
Abstract
Biologically relevant, three-dimensional extracellular matrix is an essential component of in vitro vasculogenesis models. WI-38 fibroblasts assemble a 3D matrix that induces endothelial tubulogenesis, but this model is challenged by fibroblast senescence and the inability to distinguish endothelial cell-derived matrix from matrix made by WI-38 fibroblasts. Matrices produced by hTERT-immortalized WI-38 recapitulated those produced by wild type fibroblasts. ECM fibrils were heavily populated by tenascin-C, fibronectin, and type VI collagen. Nearly half of the total type I collagen, but only a small fraction of the type IV collagen, were incorporated into ECM. Stable hTERT-WI-38 transfectants expressing TagRFP-fibronectin incorporated TagRFP into ~90% of the fibronectin in 3D matrices. TagRFP-fibronectin colocalized with tenascin-C and with type I collagen in a pattern that was similar to that seen in matrices from wild type WI-38. Human Umbilical Vein Endothelial Cells (HUVEC) formed 3D adhesions and tubes on WI38-hTERT-TagRFP-FN-derived matrices, and the TagRFP-fibronectin component of this new 3D human fibroblast matrix model facilitated the demonstration of concentrated membrane type 1 metalloprotease and new HUVEC FN and collagen type IV fibrils during EC tubulogenesis. These findings indicate that WI-38-hTERT- and WI-38-hTERT-TagRFP-FN-derived matrices provide platforms for the definition of new matrix assembly and remodeling events during vasculogenesis.
Introduction
Vasculogenesis plays critical roles in development, wound healing, and pathological events. Vasculogenesis involves endothelial cell (EC) migration, proliferation, and differentiation to form capillary tube-like structures, and these steps are modulated by the 3D environment (Schor et al. 2001). Most studies of new blood vessel formation by endothelia utilize gel-like structures with incompletely defined ECM proteins, growth factors and cytokines that are organized differently than those seen in natural matrix environments.
MatrigelTM is widely used for in vitro assays of vasculogenesis, and is derived from murine Englebreth-Holm-Swarm tumors (Kleinman and Martin 2005; Orkin et al. 1977). MatrigelTM contains matrix proteins including laminin, entactin, and collagen IV that provide an adhesive substrate for ECs during tubulogenesis; yet, these proteins are homogenized during extraction, and lack the infrastructural features of biological basement membranes. Furthermore, MatrigelTM incorporates many growth factors, including basic fibroblast growth factor, epidermal growth factor, insulin-like growth factor-1, transforming growth factor-β, platelet-derived growth factor, and nerve growth factor (Benton et al. 2009; Hughes et al. 2010; Vukicevic et al. 1992). It is therefore difficult to study the specific effects of individual growth factors on endothelial tubulogenesis in this material. Proteomics analysis has revealed the presence of transcription factors such as Kruppel-like factors 6 and 15, and other proteins that may have a potential impact on cell growth behavior in MatrigelTM (Hughes et al. 2010). Thus, MatrigelTM is a complex xenograft matrix environment that brings several non-physiological components to the study of human vasculogenesis. 3D gel-like matrices, composed of one or two purified ECM proteins such as fibrin and/or collagen I or IV, have also been used for the study of human endothelial tubulogenesis, thereby presenting a greatly simplified microenvironment as compared with natural, tissue-specific matrix (Chalupowicz et al. 1995; Hotary et al. 2003; Sieminski et al. 2004; Zhou et al. 2008).
Another approach for the study of vasculogenesis is to use 3D scaffolds produced by fibroblasts as platforms for endothelial tube formation. In vitro studies have detailed the assembly of matrix constituents by fibroblasts including fibronectin (FN), collagens (Col), and tenascin-C (TNC), and the importance of FN fibrillogenesis for matrix assembly and stability (Chung and Erickson 1997; Dzamba and Peters 1991; Sabatelli et al. 2001; Sottile and Hocking 2002). In our previous work, we detailed a technique for preparing natural, multi-component, decellularized matrix using human fetal lung fibroblasts (WI-38, ATCC) for the study of endothelial tubulogenesis (Soucy and Romer 2009; Soucy et al. 2011b). Multiple ECM protein and proteoglycan components, including FN, TNC, Col I, Col IV, Col VI, versican, and decorin, have been identified in this scaffold with unique organization patterns, and this microenvironment induced the complex patterns of cell adhesion, growth, and matrix remodeling that are required for endothelial tubulogenesis. This 3D ECM produced by human fetal fibroblasts may represent a more physiological model for the study of endothelial tubulogenesis as it is a product of human cells that incorporates multiple ECM components in characterized, native fibrillar forms and proportions, has an elastic modulus that is similar to the mechanics of decellularized lung matrix (Luque et al. 2013), and it contains neither pathological growth factors nor native factors in pathophysiological concentrations.
However, because the WI-38 fibroblast is a primary cell isolate, these cells become quiescent following multiple cumulative population doublings, most likely due to the shortening of telomeres (Pignolo et al. 1998). Transfection of WI-38 fibroblasts with a human telomerase reverse transcriptase successfully lengthened the telomeres in these cells, and extended proliferation (Milyavsky et al. 2003). We have harnessed this advance to develop a platform to manipulate ECM proteins into 3D matrix microenvironments by establishing stable transfectants of WI-38-hTERT with RFP-tagged FN cDNA; this allows for the synthesis and assembly of matrix where the primary FN constituent is TagRFP-FN. This advance addresses another limitation in vasculogenesis investigations: the inability to determine the cell type of origin (fibroblast vs. ECs) of the matrix that is formed during tubulogenesis.
In the current work, we systematically investigated ECM components and their organization in matrix produced and assembled by wild-type WI-38, WI-38-hTERT, and WI-38-hTERT-TagRFP-FN. The matrix constituents and infrastructure were similar in all three matrices and included FN, TNC, Col I, Col IV, and Col VI. Moreover, HUVECs formed active 3D adhesions, containing pY397-FAK and pY410-pCas130, as well as tubes on all three matrices. Most of the FN fibrils in the matrix produced by WI-38-hTERT-TagRFP-FN had an emission signal in the red channel, and this facilitated differentiation between FN fibrils that were made by WI-38-hTERT-TagRFP-FN from that were made by HUVECs (RFP-negative). The hypothesis that HUVECs synthesize significant amounts of FN and Col IV (Howard et al. 1976; Xue et al. 2010) during tubulogenesis (Grant et al. 1991; Wang and Su 2011; Zhou et al. 2008) was tested. Interestingly, we observed new FN fibrillogenesis by HUVECs along the newly formed tubes, suggesting that, although the 3D matrix from WI-38 fibroblasts provide initial cues for tubulogenesis, this process may also require de novo synthesis of FN fibrils by the EC. Membrane type 1 metalloprotease (MT1-MMP) was recruited to regions of active tubulogenesis, and may participate in degradation of pre-existing matrix and replacement by EC matrix. Although our immunofluorescence and immunoblotting data suggest that WI-38 do not efficiently assemble Col IV into the ECM, we show that HUVECs apparently assemble Col IV fibrils at sites of new vessel formation. Our data indicate that WI 38-hTERT- and WI-38-hTERT-TagRFP-FN-derived matrices may provide a physiologically accurate model for the study of 3D cell-matrix interactions.
Materials & Methods
Cell Culture and Reagents
WI-38 fetal human lung fibroblasts (an untransformed primary cell line; ATCC, Manassas, VA) were cultured in Eagle’s Minimal Essential Medium (Sigma-Aldrich, St. Louis, MO), supplemented with 10% FBS (Atlanta Biologics, Lawrenceville, GA), antibiotic-antimycotic solution (Cellgro, Lawrence, KS), 1 mM sodium pyruvate (Sigma-Aldrich) and non-essential amino acids (Sigma-Aldrich). WI-38-hTERT(Milyavsky et al. 2003), and WI-38-hTERT permanent transfectants expressing a tagged RFP chimera of fibronectin (TagRFP-FN) were cultured in the same medium components as for WI-38 except for the addition of L-glutamate. HUVECs, from VEC Technologies (Rensselaer, NY), were cultured in MCDB-131 complete medium.
Antibodies and Reagents
Primary antibodies used for this study included mouse monoclonal anti-fibronectin (BD/Transduction Laboratories; San Jose, CA); rabbit polyclonal anti-fibronectin (ab299, Abcam; Cambridge, MA); goat polyclonal anti-fibronectin (Santa Cruz Biotechnology; Dallas, TX); mouse monoclonal anti-tenascin-C (BC-24, Santa Cruz Biotechnology), rabbit polyclonal anti-tenascin-C (Hx-B2873) (Savarese et al. 1996); rabbit polyclonal anti-α1 collagen I (LF-67) (Bernstein et al. 1996); rabbit polyclonal anti-collagen VI (ab6588, Abcam); mouse monoclonal anti-collagen IV (col-94, Novus Biologicals; Littleton, CO); rabbit polyclonal anti-collagen IV (Novus Biologicals); mouse monoclonal anti-phospho FAK at Tyr397 (BD/Transduction Laboratories); rabbit polyclonal anti-phospho p130Cas at Tyr410 (Cell Signaling Technology; Danvers, MA); mouse monoclonal anti-MT1-MMP (LEM-2/15) (Genis et al. 2007); rabbit polyclonal anti-VE-Cadherin (Cayman Chemical; Ann Arbor, MI), mouse monoclonal anti-PECAM (4G6) (Yan et al. 1995), and affinity-purified goat polyclonal antiserum against LAP TGF-β1 (af-246-NA; R&D Systems; Minneapolis, MN). The following affinity cross-adsorbed secondary antibodies were used: Cy3-conjugated donkey anti-mouse and donkey anti-rabbit IgG; FITC-conjugated donkey anti-rabbit IgG; and Dylight 406 conjugated donkey anti-goat IgG (Jackson ImmunoResearch; West Grove, PA). Alexa Fluor 647-conjugated donkey anti-mouse IgG, and Alexa Fluor 488-, 568-, and 647-labeled carboxylic acid, succinimidyl ester mixed isomers were purchased from Molecular Probes (Carlsbad, CA).
Natural Matrix Preparation and EC Plating
Human lung fibroblasts (WI-38, WI-38-hTERT, or WI-38-hTERT-TagRFP-FN) were seeded onto FN-coated (50 μg/ml FN in Tris-Cl buffer, Invitrogen; Carlsbad, CA), 12-mm coverslips (no. 0, Thermo Fisher Scientific; Pittsburgh, PA) at confluence and grown for 10–20 days before cells were extracted to obtain the 3D ECM, as described previously (Soucy and Romer 2009). Alternatively, WI-38-hTERT-TagRFP-FN cells were seeded onto FN-coated, 35-mm glass bottom dishes (Plastek Cultureware; Ashland, MA) for 10–20 days prior to fibroblast extraction, and EGFP-FAK-transfected HUVECs were plated in these matrices for live cell imaging of 3D adhesion formation. Cellular and nuclear materials were extracted from matrices by incubation with 0.05% Triton X-100 (Thermo Fisher Scientific) and 50 mM NH4OH (Sigma-Aldrich) until the cells were floating. Samples were then washed briefly with 50 mM NH4OH, followed by incubation with 20 U/mL DNase I (Roche; Penzberg, Germany) at 37ºC for 1 hr. Matrices that were free of cellular and nuclear debris were washed three times with PBS and immediately used for antibody labeling or HUVEC seeding. HUVECs between passages 4 and 7 were seeded at a density of 2×105 cells/cm2 in MCDB-131 medium on the fibroblast-derived ECM for 1–3 days before fixing for epifluorescence analysis. HUVECs grown on coverslips with adsorbed FN (50 μg/ml) or gelatin (0.1%) were used as controls.
HUVEC Transfection
HUVECs from passages 4 to 7 were trypsinized and washed with PBS. Approximately 4×105 cells were resuspended in 20 μl of Nucleofector transfection solution and mixed with either 1 μg of EGFP-FAK DNA, 500 nM of siRNA for human MT1-MMP, or AllStars negative control siRNA (Qiagen; Valencia, CA). Transfection was performed with the Nucleofector 96-well shuttle system (Amaxa Biosystems; Gaithersburg, MD) using the P5 primary cell kit and program 96-CA-167. After nucleofection, cells were immediately transferred to pre-warmed medium and grown for 24 hr before use. Experiments were performed in duplicate wells. Trypsinized cells transfected with MT1-MMP or control siRNA were also sparsely seeded on FN-coated coverslips for anti-MT1-MMP immunofluorescence staining.
HUVEC Tube Formation
Tube formation was quantified 24 hr after plating cells on FN-derived matrix. Coverslips coated with 50 μg/ml FN at 37ºC for 1 hr were used as controls. Tubes were defined as tubular structures with a minimal width of 5–20 µm, and a length that was more than double the width. Tubes were counted per low-power field in each of four quadrants and the center of each coverslip, and these counts were averaged.
Cloning WI-38-hTERT-TagRFP-FN
TagRFP-FN was generated by modifying the FN-YPet/Neo construct that has been previously described (Ohashi et al. 2002). This construct is a modified form of the pAIPFN vector that has been used previously (Akamatsu et al. 1996; Ohashi et al. 1999). However, in this construct, the monomeric YPet fluorescent protein was inserted between FN Type III domains 6 and 7 via the insertion of a NotI restriction site, and a neomycin cassette was inserted to allow for selection of stable transfectants. To generate TagRFP-FN, TagRFP cDNA (Evrogen; Moscow, Russia via Wako; Richmond, VA) was amplified with the following primers: F:ggccaa
Purification of DOC-soluble and DOC-insoluble ECM Proteins
Due to the insolubility of the stable fibrillar matrix proteins in 2% deoxycholic acid (DOC) detergent (McKeown-Longo and Mosher 1983), it is possible to separate the cellular ECM proteins from those that have been incorporated into the matrix. Protein lysates from fibroblast cultures were obtained by scraping the contents in DOC lysis buffer (2% DOC, 20 mM Tris-Cl, pH 8.8, 2mM PMSF, 2 mM EDTA, 2 mM idoacetic acid, and 2 mM N-ethylmaleimide). Lysates were centrifuged at 15,000 × g at 4ºC for 15 min. Supernatants were taken as the DOC-soluble fractions. SDS-solubilization buffer (1% SDS, 20 mM Tris-Cl, pH 8.8, 2 mM PMSF, 2 mM EDTA, 2 mM idoacetic acid, and 2 mM N-ethylmaleimide) was then added to the pellets from the DOC buffer extractions to obtain the DOC-insoluble fractions. Total protein concentrations were measured in DOC-soluble fractions using the BCA protein assay kit (Pierce; Rockford, IL).
Immunoblotting
Volumes normalized for protein content of DOC-soluble or DOC-insoluble samples were boiled in 2× Laemmli sample buffer (0.1 M Tris HCl, pH 6.8, 0.2 M DTT, 25% glycerol, 1 mg/mL bromophenol blue, 4% SDS) for 5 min before being subjected to SDS-PAGE using 4–15% gradient gels for separation and analysis (Laemmli et al. 1970). Proteins were transferred to nitrocellulose membranes for western blotting. Membranes were blocked for 1 hr with 1% BSA (Sigma-Aldrich) in TBST (50 mM Tris-HCl, pH 7.6, 150 mM NaCl, 0.05% Tween-20), followed by incubation with primary antibody at 4ºC for 18 hr. Membranes were then washed three times with TBST before incubation with the appropriate horseradish peroxidase-conjugated anti-mouse, anti-rabbit or anti-goat IgG secondary antibody at room temperature for 1 hr. Proteins were visualized using enhanced chemiluminescence (Thermo Fisher Scientific) and exposure to X-ray film (Denville Scientific Inc.; South Plainfield, NJ) (Chang et al. 2007). Autoradiographs were scanned using an Epson Perfection scanner (Long Beach, CA).
Immunofluorescence Labeling and Epifluorescence Microscopy
Samples were permeabilized for 2 min with 0.5% Triton X-100 in 3% paraformaldehyde (Sigma-Aldrich) followed by fixation with 3% paraformaldehyde for additional 20 min. Antibodies were diluted in 0.1% BSA and were incubated for 30 min at 37°C. Imaging was accomplished with an epifluorescence Nikon TE-200 microscope (Melville, NY) and a Coolsnap HQ CCD camera (Roper, Duluth, GA) or a Rolera EMCCD camera (QImaging, Surrey, British Columbia) with Openlab or Volocity software (PerkinElmer/Improvision, Waltham, MA). Optical separation of emission signals was optimized by fluorophore choice.
Image analysis
Image z-stacks were deconvolved and then reconstructed in 3D using Huygens Essential software (Scientific Volume Imaging; Hilversum, The Netherlands) using the conditions as previously described (Soucy and Romer 2009). Volumetric colocalization of FN with TNC and Col VI was quantified using Volocity (PerkinElmer; Waltham, MA).
Statistics
Statistical differences in the colocalization of FN with TNC and Col VI were determined using an unpaired Student’s t-test in GraphPad Prism version 5.0 (GraphPad Software; San Diego, CA). All reported data represent the mean values ± SEM. Significance was considered to be present at p<0.05 for all data.
Results
Matrix Assembly in WI-38 WT and WI-38-hTERT Human Fetal Lung Fibroblasts
Cell morphology and proliferation rates of WI-38- hTERT (P38, P45, and P51), and WI-38 wild type (P4) human fetal lung fibroblasts were analyzed. All cells exhibited a polygonal cell shape with prominent focal adhesions and actin stress fibers before reaching confluence, and had an elongated, polarized, slender cell shape with microtubules and stress fibers parallel to the long axis after reaching confluence (Supplemental Fig. S1). Doubling times were measured during the exponential growth phase and were 2.03 days for WI-38-WT, 2.35 days for WI-38-hTERT P38, 2.02 days for P45 and 2.67 days for P51. All cells reached a plateau of 1.71–1.91×106 cells/cm2 at day 7 after plating. Synthesis and assembly of ECM proteins was studied using the methods of McKeown-Longo and Mosher (1983) established to distinguish short, fine fibrils of cellular FN that have just begun to associate with integrins from more mature, assembled fibrils based upon solubility in 2% DOC (Wierzbicka-Patynowski et al. 2004; Van Duyn Graham et al. 2010). DOC-soluble and DOC-insoluble fractions of FN, TNC, and Col I, IV, and VI from both WI-38-WT and WI-38-hTERT were analyzed at 2, 4, or 6 days after plating. These data demonstrated very similar matrix assembly dynamics in the two cell types (Fig. 1 and Supplemental Fig. S1D). Incremental quantities of TNC, Col VI and FN were documented in both the DOC-soluble and DOC-insoluble fractions from days 2–6, and each of these proteins were more abundant in the DOC-insoluble fractions. These data substantiate the similar synthesis and transport of proteins for ECM assembly in the WT and hTERT-transformed WI-38 fibroblasts.
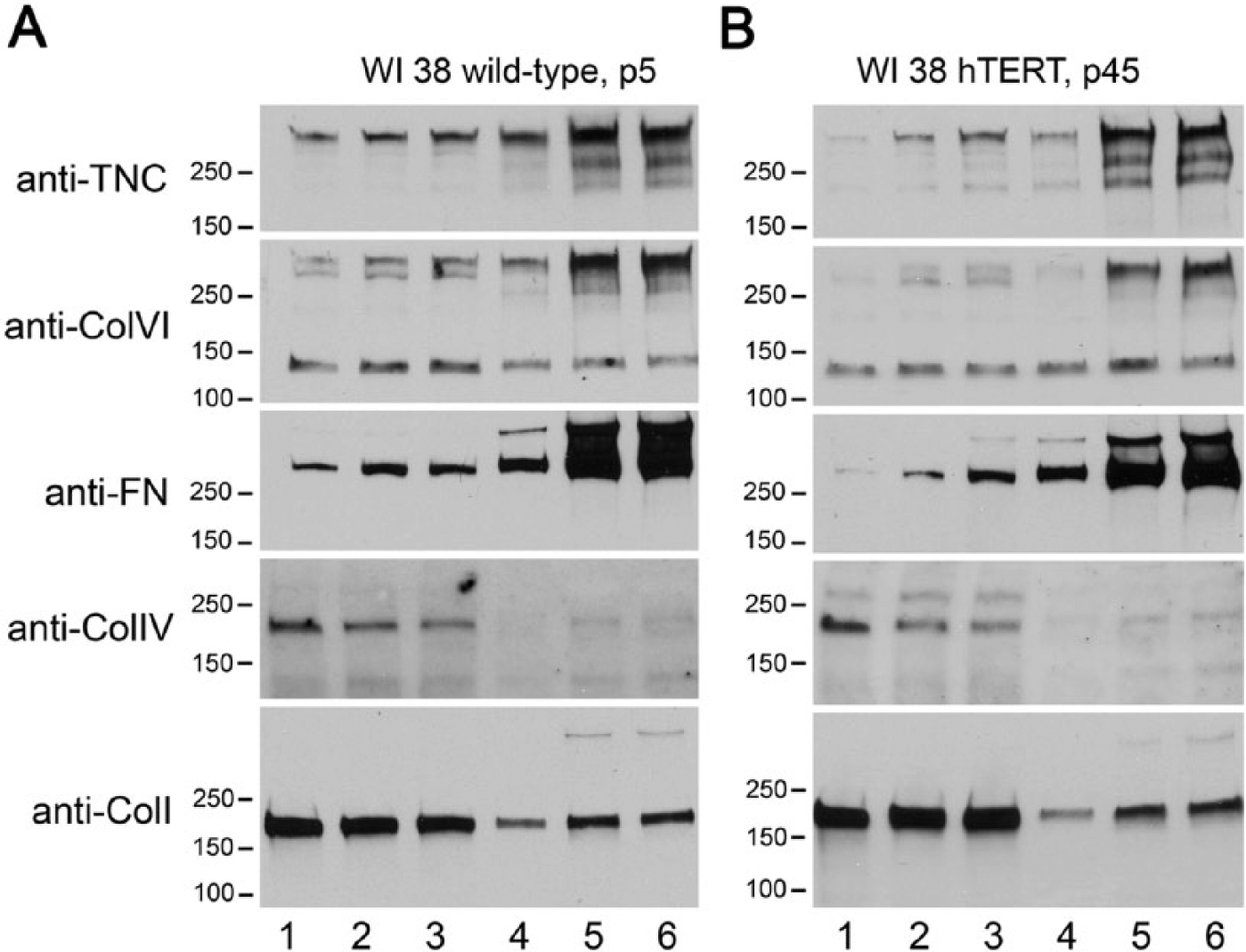
Protein constituents of matrices produced by WI-38-WT and WI-38-hTERT cells. ECM protein components including tenascin-C (TNC), type VI collagen (Col VI), fibronectin (FN), Col IV and Col I collagen were quantified by immunoblotting in the DOC-soluble fractions (lanes 1-3) and DOC-insoluble fractions (lanes 4-6) of WI-38-WT cells at passage 5 (panel A), and WI-38-hTERT cells at passage 45 (panel B). Samples were collected at 2 (lanes 1 and 4), 4 (lanes 2 and 5), or 6 days (lanes 3 and 6) after plating.
Matrix Infrastructure in WI-38-hTERT-derived ECM
The contributions of individual proteins to the 3D matrices produced by WT and WI-38-hTERT cells were analyzed by first labeling all of the amine groups in extracted matrices with Alexa Fluor 488 carboxylic acid succinimidyl ester, and then labeling matrices with antibodies to FN, TNC, Col IV, Col VI, and Col I (followed by species-specific, Cy5-conjugated secondary antibodies). Data indicate substantial and similar contributions of FN, TNC, Col I and Col VI to the 3D matrices produced by both cell types, and a sparse deposition of Col IV (Fig. 2A and 2B). These findings were concordant with both the immunoblotting data above that show an abundance of all of the studied ECM proteins except for Col IV in the DOC-insoluble protein fractions, and with previous observations that FN was patterned into thin, fibrillar structures that were distinct from the other proteins in WI-38-derived matrices (Soucy and Romer 2009).
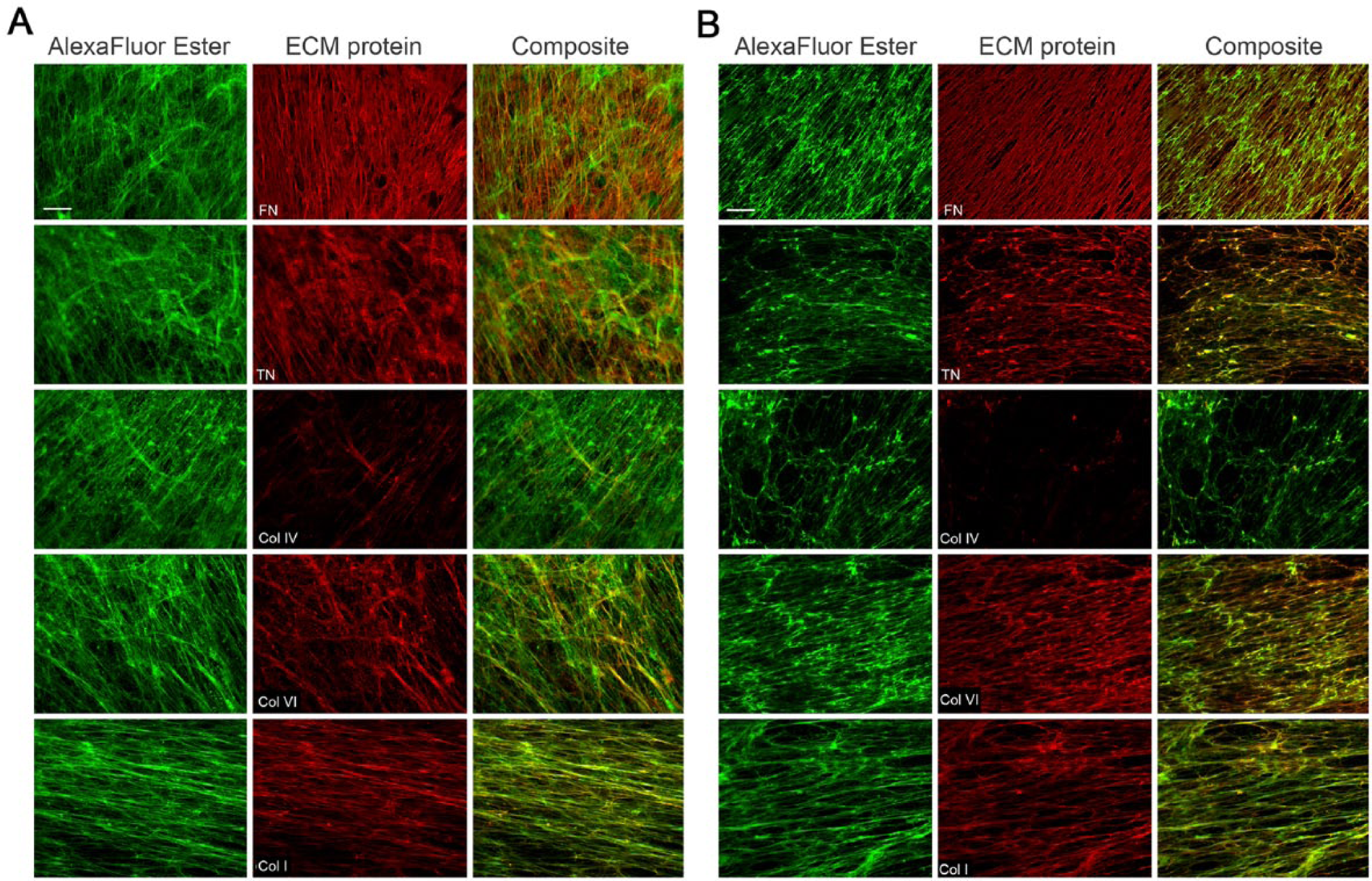
Patterns of ECM fibril organization in matrices produced by WI-38-WT and WI-38-hTERT cells. WI-38-WT (A) or -hTERT (B) fibroblasts were plated on fibronectin (FN)-coated coverslips and grown for 7 days. Cells were extracted, and 3D matrices were labeled with Alexa Fluor 488 carboxylic acid, succinimidyl ester (“Alexa Fluor Ester”, green), and then with antibodies against ECM proteins including FN, tenascin-C (TNC) and collagens (Col) I, -IV, and -VI, all followed by Cy5-conjugated secondary antibodies (all in red). Composite images demonstrate co-localization of individual ECM proteins with the total ECM protein. Scale bar, 20 μm.
Protein Organization in WI-38-hTERT-TagRFP-FN Matrix
To define the efficiency of TagRFP-FN incorporation in WI-38 matrix patterned by permanent TagRFP-FN transfectants, decellularized matrices labeled with antibodies against TNC, Col I, Col IV, or Col VI, or FN were analyzed by epifluorescence together with the TagRFP-FN (Fig. 3). TagRFP-FN was incorporated into the fibrillar structure of the 3D matrices. TNC, Col I, and Col VI all demonstrated some colocalization with FN, but Col IV did not. These colocalization relationships were similar to those observed in WT- and hTERT- fibroblasts (also see Fig. 4B). To determine how much of the FN fibrils in the matrix produced by WI-38-hTERT-TagRFP-FN cells showed the RFP signal, we labeled total FN with rabbit anti-FN antibody followed by Cy5-conjugated secondary antibody. Our result showed that approximately 98.3 ± 1.9% of the TagRFP-FN was labeled with anti-FN antibody whereas approximately 87 ± 7% of the total FN labeled with anti-FN also showed an RFP signal (Fig. 3B). This suggested that a high percentage of FN in the total matrix content had a TagRFP-FN signal.
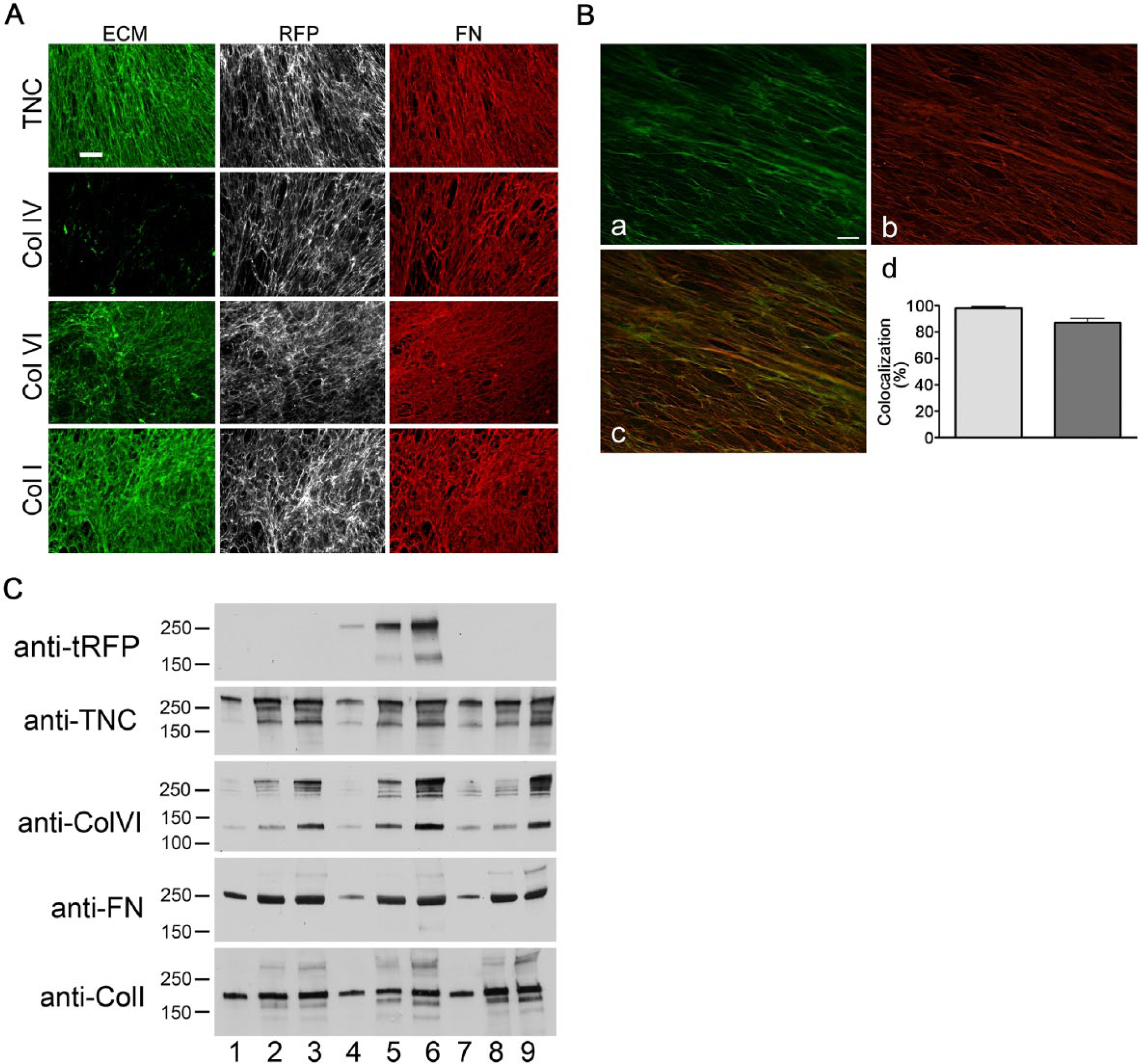
Patterns of fibril organization of matrices produced by WI-38-hTERT-TagRFP-FN fibroblasts. (A) WI-38-hTERT-TagRFP-FN cells produced matrix in 7 days, incorporating stably expressed TagRFP-chimeric fibronectin (FN) into the fibrillar matrix infrastructure (grey scale images). Matrices were labeled with antibodies against FN (red) and FITC-conjugated secondary antibody, as well as antibodies against tenascin-C (TNC), or collagen (Col) I, IV, or VI (each in green) followed by Cy5-conjugated secondary antibody. Scale bar, 20 μm. (B) Three-dimensional colocalization analysis is shown for deconvolved image z-stacks of TagRFP-FN (a, shown in green, emission maximum at 584 nm), and total FN labeled with antibody and Cy5-conjugated secondary antibody (b, shown in red, emission maximum at 670 nm) in the same field. Images depict one plane of a deconvolved z-stack. Scale bar, 20 μm. Colocalization was analyzed with Volocity software (d). Approximately 98.28% ± 1.952 (SD) of TagRFP-FN colocalized with total FN, whereas 87.14% ± 6.979 of the total FN colocalized with TagRFP-FN. (C) ECM protein components including TNC, FN, Col I and VI, in the DOC-insoluble fractions from WI-38 (lanes 1–3), WI-38-hTERT-TagRFP-FN (lanes 4–6), and WI-38-hTERT (lanes 7–9) were obtained 1 day (lanes 1, 4, and 7), 4 days (lanes 2, 5, and 8) and 8 days (lanes 3, 6, and 9) after cell plating and evaluated by immunoblotting. Antibody against TagRFP was used to show the TagRFP-FN produced by WI-38-hTERT-TagRFP-FN.
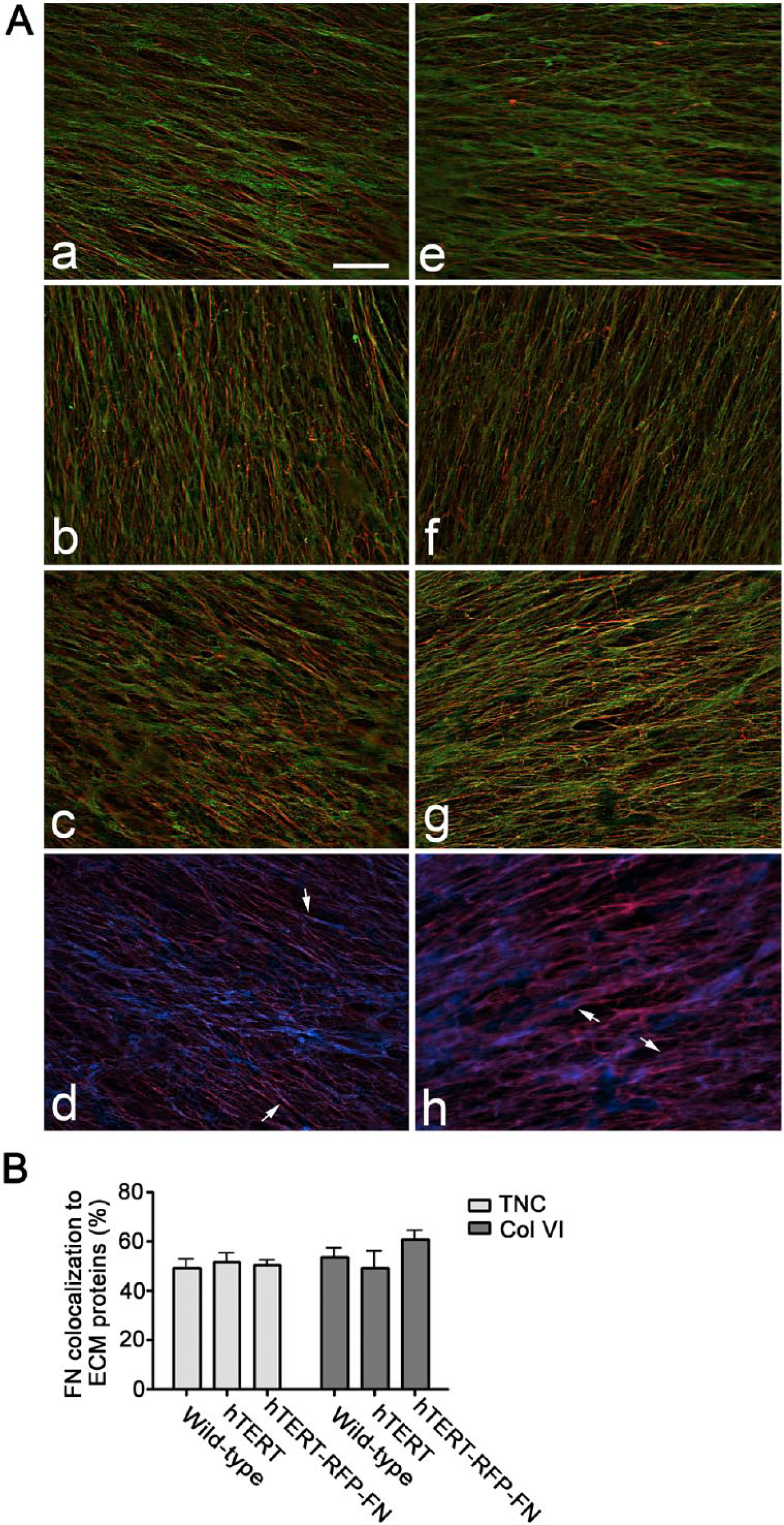
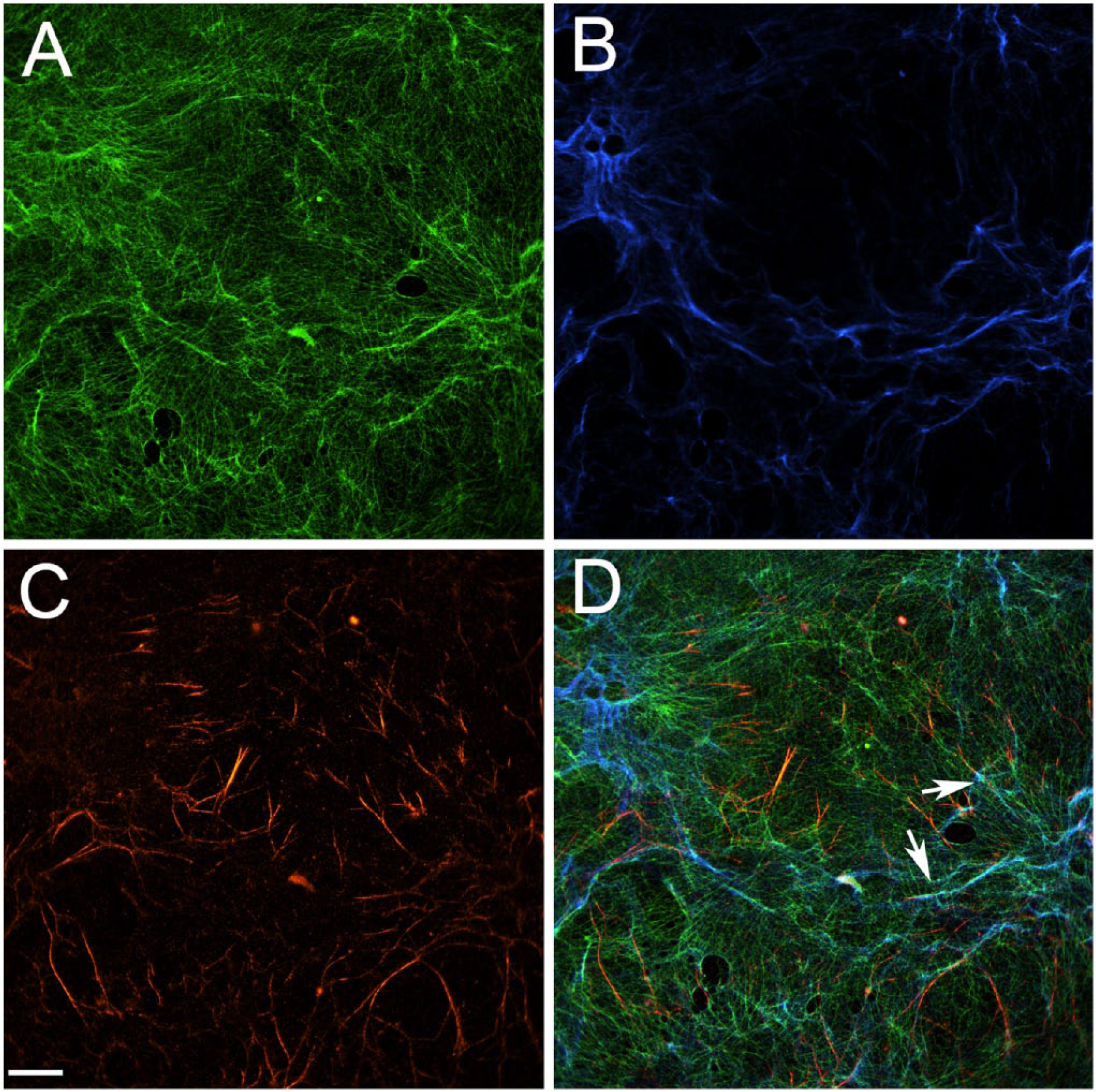
Distribution and colocalization of TNC or Collagen VI with FN in three fibroblast-derived matrices. (A) Composite immunofluorescence images of matrices produced by WI-38-WT (a and e), WI-38-hTERT (b and f) and WI-38-hTERT-TagRFP-FN (c, d, g and h) are shown. Fibronectin is shown in red (a, b, c, e, f, g); TNC (a, b, c) and collagen VI (e, f, g) are shown in green. TagRFP-FN from WI-38-hTERT-TagRFP-FN is shown in blue (d and h), and TNC and collagen VI each appear red in panels d and h, respectively. Colocalization of TNC or collagen VI with FN appears yellow (a, b, c, e, f, g), and colocalization of TagRFP-FN with TNC or collagen VI appears fuschia (d and h). Images depict one plane of a deconvolved z-stack. Representative discrete colocalization foci are pointed out by small arrows. Scale bar = 20 μm. Cy5-conjugated goat anti-rabbit IgG was used to label tenascin-C and Collagen VI while FITC-conjugated goat anti-mouse IgG was used to label fibronectin. Mean percentage colocalization of fibronectin to TNC or Collagen VI in matrices produced by WI-38-WT, WI-38-hTERT and WI-38-hTERT-TagRFP-FN are shown (B): the percentage of fibronectin that colocalized to tenascin-C was 49.33, 51.83, and 50.63 % in the 3 matrices, respectively. The percentage of fibronectin that colocalized to collagen VI was 43.47, 59.00, and 63.40% in the three matrices.
DOC-soluble and DOC-insoluble fractions of ECM proteins from WI-38-hTERT-TagRFP-FN were compared with those from WT WI-38 (Supplemental Fig. S2). This analysis showed that all four ECM proteins tested were efficiently assembled into fibrils in both WI-38 and WI-38-hTERT-TagRFP-FN matrices, in similar proportions. Matrix produced by WI-38-hTERT-TagRFP-FN (but not by wild type WI-38) was labeled by anti-TagRFP antibody (Fig. 3C).
Characterization of Matrices Produced by WI-38-WT, -hTERT, and -hTERT-TagRFP-FN Cells
FN fibrils are believed to be an important signal-initiating element of fibroblast-derived matrix, and have been shown to have different topography and localization characteristics compared with other ECM proteins in WI-38 matrix. We studied the colocalization of FN with TNC and Col VI in matrices produced by WI-38, WI-38-hTERT, and WI-38-hTERT-TagRFP-FN fibroblasts, and found very similar colocalization relationships in all three matrices (Fig. 4).
3D Adhesions Containing pY397-FAK and pY410-p130Cas Are Formed in All of the Studied Matrices
FAK and pCas130 are adhesion components that contribute to high traction forces and cell motility in 3D matrices (Fraley et al. 2010), and have been associated with EC tubulogenesis. In the present study, HUVECs formed 3D adhesions containing activated, tyrosine-phosphorylated pY397-FAK and pY410-p130Cas on the matrices produced by hTERT-expressing WI-38 (Fig. 5). HUVECs were elongated and showed smaller adhesions on 3D matrices produced by all three WI-38 cell populations. EGFP-FAK-transfected HUVECs were seeded onto the matrix produced by WI-38-hTERT-TagRFP-FN and time-lapse images (Supplemental Fig. S2) demonstrate the utility of the chimeric TagRFP-FN-containing matrix with real-time imaging of cell adhesion.
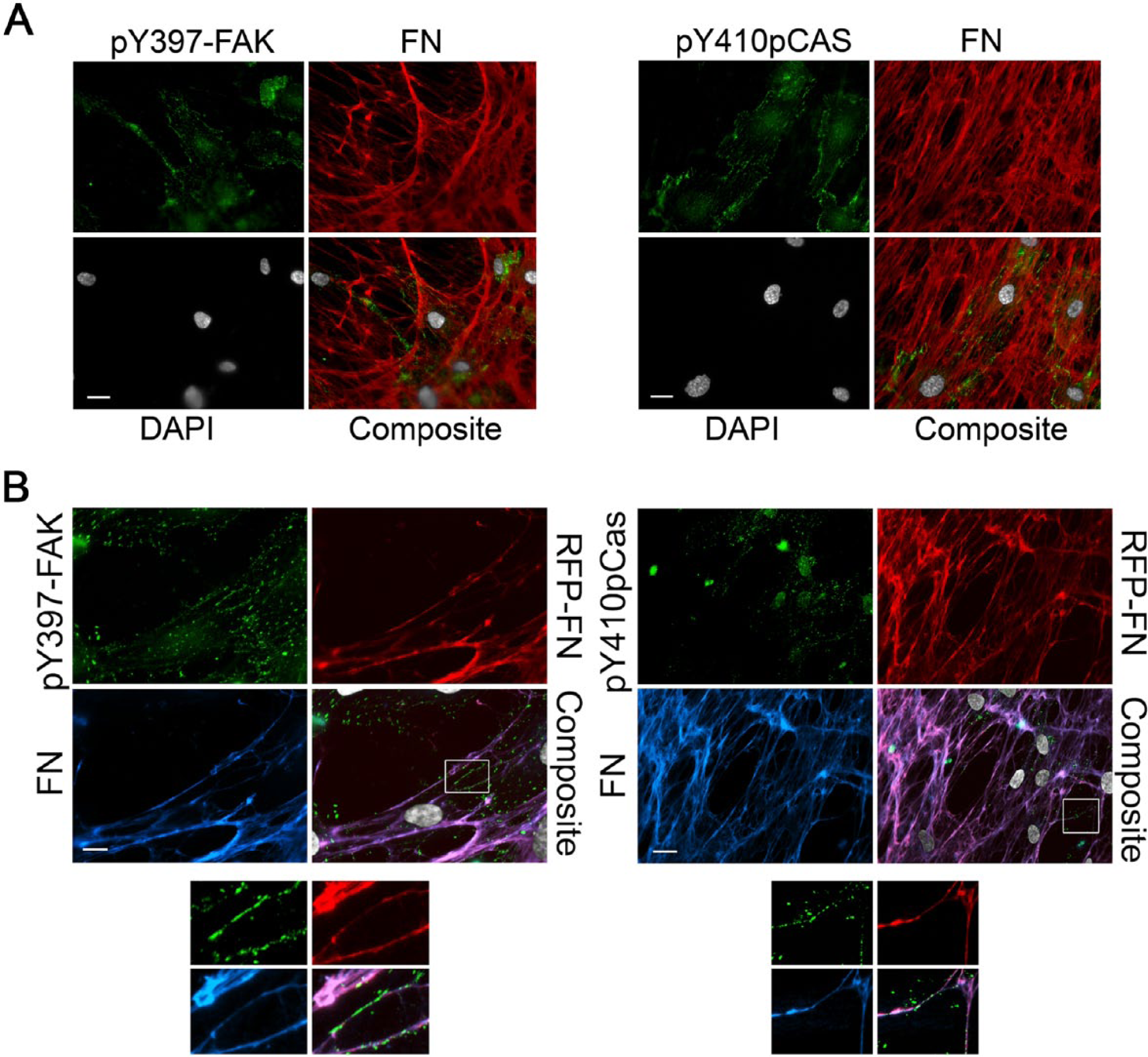
3D endothelial cell (EC) matrix adhesion formation on matrices produced by WI-38-hTERT and WI-38-hTERT-TagRFP-FN cells. HUVECs were seeded onto matrices produced by WI-38-hTERT (A) or WI-38-hTERT-TagRFP-FN (B) cells for 24 hr. Cells were fixed and labeled for fibronectin (FN; red in (A) and blue in (B)), DAPI (grey), and either pY397-FAK (green in panel on left) or pY410-p130Cas (green in panels on right). TagRFP-FN is shown as red in (B). Scale bar, 20 μm. Insets from regions marked by boxes in 5B were magnified 2.7×, and each appears below the respective original image.
HUVEC Tubulogenesis in WI-38-hTERT and WI-38-hTERT-TagRFP-FN Matrices
We next tested the functionality of the decellularized matrix produced by WI-38-hTERT and WI-38-hTERT-TagRFP-FN cells, as compared with the parental WT WI-38 cells, via the induction of HUVEC tubulogenesis. Prominent networks of tubes were formed by HUVECs in both WI-38-hTERT and WI-38-hTERT-TagRFP-FN matrices, similar to that produced by WT WI-38 cells (Fig. 6), suggesting that the requisite morphogenic cues were present in microenvironments produced by all three cell types.
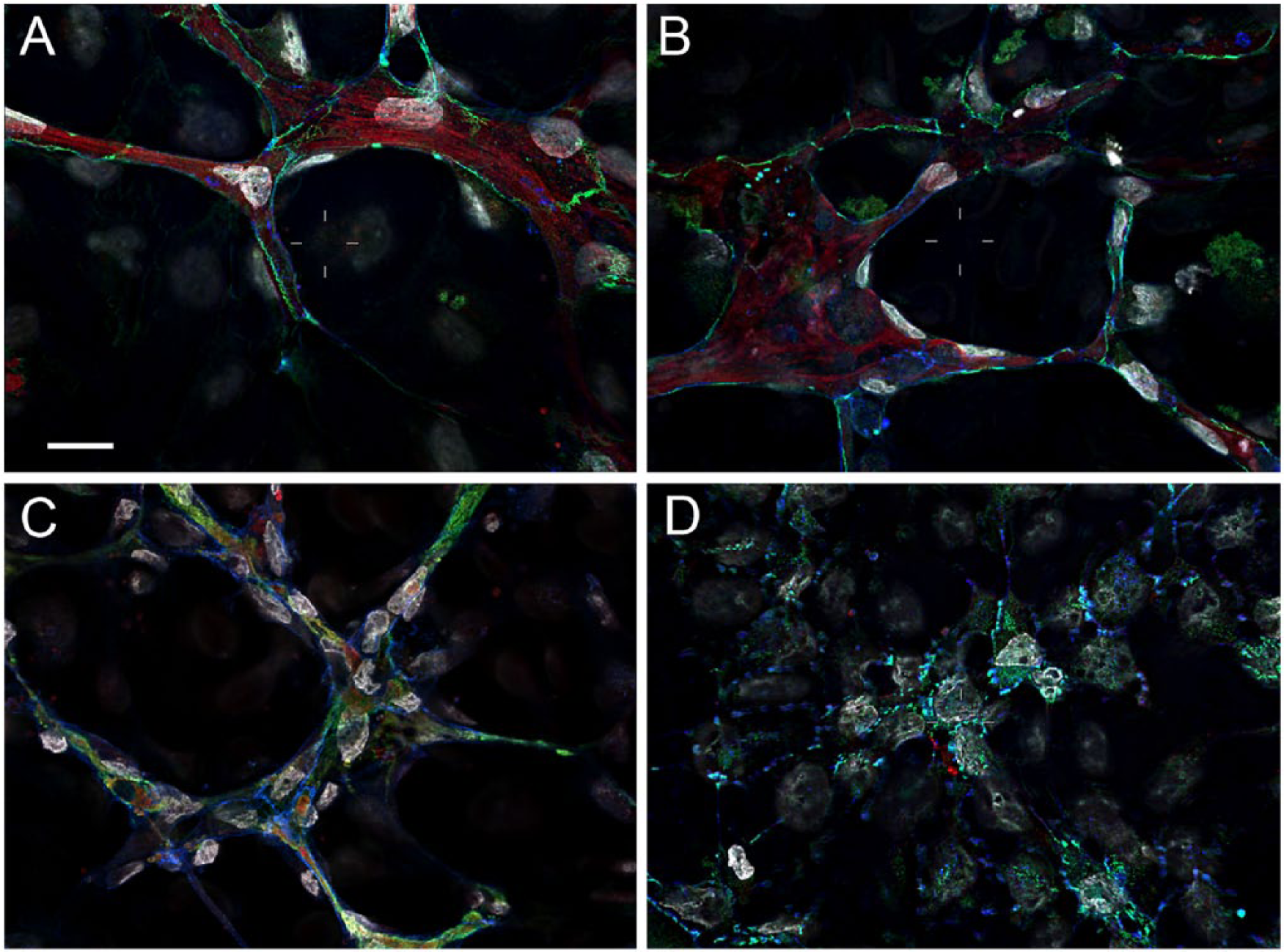
Endothelial tubulogenesis in matrices produced by WI-38-hTERT and WI-38-hTERT-TagRFP-FN cells as compared with parental cells. Decellularized matrices produced by the parental WI-38 cells (A) and by both the WI-38-hTERT (B) and WI-38-hTERT-TagRFP-FN (C) cell lines induced tubulogenesis in HUVECs. No tubulogenesis was observed on FN-coated coverslips (D). Antibody labeling of VE-Cadherin (vascular endothelial-cadherin, green) and FN (red) is shown. DAPI is in grey, and phalloidin is in blue. Scale bar, 20 μm.
MT1-MMP and Tubulogenesis in WI-38 Matrix
We next examined the role of MT1-MMP in EC tubulogenesis on fibroblast-derived natural 3D matrices. We demonstrated that both the general MMP inhibitor GM6001 (but not the matched biochemical control), and siRNA specific for MT1-MMP (but not the negative control siRNA) significantly reduced tube formation by 24 hr in these matrices. Transfected HUVECs were also sparsely seeded on FN-coated coverslips and assayed for MT1-MMP expression by immunofluorescence (Fig. 7A).
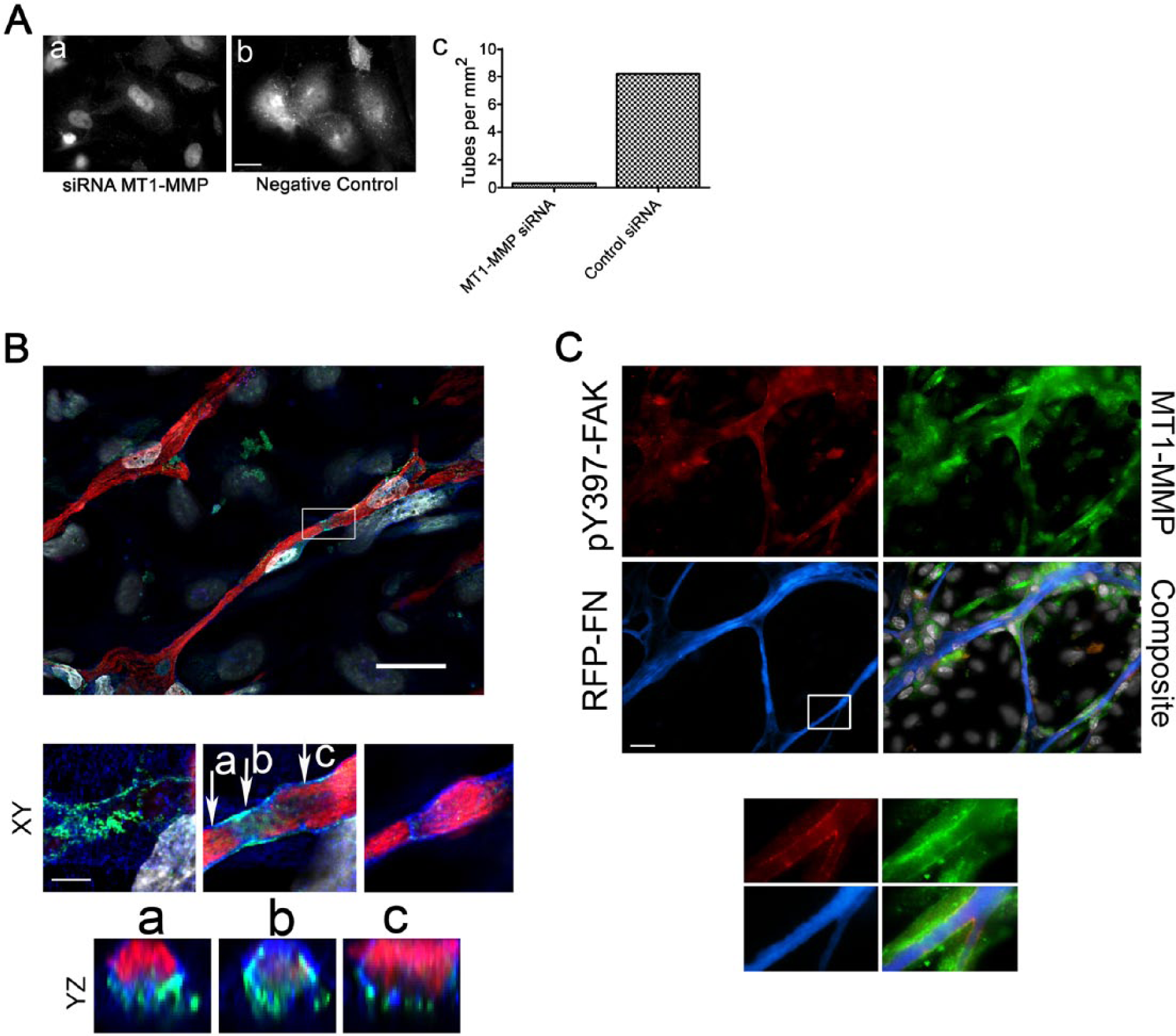
Clarification of the role of membrane type 1 metalloprotease (MT1-MMP) in WI-38 matrix remodeling during HUVEC tube formation using TagRFP-FN. (A) siRNA specific for human MT1-MMP abrogated MT1-MMP detection by immunofluorescence in HUVECs plated on WI-38-derived matrices (a), whereas negative control siRNA did not (b). MT1-MMP siRNA also reduced tubulogenesis at 24 hr (c; Scale bar, 20 μm). (B) 3D reconstructions of a HUVEC tube formed on WI-38 matrix depict the close spatial relationships between fibronectin (FN; red), platelet EC adhesion molecule (PECAM-1; green), and matrix metalloproteinase 14 (MMP14; blue) in the XY and YZ (magnified 2×) planes of the tube (DAPI is in grey). Scale bar, 5 μm. (C) HUVEC tubulogenesis in WI-38-hTERT-TagRFP-FN matrix, showing pY397-FAK in HUVEC focal adhesions (red), MT1-MMP (green), and TagRFP-FN (blue). Nuclei are grey (DAPI). Scale bar, 18 μm. Insets from the region marked with a box are shown for all channels and for the merged composite (with 3.4× magnification). MT1-MMP is concentrated at EC adhesion sites that contain activated FAK and are apparently degrading the TagRFP-FN.
The subcellular localization of MT1-MMP was also investigated during HUVEC tubulogenesis (Fig. 7B). The deconvolved images show that FN fibers were enveloped by adhesions containing MT1-MMP that may mediate matrix degradation and remodeling—an important component of the tubulogenesis process. Because FAK has been previously implicated in MT1-MMP focusing events at cell adhesion sites in the work of others (Kwiatkowska et al. 2011; Takino et al. 2010; Wu et al. 2005), we examined the spatial relationships between FAK, MT1-MMP, and FN using WI-38-hTERT-TagRFP-FN matrix. Interestingly, we observed tight colocalization of pY397-FAK and MT1-MMP at the TagRFP-FN fibers that were being remodeled by ECs (Fig. 7C). These data are in agreement with current concepts of the role of MT1-MMP in EC-mediated matrix remodeling, and demonstrate the utility of matrices assembled by WI-38-hTERT-TagRFP-FN cells in the study of matrix dynamics.
ECs Synthesize and Assemble FN and Col IV during Tubulogenesis in Natural Matrix
We have previously shown that fetal lung fibroblasts assemble a multicomponent matrix that is instructive for microvascular morphogenesis (Soucy and Romer 2009; Soucy et al. 2011a). These matrices have abundant FN fibrils that are highly probable adhesion sites for the ECs that trigger tyrosine phosphorylation of FAK and p130Cas. These data are in concert with the findings of Zhou et al. (2008), where a Col I and fibrin gel model was used to implicate FN fibrillogenesis as an important event in EC tube formation (Zhou et al. 2008). We therefore used fibroblast-derived matrices containing TagRFP-FN to identify whether FN fibrils that are contiguous to new EC tubes are synthesized by fibroblasts and rearranged by ECs, or actually synthesized by HUVECs themselves. Our data indicate that HUVECs participate directly in new FN fibrillogenesis during the formation of EC tubes in 3D matrix (Fig. 8A) and in the formation of Col IV fibrils that are distinct from the remodeled 3D TagRFP-containing matrix (Fig. 8B, C).
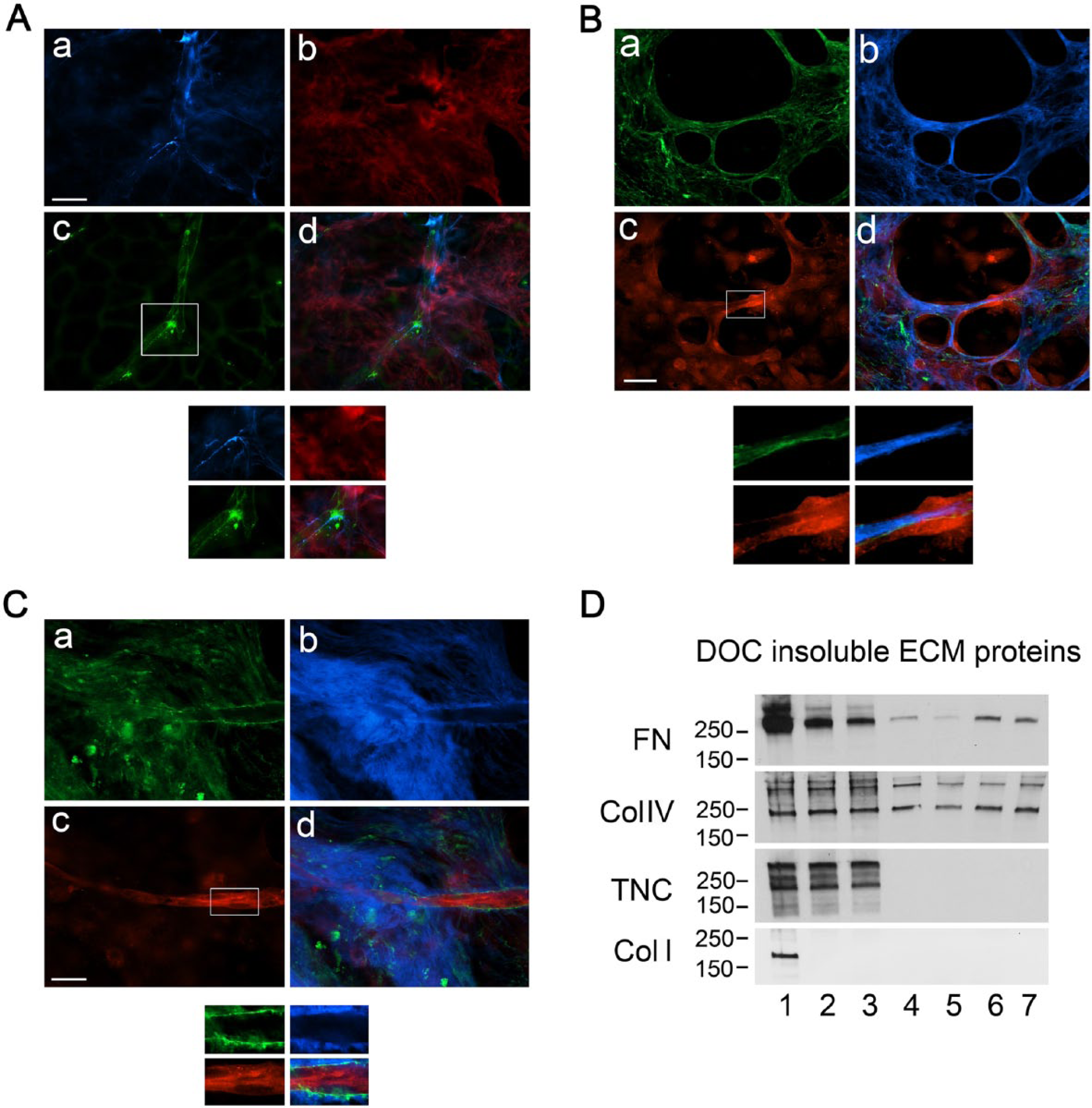
FN and Col IV fibril assembly by HUVEC during tubulogenesis in 3D matrices. HUVEC cells were plated on WI-38-hTERT-TagRFP-FN matrices (7 days old (panel B) or 14 days old (panels A and C) for tubulogenesis assays. Following fixation, samples were labeled as follows. For panel A, antibodies to FN (blue) and VE-cadherin (green) were used for epifluorescence analysis, and chimeric, fibroblast-derived matrix (14 days) labeled with TagRFP-FN is shown in red. For panels B and C, antibody labeling for collagen IV is shown in green, and for MT1-MMP in red, while the TagRFP-FN is shown in blue. FN and collagen IV fibrillogenesis appear to have been initiated by the nascent HUVEC tubes themselves (panels A and C, respectively. Scale bars = 20 μm. Insets marked by the boxes were magnified 1.8-fold and shown for all channels below the respective figures. (D) Immunoblots of DOC-insoluble ECM proteins are shown from a control WI-38 population prior to cell extraction (lane 1), and from HUVEC plated for 1 or 2 days on decellularized W-I38 matrix (lanes 2 and 3), or adsorbed FN (50 μg/ml; lanes 4 and 5), or gelatin (0.1%; lanes 6 and 7). HUVEC that were plated on FN or gelatin organized DOC-insoluble FN and collagen IV. DOC-insoluble FN in the WI-38-derived matrix was degraded by HUVEC during tube formation, but collagen IV was not.
To investigate ECM synthesis and fibrillogenesis by HUVECs themselves, cells were grown on 3D WI-38 matrices or on FN- or gelatin-coated cover slips. WI-38 fibroblasts and the matrix they produced served as controls. Immunoblotting analysis indicated that the ECs synthesized and assembled fibrils of both FN and Col IV on the 2D matrix-coated coverslips, but did not form fibrils of Col 1 or TNC (Fig. 8D). The DOC-insoluble extract of WI-38 with the matrix showed substantial amounts of all four proteins. Col I was rapidly degraded by the HUVECs. FN content in the DOC-insoluble extract showed daily decrements, suggesting that the rate of FN degradation by the ECs exceeded the rate of EC-specific FN fibrillogenesis. The total amounts of TNC and Col IV in the DOC-insoluble fraction of the fibroblast matrix were apparently not changed by the ECs. Figure 9 shows that HUVECs assemble a dense FN matrix over a 2-week period, and that type IV collagen and latent TGF-β are both incorporated in this endothelial-specific matrix microenvironment. These data indicate that HUVECs directly synthesize and assemble both FN and Col IV matrix fibrils.
Latent transforming growth factor (TGF)-β is present in matrix produced by HUVECs. Cells were grown on fibronectin (FN)-coated coverslips for 14 days prior to extraction. 3D matrices were then fixed and labeled with antibodies against FN (green), type IV collagen (Col IV; blue), and latent TGF-β (red). Arrows highlight regions of colocalization of FN and/or Col IV with latent TGFβ-1. Scale bar, 20 μm.
Discussion
Accumulating data suggest that the ECM provides a template for tissue morphogenesis (Lutolf and Hubbell 2005; Macchiarini et al. 2008; Ott et al. 2008). The identification of specific matrix cues for specialized cell responses has become a critical and exciting new frontier for developmental biology, regenerative medicine, and tissue engineering. The use of natural, cell-derived matrices for these studies has many advantages. Natural matrices are complex substrates with several attributes that have proven functionality. These include spatial orientation cues that guide the infiltration and growth of axons, fibrillar elements that serve as a template for cell migration and morphogenesis, and mechanical heterogeneities that serve as trophic stimuli for specific cell subpopulations and may direct the topography of tissue pathology in cancer (Kutys et al. 2013; Lopez et al. 2011; Nardi 1983). The current work was undertaken to improve the number and quality of matrix biology insights that may be obtained from studies of tissue morphogenesis and tumorigenesis in natural matrix models. The development of stable transfectants of tissue-specific fibroblasts enabled us to establish a platform for the study of matrix molecular biology and the roles of individual matrix constituents and their functional domains in the morphogenic matrix microenvironment.
The systematic analysis of ECM components and infrastructure, as well as EC adhesion signaling and morphogenesis, is reported here for matrix produced by both WI-38-hTERT and WI-38-hTERT-TagRFP-FN. These data demonstrate that matrices produced by the hTERT-transformed cells were quantitatively and functionally nearly identical to that produced by the parent fibroblast line. Milyavsky et al. (2003) reported that WI-38-hTERT displayed patterns of proliferation and growth that quantitatively recapitulated those of WT for up to 150 population doublings. The work presented here corroborates those observations, as we used WI-38-hTERT clones with 33-50 cumulative population doublings. Others report favorable results in studies that have harnessed hTERT-transformed fibroblasts and human mesenchymal stem cells for tissue engineering applications ranging from vocal cord lamina propria to vascularized tissues (Burns et al. 2011; Chen and Thibeault 2009).
Using a 3D gel model, Zhou et al. (2008) reported that HUVEC FN fibrillogenesis was important for endothelial tube formation. Their model employed a mixture of fibrin and Col I to form a gel in which growth factors, including vascular endothelial growth factor (VEGF), human hepatocyte growth factor (HGF) and TGF-α and -β1, were used to induce HUVEC tubulogenesis (Hotary et al. 2003). They found that ECs initiated the process of new blood vessel formation by using soluble FN to assemble and deposit a pericellular network of fibrils to support cytoskeletal organization, actomyosin-dependent tension, and appropriate viscoelastic properties. When the assembly of FN fibrils by ECs was inhibited by antibodies or peptides, HUVEC tubulogenesis was significantly decreased. In other work, secretome analyses of cell populations associated with high vs. low levels of angiogenesis, the process of budding new blood vessels from preexisting ones, have also implicated FN production as a pro-angiogenic process (Newman et al. 2013). We hypothesized that WI-38 fibroblast-derived matrix also provided stimulatory cues for both FN fibril assembly and tubular morphogenesis by ECs. The WI-38-hTERT-TagRFP-FN clone that we isolated produced a 3D matrix that was similar to that produced by WT WI-38. In this matrix, approximately 90% of the FN fibrils were positive for RFP. This allowed us to distinguish fibroblast-derived FN fibrils from EC-derived FN, and to demonstrate the location and dynamics of EC-specific FN fibrillogenesis during new blood vessel formation. Potential applications of our reagents and insights to in vivo work include subcutaneous or intrapleural implantation of pre-assembled matrix modules into transgenic mice to define the contributions of specific integrins to angiogenesis or vasculogenesis at those sites.
Our data further indicate that HUVECs produced Col IV and latent TGF-β in regions of 3D matrices that are contiguous with de novo blood vessel formation, vasculogenesis, and that EC-derived matrix may replace fibroblast-derived matrix during this process. Indeed, the finding that siRNA-mediated knock down of MT1-MMP (matrix metalloproteinase 14) production by HUVECs drastically inhibited EC tubulogenesis indicates that remodeling and replacement of fibroblast matrix may be indispensable components of vasculogenesis, and this is consonant with work from Arroyo’s group on MT1-MMP in the context of angiogenesis (Genis et al. 2007). Genetic and proteomic analysis of EC-derived matrix proteins will be needed to discern the key attributes that contribute to endothelial morphogenesis. Our data indicating that cell type-specific matrix production is important in the context of this morphogenic program is supported by the findings of others that indicate that fibroblasts, smooth muscle cells and ECs uniquely contribute to matrix pathophysiology during aortic aneurysm formation and diabetic vasculopathy (Jones et al. 2009; Raman et al. 2011).
The WI-38-hTERT-TagRFP-FN matrix described in the current work may find use in a variety of developmental biology and tissue engineering applications. As a platform for matrix molecular biology, it may serve as a means for testing structure-function relationships in the domains of important matrix proteins. As a marker for cell population-specific contributions to matrix, this approach may be harnessed to precisely identify sources of developmental and pathological alterations in the matrix microenvironment during morphogenesis, fibrosis, and tumor invasion. Additionally, this reagent will facilitate the definition of detailed ECM-cell interactions and cell-mediated matrix turnover in real time (as illustrated by our study of EGFP-FAK-transfected HUVEC on WI-38-hTERT-TagRFP-FN matrix in Supplemental Fig. S2), and may be modified to define specific molecular interactions by FRET.
In summary, the work we presented here describes a biologically relevant 3D ECM model using the matrix produced by WI-38-hTERT-TagRFP-FN for the study of endothelial morphogenesis. This unique model allows us to dissect the contribution of the ECM fibrils made by WI-38 fibroblasts from that of EC-derived matrix. We observed degradation of fibroblast-derived WI-38 FN fibrils by MT1-MMP and the new assembly of both FN and collagen IV fibrils around the newly formed HUVEC tubes that was concomitant with this matrix remodeling, and the presence of latent TGF-β in matrix produced by ECs. This system may represent an effective new platform for the study of the interaction of EC matrix interactions during vasculogenesis or angiogenesis by high-resolution microscopy of living systems, and may contribute to the design of biomimetic and hybrid scaffolds for vascular regeneration.
Footnotes
Acknowledgements
The authors thank the following investigators for their kind gifts: Larry Fisher for the rabbit polyclonal anti-α1collagen I antibody, Harold Erickson for the rabbit polyclonal anti-tenascin-C antisera, and Alicia Arroyo for the monoclonal anti-MT1-MMP antibody.
The authors will endeavor to provide research materials that they have developed for this work for collaborative use by others in accordance with the policies of the NIH, NSF, AHA, DOD, and the Johns Hopkins University.
Supplementary Material
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants to L.R. from the