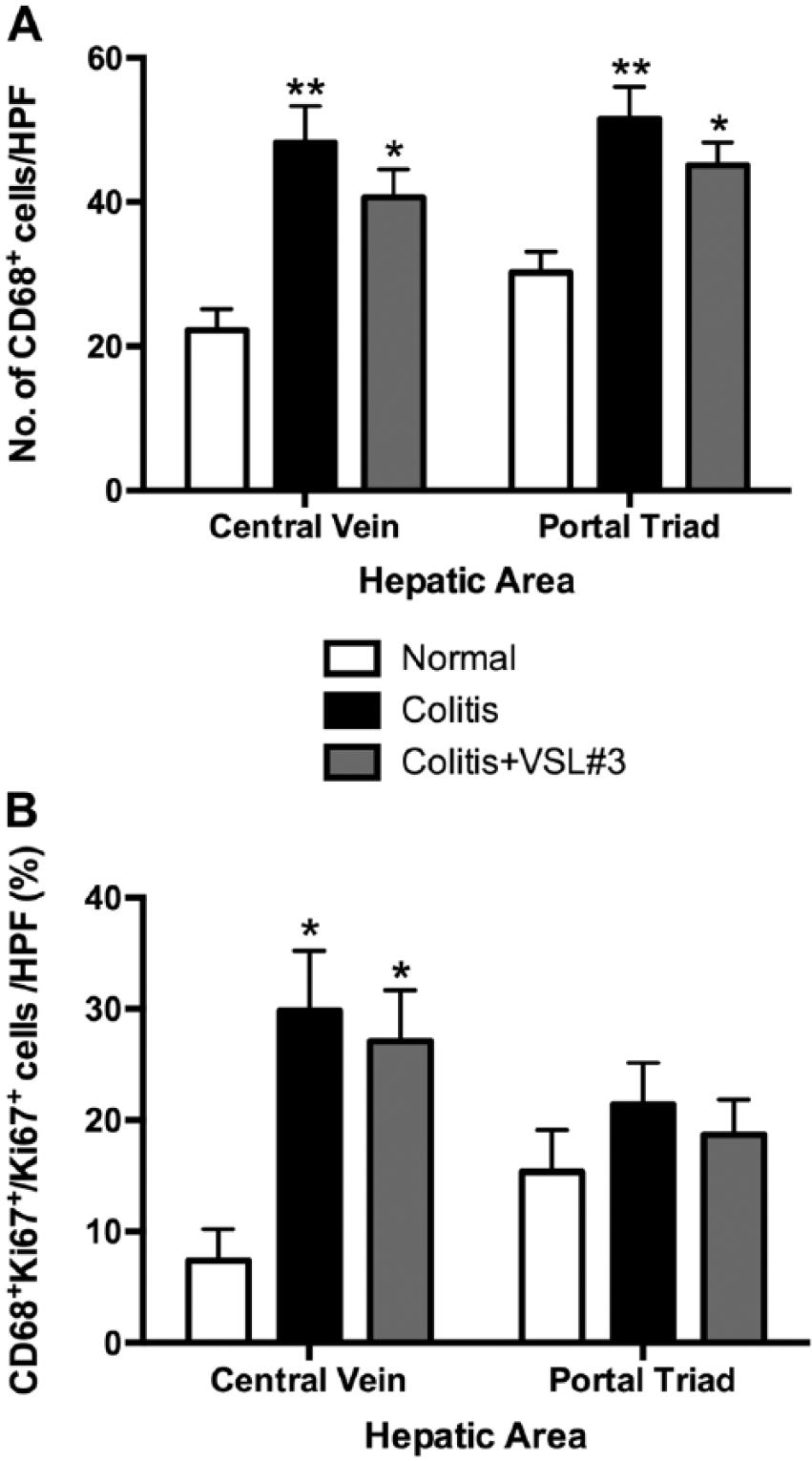
VSL#3 treatment does not affect colitis-induced macrophage accumulation in the liver. Liver sections were stained by double immunofluorescence for CD68 and Ki67. (A) number of CD68+ cells (macrophages) per high power field (HPF=0.523 mm2) of liver parenchyma surrounding central veins or portal areas. (B) percentage of Ki67+ cells (proliferating cells) that are CD68+Ki67+ (macrophages) per HPF of liver parenchyma surrounding central veins or portal areas. *p<0.05, **p<0.01 versus normal group. n=10–21 central vein HPFs, 10–24 portal triad HPFs per group.
Discussion
In the present study we have shown that treatment with the probiotic mixture VSL#3 in acute TNBS-induced colitis leads to the attenuation of colitis, a reduction in serum cytokine and chemokine levels, modification of the intestinal flora, and diminishment of colonic macrophage infiltration. However, VSL#3 treatment did not decrease the percentage of colonic macrophages with an M1 phenotype, as determined by double immunofluorescence for CD68 and iNOS. This is consistent with our previous finding that M1 macrophages maintained their pro-inflammatory phenotype despite exposure to VSL#3 in vitro. 30 However, this contrasts with a previous report that VSL#3 treatment reduces colonic M1 macrophages, defined as F4/80+MCP-1+ cells. 29 It is worth pointing out that the prevalence of F4/80+MCP-1+ cells in the aforementioned study did not correlate with histopathological changes between mice expressing or deficient in PPAR-γ treated with VSL#3, that F4/80 staining is more prevalent on resident, M2-like colonic macrophages than in pro-inflammatory M1 macrophages,12,15 and that MCP-1 is not routinely used to identify M1 macrophages in the mouse. 39 We, therefore, conclude that the combined current evidence does not support the reduction of M1 macrophages as a mechanism of action for the anticolitic effect of VSL#3 in animal models of colitis. In light of these findings and to improve the treatments available for patients with IBD, future studies, should, therefore explore the possibility of combining the probiotic VSL#3 with other treatments that do reduce the pro-inflammatory phenotype of M1 macrophages and/or augment the anti-inflammatory phenotype of M2 macrophages in hopes of obtaining a synergistic effect between the two treatments. Possible candidates include other probiotics, such as Clostridium butyricum 7 ; existing treatments, such as anti-TNF-α antibodies40,41; and new treatments, such as adoptive transfer.42,43
Here, we have shown that the VSL#3-induced reduction in colonic damage was most evident at the histological level in the proximal colon. There are several factors that could contribute to this finding. The colonic microbial population is higher near the rectum and anus than it is near the ileocecal valve. As dictated by the anatomy and physiology of the gastrointestinal tract, intestinal contents first reach the colon through the ileocecal valve before sequentially traversing the different colonic segments and reaching the rectum. Therefore, it is possible that the proximal-most segments of the colon are exposed to higher concentration of the probiotic, its products, and/or components than the distal-most colonic segments. Although it is possible that the most proximal portions of the colon are the sites where TNBS dwells the least, we do not feel that this contributes to decreased proximal colon injury given that average microscopic damage scores for untreated colitis rats were comparable across the proximal, middle, and distal colonic segments.
We have previously demonstrated that VSL#3 ameliorates inflammation and delays the transition from chronic inflammation to neoplasia in chronic TNBS-induced colitis in rats. 31 This chemopreventive effect has subsequently been confirmed by others.44,45 Interestingly, in the current study, we found that mucosal macrophages in the colon of rats with chronic colitis with or without VSL#3 treatment and of age-matched normal rats, all from our previous study, did not differ in number. Given that the colitis protocol for this chronic model begins with an acute colitis induced identically to the colitis protocol used in the current study, it is likely that the macrophage infiltration that we observed 72 hr after colitis induction dissipates over time, despite prolonged reactivation of colitis via intravenous administration of TNBS. Treatment with VSL#3 in this chronic colitis model does not significantly affect macrophage numbers in the colonic mucosa. There are some differences, however, between the animals in the present study and those of our prior study that should be noted. Rats in the VSL#3-treated group consumed an average of 31.77 ± 2.69 × 106 CFU per gram of body weight each day, a significantly higher amount of probiotic than the average 21.30 ± 0.83 × 106 CFU per gram of body weight consumed daily by rats in our previous study using a chronic colitis model for the same early time points. 31 Interestingly, VSL#3-treated and untreated rats in our prior study lost approximately 10% of their precolitis body weight 72 hr after colitis induction, in contrast with the almost 20% reduction observed in the present study and suggesting that the colitis in the present study was more severe than that of our previous study. This difference is intriguing given that the rats in the current study were, on average, heavier than those in the previous study both at the start of the study protocol and on the day of colitis induction, and that, as previously mentioned, rats in the current study consumed more VSL#3 per body weight than those in our prior study. Nevertheless, in both the previous and present studies, treatment with VSL#3 did not significantly reduce weight loss over the 72 hr following colitis induction. Another reason that could account for the different patterns of colonic macrophage infiltration between the acute colitis and chronic colitis rats is that the chronic colitis reactivation in the chronic model causes the inflammatory milieu to evolve in such a way that results in a normalization of macrophage recruitment to the colon and that is dependent on other subsets of immune cells. Future studies should focus on the dynamics of the inflammatory response in the progression toward the chronic inflammatory state in this model of chronic colitis.
Our findings with regard to serum cytokine and chemokine levels shed light on the inflammatory profile of acute TNBS colitis in the rat. The significant elevation of serum levels for IL-12p70, IL-5, and IL-13, and IL-17A suggest that Th1, Th2, and Th17 helper T cells, respectively, are involved in colitis development at this early time point. Interestingly, VSL#3 decreased serum levels of IL-2, a cytokine important for T-cell activation, and levels for IL-12p70, IL-5, IL-13, and IL-17A were not significantly elevated in VSL#3-treated rats, suggesting that VSL#3 treatment decreased the activation of Th1, Th2, and Th17 helper T cells. Curiously, VSL#3 treatment not only reduced levels of IL-2 and the chemokine eotaxin but also decreased the levels of the pro-healing factors EGF and VEGF. Although this seems somewhat paradoxical, we believe that this is attributable to milder colitis that developed in VSL#3-treated rats in comparison with untreated rats. This notion is supported by the general tendency for lower serum cytokine and chemokine levels, with the exception of leptin, in colitic rats treated with VSL#3 compared with untreated colitic rats along with the reduction in macroscopic damage, microscopic damage, and macrophage infiltration observed in VSL#3-treated rats. The VSL#3-induced shift of cytokine and chemokine levels toward those of normal controls is consistent with previously reported studies. 46 This explanation is also supported by the VSL#3-induced restoration of colonic transcript levels for pro-inflammatory, anti-inflammatory, and barrier proteins to or past normal levels that was observed. Whereas this restorative pattern is consistent between protein-level analyses in serum and the transcript-level analyses in the colon, it is intriguing that expression patterns for certain factors, specifically IL-12 and CCL2, are discordant at the protein and transcript levels. Two possible explanations for this observation are as follows: one, that the sources of the serum cytokine and chemokine protein levels include sites other than the colon (e.g., mesenteric lymph nodes, liver), as occurs in acute phase reactions; and two, that the different inflammatory states induced by colitis alone or in combination with VSL#3 treatment alter the processing and half-life for colonic transcripts.
The colitis-induced accumulation of hepatic macrophage observed herein is consistent with previous studies. 38 Despite reducing systemic cytokine and chemokine levels, VSL#3 treatment did not reduce macrophage accumulation in the liver. The findings that proliferating macrophages were increased in hepatic parenchyma surrounding central veins but not portal triads, and that most hepatic macrophages were not actively proliferating, suggest that the accumulation of macrophages in the liver during colitis represents a combination of both local proliferation and infiltration of circulating cells.
Several studies in animals and humans have shown that VSL#3 can modify the composition of the gut microflora.29,31,47-58 Here, we show that even a 10-day course of VSL#3 administration can alter the fecal microflora. This is very relevant given that we still lack a full understanding of the impact of microflora composition changes on colon function during inflammation. As reviewed by Marchesi and others,59-61 the gut microflora is relatively stable during steady state conditions in healthy subjects, being mainly composed of the Firmicutes and Bacteroidetes phyla, with far lesser amounts of Proteobacteria and Actinobacteria phyla. However, under inflammatory conditions in IBD, the microflora is much more unstable with a dysbiosis occurring and a shift toward more potentially harmful bacteria, with many studies observing a reduction in microflora diversity. Differing reports have emerged but lower levels of Firmicutes are commonly found together with changes in Bacteroidetes. It is apparent, however, that the composition also varies dependent upon disease state, both over time and whether the condition is currently active or quiescent. Our analyses find that, in this model of acute TNBS colitis, VSL#3 treatment specifically increased the percent microbial composition attributable to members of the Akkermansia and Clostridium genus and decreased the percent microbial composition attributable to members of the Prevotella genus. Members of the Akkermansia genus, specifically A. muciniphila, have been suggested to enhance epithelial barrier integrity. 62 Certain members of the Clostridium genus have been shown to have protective effects against colitis.7,63 Members of the Prevotella genus are increased in the flora of patients with IBD.64,65 Therefore, one possible mechanism by which VSL#3 attenuated colitis in our animals was by the modulation of these bacteria.
In summary, the VSL#3 induced-changes that we have observed are likely due to a combination of several mechanisms, including the modification of the gut microflora,54,57 secretion of bacterial products such as conjugated linoleic acid 29 and lactocepin, 66 ligation of Toll-like receptor 9 by VSL#3 bacterial DNA, 67 and enhancement of the epithelial barrier.53,68,69 Whether either the presence of colonic macrophages or the inhibition of colonic macrophage infiltration is essential or important for mediating the beneficial effects of VSL#3 should be the subject of future investigations.
Footnotes
Acknowledgements
The authors acknowledge Ábner Vázquez, Siomara Hernández, Josué Cabán, Pablo López, Raphael Sanchez, Yasuhiro Yamamura, and Alcira Benitez Barros for technical assistance. Sigma Tau Pharmaceuticals generously donated the probiotic formulation VSL#3 that was used in this study. The authors would also like to thank the Ponce Health Sciences University (PHSU) Molecular Genomics Core Laboratory (RR003050/MD007579) for the support provided.
Author Contributions
RAI, AAI, and CBA designed the study. RAI, AL, MLC, MIGT, and GC performed the experiments. RAI analyzed the data. RAI drafted the manuscript. AL, MLC, MIGT, GC, AAI, and CBA critically revised the manuscript for important intellectual content. All authors have read and approved the final manuscript.
Competing Interest
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: RAI was supported by a William Townsend Porter Predoctoral Fellowship from the American Physiological Society and by the National Institute of General Medical Sciences (NIGMS) of the National Institutes of Health (NIH) R25GM082406. GC and CBA were supported in part by NIGMS-Institutional Development Award (IDeA) Networks of Biomedical Research Excellence (INBRE) P20 GM103475. AL and MIGT were supported in part by NIGMS R25GM096955.