
Abstract
Mast cells are versatile effector cells of the immune system, contributing to both innate and adaptive immunity toward pathogens but also having profound detrimental activities in the context of inflammatory disease. A hallmark morphological feature of mast cells is their large content of cytoplasmic secretory granules, filled with numerous secretory compounds, including highly negatively charged heparin or chondroitin sulfate proteoglycans of serglycin type. These anionic proteoglycans provide the basis for the strong metachromatic staining properties of mast cells seen when applying various cationic dyes. Functionally, the mast cell proteoglycans have been shown to have an essential role in promoting the storage of other granule-contained compounds, including bioactive monoamines and different mast cell-specific proteases. Moreover, granule proteoglycans have been shown to regulate the enzymatic activities of mast cell proteases and to promote apoptosis. Here, the current knowledge of mast cell proteoglycans is reviewed.
Mast cells are currently emerging as major effector cells in numerous disorders. In most settings, mast cells have become infamous for their detrimental actions, as exemplified by diseases such as allergic asthma, contact dermatitis, arthritis, atherosclerosis, and cancer (Lee et al. 2002; Yu et al. 2006; Soucek et al. 2007; Sun et al. 2007; Dudeck et al. 2011). However, in other settings, mast cells are known to be beneficial, most notably in the context of host defense toward insults by bacteria, parasites, and various toxic substances (Echtenacher et al. 1996; Malaviya et al. 1996; Maurer et al. 2004; Dawicki and Marshall 2007; Akahoshi et al. 2011). Many of the actions of mast cells, both detrimental and beneficial, can be ascribed to those compounds that they secrete when activated. These include a large number of preformed substances present within the mast cell secretory granules, such as bioactive monoamines (histamine, serotonin, dopamine), certain preformed cytokines (e.g., tumor necrosis factor [TNF]), highly anionic serglycin proteoglycans (PGs) containing glycosaminoglycan (GAG) side chains of either heparin or chondroitin sulfate (CS) type, and a panel of mast cell-specific proteases, the latter encompassing chymases, tryptases, and carboxypeptidase A3 (CPA3) (Pejler et al. 2010; Lundequist and Pejler 2011). In addition to releasing preformed compounds from secretory granules, activated mast cells synthesize numerous other compounds de novo, including eicosanoids and a large number of cytokines, chemokines, and growth factors (Metcalfe et al. 1997; Kalesnikoff and Galli 2008).
A hallmark feature of mast cells from all species is their striking metachromatic staining with various cationic dyes such as toluidine blue. These characteristic staining properties have been used since the late 19th century to identify mast cells as such (Ehrlich 1878). The strong metachromasia is explained by binding of the respective dyes to the highly anionic PGs of serglycin type present within the mast cell secretory granules. The notion that mast cell granules contain PGs has been well established for many years, and much work has been conducted to define the nature of the granule PGs, as well as to address their functional properties. Here, the current status of knowledge of the mast cell PGs is reviewed.
Identification of Glycosaminoglycans as Components of Mast Cell Granules
In pioneering work, evidence was obtained suggesting that GAGs of the heparin type were present in mast cells (Holmgren and Wilander 1937; Jorpes et al. 1937). These findings were confirmed and extended by showing that heparin is a main component (~30% of the dry weight) of granules isolated from rat peritoneal mast cells (Lagunoff et al. 1964). In these early studies, the identification of heparin was based on its metachromatic properties (Hill 1957; Fillion et al. 1970; Slorach 1971), whereas later studies have used various other methods such as [35S]sulfate labeling, sensitivity to degradation by nitrous acid (pH 1.5) or heparinase, and content of hexuronic acid (glucuronic acid [GlcUA]/iduronic acid [IdoUA]) (Ögren and Lindahl 1971; Yurt, Leid, and Austen 1977; Metcalfe et al. 1979).
Most of the early work on mast cell heparin was conducted on cells of rat origin (e.g., isolated from peritoneum), and through studies of rat mast cells, it was shown that heparin was released after challenge with non-physiological stimulants such as oil (Hill 1957) and compound 48/80 (Fillion et al. 1970; Slorach 1971). Importantly, heparin was released concomitantly with histamine, indicating the presence of heparin within the secretory granules (Fillion et al. 1970; Slorach 1971). More firm proof for the location of heparin within granules came when Yurt et al. showed that immunological activation of mast cells through IgE receptor crosslinking caused release of 35S-labeled heparin into the extracellular space (Yurt, Leid, Spragg, et al. 1977).
The presence of heparin in human mast cells was first suggested through studies of lung tissue fragments (Metcalfe et al. 1979). It was demonstrated by biosynthetic 35S-labeling experiments that lung tissue contains GAGs of heparin type, and it was also demonstrated that heparin was produced by cell preparations partly enriched for mast cells (Metcalfe et al. 1979). Moreover, based on sensitivity to digestion by heparinase and chondroitinase ABC, respectively, it was suggested that human lung mast cells, in addition to heparin, also contain chondroitin sulfate (CS). In a subsequent study, it was shown that CS accounted for around one-third of the 35S-labeled GAGs produced by highly purified human lung mast cells, the remainder being identified as heparin (Thompson et al. 1988). Interestingly, it was shown that ~25% of the CS disaccharide units were of the “oversulfated” GlcUA-N-Acetylgalactosamine (GalNAc)-4,6-di-O-sulfate (CS-E) type, in addition to those of the more common monosulfated GlcUA-GalNAc-4-O-sulfate type (Thompson et al. 1988).
Studies of PGs in mouse mast cells were initially based on a murine mastocytoma. In this system, 35S-labeling experiments provided evidence for the synthesis of heparin (Ögren and Lindahl 1971). Heparin synthesis was also demonstrated in murine lymph node-derived mast cells maintained in co-culture with fibroblasts (Bland et al. 1982). In another study, Razin et al. (1982) showed that mast cells differentiated from bone marrow precursors (i.e., bone marrow-derived mast cells [BMMCs]) synthesized PGs that exclusively contained CS chains rather than heparin (Razin et al. 1982). Moreover, it was demonstrated that the CS chains were of the oversulfated CS-E type (Razin et al. 1982).
Identification of Mast Cell Heparin and CS as Protease-Resistant PGs
Commercially available heparin (e.g., pig mucosal heparin) is composed of free GAG chains with molecular mass ranging from ~5000 to ~20,000Da, with an average of ~15,000. However, it was shown by Horner that the heparin present in rat skin was of much higher molecular mass (~1 × 106Da), suggesting a macromolecular organization with several heparin chains attached to a protein core (Horner 1971). Proof for this notion came by showing that free GAGs were liberated from the rat skin “macromolecular” heparin by alkaline beta-elimination (Robinson et al. 1978), thus identifying the skin heparin as a PG. It was also shown that the heparin isolated from purified rat peritoneal mast cells was present in a PG form (Yurt, Leid, and Austen 1977; Levi-Schaffer et al. 1985). In addition, the CS recovered from murine BMMCs (Razin et al. 1982, 1984) and murine lymph node-derived mast cells (Bland et al. 1982) were also identified as PGs. In contrast, studies of a murine mastocytoma suggested that the heparin was predominantly present as free GAG chains (Ögren and Lindahl 1971).
A characteristic and apparently unique feature of the mast cell granule PGs is their strong resistance to protease digestion, as shown by numerous investigations (Yurt, Yurt, Leid, and Austen 1977; Bland et al. 1982; Stevens et al. 1985, 1986; Thompson et al. 1988). The most likely explanation for this phenomenon is a tight clustering of GAG chains onto the Ser-Gly repeat region of the serglycin core protein (see “Identification of the Mast Cell PG Core Protein”), which may sterically and/or electrostatically hinder the access of proteases.
GAG Expression among Mast Cell Subclasses
Mast cells show considerable heterogeneity depending on tissue location, and it is also known that different mast cell subclasses differ substantially in their expression of GAGs (Table 1). In rodents, mast cells are classified as either connective tissue-type mast cells (CTMCs) or mucosal-type mast cells (MMCs). CTMCs typically reside in skin, the peritoneal cavity and many other locations, including the intestinal submucosa. In contrast, MMCs are predominant in the mucosal layer of the intestine, where their numbers expand dramatically during, for example, parasitic infection (Metcalfe et al. 1997). CTMCs and MMCs are distinguished by their expression profile of mast cell-specific proteases, with CTMCs expressing mouse mast cell protease (mMCP)-4, -5, -6, -7 and CPA3, whereas MMCs instead express predominantly mMCP-1 and -2 (Pejler et al. 2007). With regard to GAG expression, studies performed on rat CTMCs (peritoneal, skin) showed that this population contains GAGs of heparin type (Yurt, Leid, and Austen 1977; Robinson et al. 1978). Subsequently, Enerbäck et al. (1985) demonstrated that mucosal mast cells of Nippostrongylus brasiliensis-infected rats have only a low content of heparin but instead contain GAGs of the CS type. Moreover, it was shown that the CS purified from infected mucosa was of the oversulfated CS-E type (Enerbäck et al. 1985). Stevens et al. (1986) provided additional support for the presence of oversulfated CS in MMC populations of N. brasiliensis-infected rats. However, in slight discrepancy with the studies of Enerbäck et al. (1985), the oversulfated CS was reported to contain IdoUA-2-O-sulfate-GalNAc-4-O-sulfate (“CS-diB”) disaccharides rather than the CS-E variant (Stevens et al. 1986). Notably, the presence of IdoUA-GalNAc disaccharides (regardless of the sulfation pattern) defines a GAG species as a dermatan sulfate, and it is therefore more proper to classify the mast cell GAGs containing the IdoUA-2-O-sulfate-GalNAc-4-O-sulfate disaccharide as dermatan sulfate than as CS. GAGs containing the IdoUA-2-O-sulfate-GalNAc-4-O-sulfate disaccharide (i.e., dermatan sulfate) have also been identified in PGs purified from RBL-1, a rat mast cell-like cell line (Seldin et al. 1985), and in rat BMMCs (Broide et al. 1988).
Properties of Mast Cell PGs
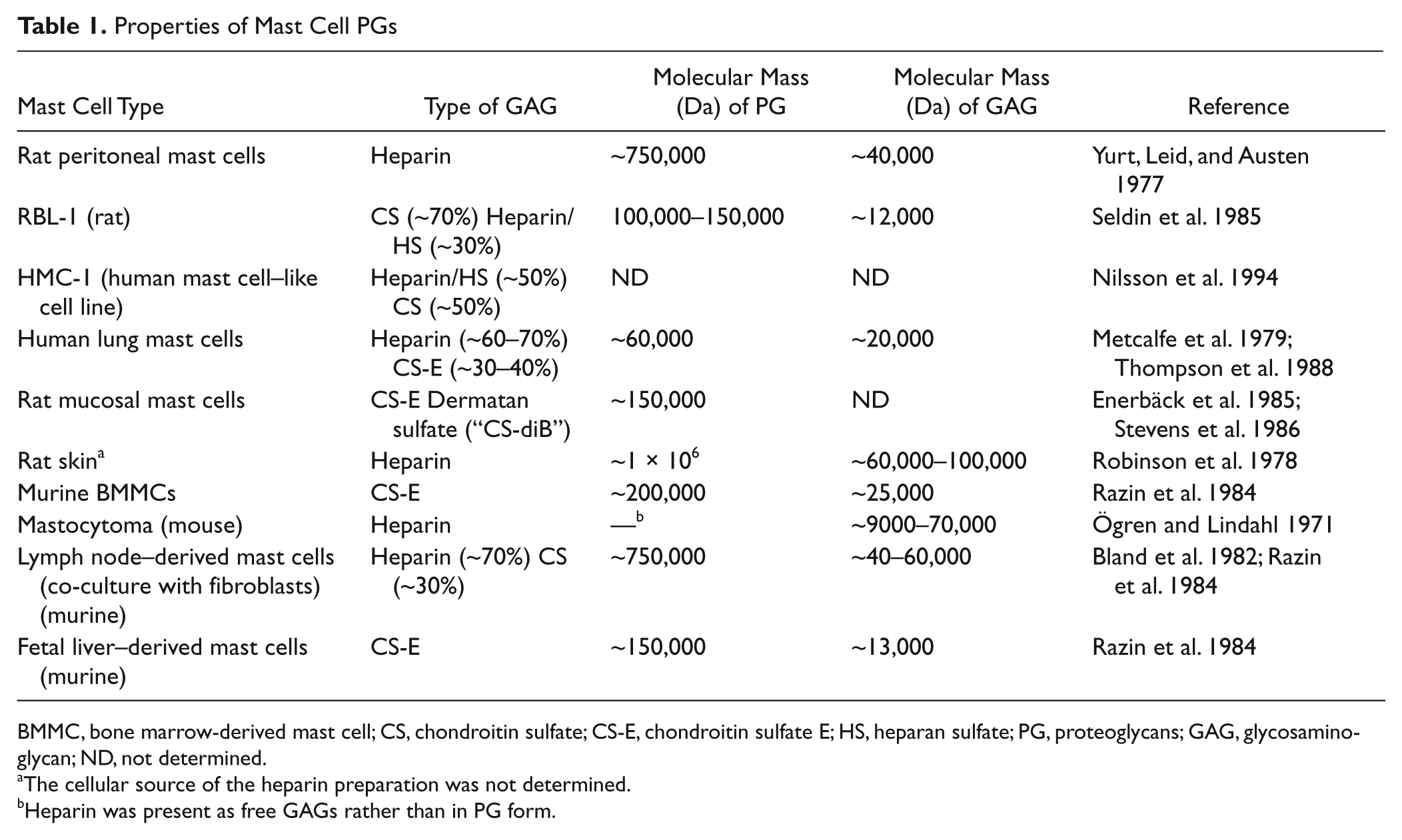
BMMC, bone marrow-derived mast cell; CS, chondroitin sulfate; CS-E, chondroitin sulfate E; HS, heparan sulfate; PG, proteoglycans; GAG, glycosaminoglycan; ND, not determined.
The cellular source of the heparin preparation was not determined.
Heparin was present as free GAGs rather than in PG form.
In mouse, most studies of mast cell PGs have been conducted on different variants of BMMCs. Based on their PG expression profile, BMMCs may be classified as being of MMC type—that is, they express predominantly CS-type PGs (Razin et al. 1982, 1984). However, based on their protease expression profile, this population of mast cells shows a typical CTMC-like phenotype—that is, they express high levels of mMCP-5, -6 and CPA3 but low levels of the MMC markers mMCP-1 and -2 (Pejler et al. 2007). Hence, BMMCs show a mixed phenotype as compared with the CTMC and MMC populations recovered in vivo. Predominant synthesis of CS over heparin/heparan sulfate (HS) has also been reported in murine embryo-derived mast cells (Dagälv et al. 2011), whereas, in contrast, fetal skin-derived mast cells synthesize predominantly heparin (Feyerabend et al. 2006; Wang et al. 2011).
In humans, the subclassification of mast cells is based on their protease content, with mast cells expressing both tryptase and chymase being classified as “MCTC,” whereas mast cells expressing tryptase only are classified as “MCT.” Although there is less knowledge of the PGs of human than of murine mast cells, the available data suggest that the PG expression profiles of the different human mast cell subclasses are less distinct as compared with the situation in rodents. As reported by Thompson et al. (1988), human lung mast cells contain a mixture of heparin (~2/3) and CS (~1/3). However, because it was not demonstrated whether the mast cell population used in the study was of the MCTC or MCT type (or a mixture), it was not clear if heparin and CS expression, respectively, was a specific feature related to either of the human mast cell subclasses. Using an indirect approach based on labeling of different mast cell subclasses with antithrombin III-conjugated gold particles, evidence was obtained suggesting that heparin was present in both MCTC- and MCT-type mast cells (Craig et al. 1993). Hence, it appears that heparin synthesis is not confined to a particular subclass of human mast cells.
In comparison to rodents, there is less knowledge of the specific PG expression profiles among human mast cell subclasses. Moreover, the PG expression profiles of the various mast cell subclasses appear to be less distinct in humans as compared with rodents.
Together, the studies cited above highlight that the nature of the GAGs synthesized differs dramatically between different mast cell subclasses. However, there is only rudimentary knowledge of the mechanisms that regulate which type of GAG will be synthesized by a given mast cell subtype. In one study, Galli and colleagues showed that prolonged incubation of BMMCs with stem cell factor (SCF; i.e., the main mast cell growth factor) favored the synthesis of heparin over CS (Tsai et al. 1991). However, the mechanism by which SCF promotes heparin synthesis is not known.
Identification of the Mast Cell PG Core Protein
Clues as to the nature of the mast cell secretory granule PG core protein came from several studies showing that purified mast cell PGs were unusually rich in Ser and Gly residues (Robinson et al. 1978; Metcalfe et al. 1980; Seldin et al. 1985; Stevens et al. 1985). Based on these findings, it was postulated early on that the GAG attachment region of the mast cell PG core protein may contain extended Ser-Gly repeats. A CS PG species containing such a Ser-Gly repeat region was subsequently cloned from a rat yolk sac tumor (Bourdon et al. 1985, 1986), and it was later shown that mRNA coding for this PG species was expressed in mast cells (Tantravahi et al. 1986; Bourdon et al. 1987). Subsequent work resulted in the cloning of a core protein of identical sequence in RBL-1, a rat mast cell-like cell line (Avraham et al. 1988), and of a human analogue in promyelocytic leukemia HL-60 cells (Stevens et al. 1988). The murine counterpart of this PG core protein was cloned both from BMMCs (Avraham et al. 1989) and from a murine mastocytoma (Kjellén et al. 1989). The most striking feature of the core protein sequence is the characteristic, extended Ser-Gly repeat region, with each Ser-Gly repeat constituting a potential GAG attachment site. Based on this unique feature, the mast cell PG core protein has been named serglycin. Notably, although the Ser-Gly repeat region can be found in serglycin of all species, its length varies from 18 and 21 amino acid residues in human and mouse serglycin, respectively, and up to 49 amino acid residues in rat serglycin. Hence, rat serglycin has the potential to accommodate a much higher number of attached GAG chains than serglycin from other species, and this may be the basis of the very high molecular weight of rat heparin PGs (Horner 1971).
Collectively, the findings described above have identified serglycin as a major PG species expressed by mast cells. However, there is a remarkable lack of knowledge with regard to the possible expression by mast cells of non-serglycin PG species. For example, there is no documentation that mast cells express typical cell surface PGs, such as syndecans or glypicans, or extracellular matrix (ECM) PGs. To address this issue, the expression of various PG species in mast cells was assessed by gene array analysis. As displayed in Table 2, serglycin is expressed at extremely high levels in mast cells, with expression levels approaching those of standard housekeeping genes such as glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and β-actin. In contrast, the expression of most other PG species is many fold lower, and in many cases, barely exceeding background noise (Table 2). Hence, serglycin is indeed by far the dominating PG species expressed by mast cells.
Gene Array a Analysis of Proteoglycan Expression in Murine Bone Marrow–Derived Mast Cells
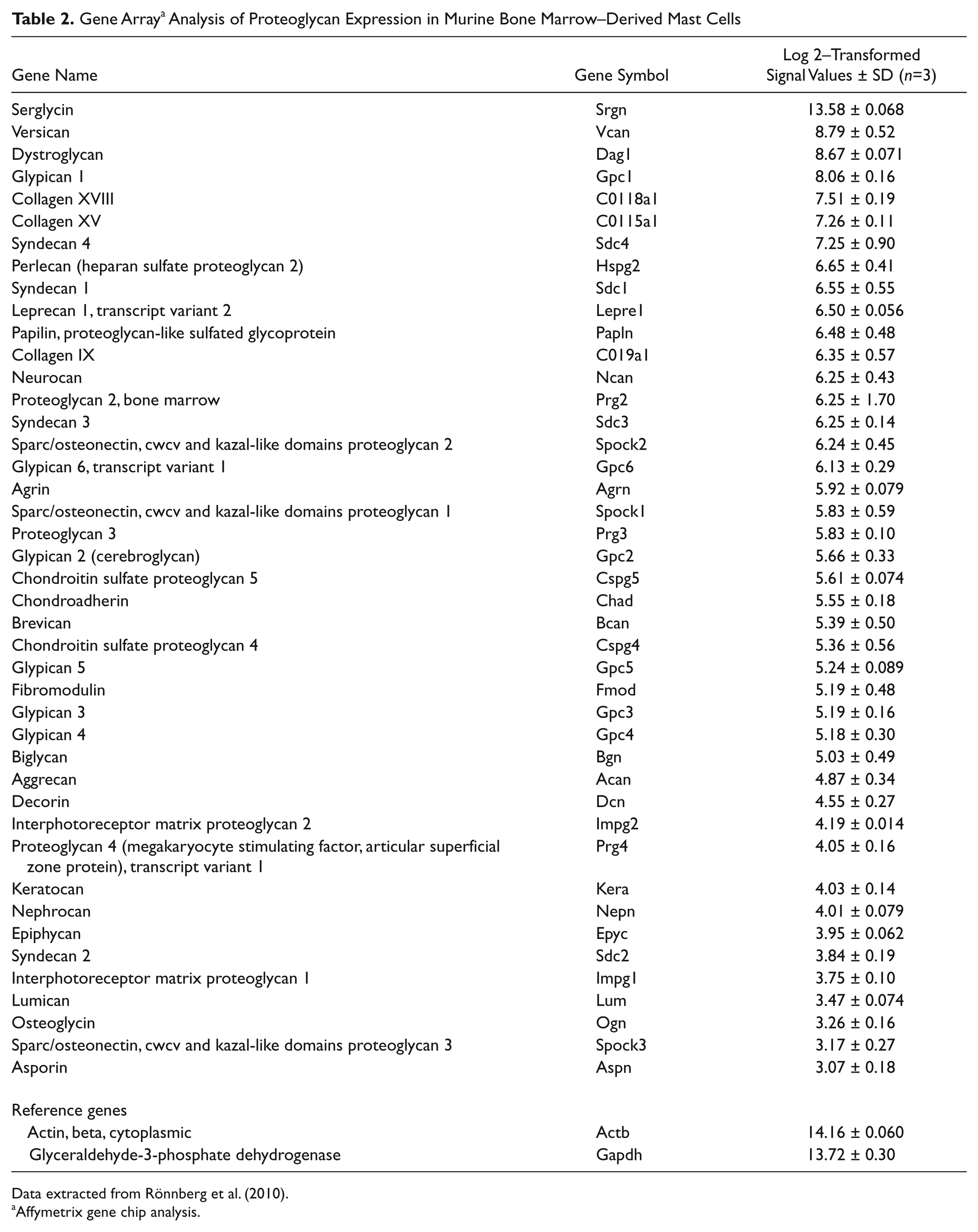
Data extracted from Rönnberg et al. (2010).
Affymetrix gene chip analysis.
Essential Role for Mast Cell PGs in Regulating Granule Storage
Important insight into the function of mast cell PGs came when the gene for serglycin was targeted in mice (Åbrink et al. 2004). The absence of serglycin was shown to result in a striking lack of metachromatic staining of both peritoneal mast cells (CTMC type) and BMMCs, showing that serglycin is essential for the morphological characteristics of mast cells (Åbrink et al. 2004) (Fig. 1). In addition, the lack of serglycin resulted in defective staining properties of MMC-type mast cells, suggesting that serglycin has a similar impact on mast cells of all subclasses (Braga et al. 2007). It was also shown that the absence of serglycin resulted in almost complete abrogation of 35S-sulfate incorporation into PGs of both CTMC- and MMC-like BMMCs, adding to the notion that serglycin is the totally dominating PG species of mast cells (Åbrink et al. 2004; Braga et al. 2007) (see also Table 2).
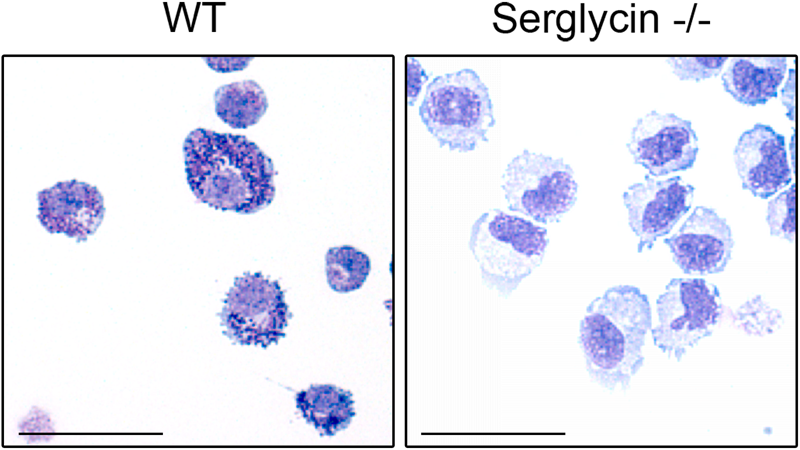
Proteoglycans of serglycin type are essential for the metachromatic staining properties of mast cells. Cytospin slides were prepared from wild-type (WT) (A) and serglycin–/– (B) bone marrow–derived mast cells and were stained with May Grünwald/Giemsa. Note the characteristic strong metachromatic, granular staining of wild-type cells and the lack of corresponding staining in cells lacking serglycin. Size bars: 50 µm.
Functionally, it has been believed for a long time that serglycin has an important role in mediating the storage of secretory granule compounds such as the various mast cell-specific proteases and biogenic monoamines (e.g., histamine) (Schwartz and Austen 1980). The basis for this notion is the high anionic charge of the GAG chains of mast cell-expressed serglycin and the matching basic charge of many of the secretory granule compounds, with the probability that serglycin may interact electrostatically with such compounds. In support of this notion, several studies performed in purified systems have revealed that various mast cell proteases show strong electrostatic interactions with heparin (Schwartz, Lewis, et al. 1981; Sayama et al. 1987; Pejler and Maccarana 1994; Hallgren et al. 2000). Moreover, mast cell proteases are released from activated mast cells in complex with PGs (Serafin et al. 1986, 1987). It is also known that histamine interacts electrostatically with heparin (Chuang et al. 2000).
Strong support for a productive interaction between serglycin and other secretory granule compounds was obtained by analysis of mast cells lacking serglycin. In accordance with earlier assumptions, it was found that the ablation of serglycin expression caused severely defective storage of granule proteases (Åbrink et al. 2004; Braga et al. 2007) and of various bioactive amines (Ringvall et al. 2008; Rönnberg et al. 2012). Notably, though, the storage-promoting effect of serglycin is not universal—that is, whereas certain proteases (mMCP-4, -5, -6 and CPA3) are strongly dependent on serglycin for storage, others (mMCP-1 and mMCP-7) are stored independently of serglycin, and yet others (mMCP-2) show an intermediate serglycin dependency (Åbrink et al. 2004; Braga et al. 2007). The differential serglycin dependency of these proteases is most likely explained by their respective surface electric charge, with strongly serglycin-dependent proteases (mMCP-4, -5 and CPA3) having a higher net positive charge than serglycin-independent compounds (mMCP-1, β-hexosaminidase) (Pejler et al. 2007).
Mast Cell PGs Affect the Enzymatic Properties of Mast Cell Proteases
In addition to affecting the granule proteases at the level of storage, mast cell PGs have been shown to influence their enzymatic properties. However, as described below, the nature of this influence differs between individual mast cell proteases.
Chymase
Mast cell chymase binds electrostatically with exceptionally high affinity to heparin, with high NaCl concentrations needed to dissociate the enzyme from heparin (Sayama et al. 1987; Pejler and Maccarana 1994). Mast cell chymase is fully enzymatically active regardless of the presence or absence of anionic GAGs. However, the strong electrostatic interaction of chymase with PGs has been shown to affect the kinetics of substrate cleavage dramatically. For example, it has been shown that the inactivating cleavage of thrombin by chymase is strongly enhanced in the presence of heparin (Pejler and Karlström 1993), and it was also shown that the cleavage of positively charged (but not uncharged) peptide substrates was stimulated by heparin and that this enhancing activity was explained by effects on the Km of chymase for the respective substrate (Pejler and Sadler 1999). Mechanistically, it has been shown that the GAG moiety of the chymase:GAG (heparin) complex serves as a bait that captures potential substrates having a matching GAG-binding (positively charged) domain, resulting in efficient presentation of such substrates to chymase and enhanced cleavage rates (Fig. 2) (Pejler and Sadler 1999). Conversely, proteins devoid of a GAG-interacting domain may be shielded from the chymase:GAG complex, which will disfavor their interaction with chymase. As examples of the latter, it has been shown that chymase in complex with heparin is protected from several macromolecular protease inhibitors, most likely explained by such a shielding mechanism (Pejler and Berg 1995). The association of chymase with heparin will thus result in protection of chymase toward inhibition by physiological inhibitors and will promote the chymase-catalyzed cleavage of substrates carrying a GAG-interacting domain. Hence, the association of chymase with GAGs will have a major impact on the biological properties of mast cell chymase. However, it should be noted that the presence of a GAG-interacting domain is not an absolute requirement for efficient proteolysis of a substrate by chymase (i.e., several substrates with no apparent GAG-interacting region are good substrates for chymase). An important example of the latter is angiotensin I (Urata et al. 1990).
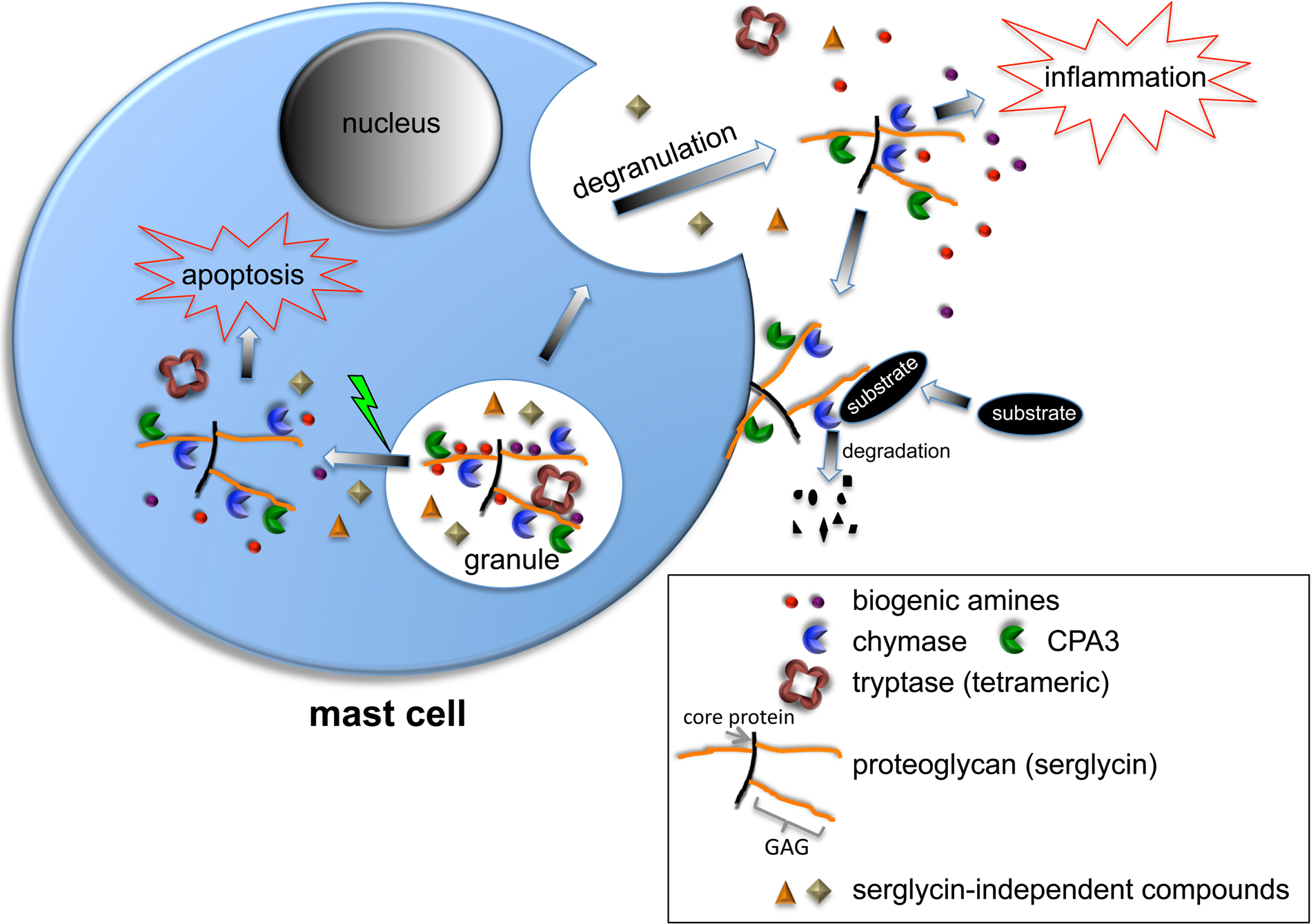
Functions of granule proteoglycans (serglycin) in mast cells. In non-activated mast cells, serglycin is located within secretory granules and promotes the storage of various proteases and biogenic amines (histamine, serotonin, dopamine). Note the tetrameric organization of tryptase, which is dependent on the interaction with serglycin. Note also that certain granule compounds are stored independently of serglycin (e.g., β-hexosaminidase). When mast cells degranulate, serglycin is released. During this process, certain compounds may be detached from serglycin (e.g., histamine), whereas others remain attached. As depicted, serglycin-protease complexes may bind to the mast cell surface after degranulation. If the granule membrane is damaged, fully active proteases in complex with serglycin enter the cytosol and can cause apoptosis. GAG, glycosaminoglycan.
Tryptase
The secretory tryptases found in mast cell granules have unique tetrameric structures in which their active sites face a narrow central pore (Fig. 2), with this macromolecular organization restricting the access of tryptase’s active sites both to substrates and inhibitors (Pereira et al. 1998). In early studies, it was observed that human β-tryptase was dependent on heparin for stability—that is, it was found that tryptase rapidly lost enzymatic activity and dissociated into inactive monomers in the absence of heparin (Schwartz and Bradford 1986; Alter et al. 1987). In subsequent studies, the effect of GAGs on the assembly of the tryptase tetramer was assessed. It was found that recombinant murine mMCP-6 and human β-tryptase were recovered as enzymatically inactive monomers in the absence of GAGs (Hallgren et al. 2000, 2005). However, when heparin was added, the respective tryptases underwent tetramerization and became enzymatically active (Hallgren et al. 2000, 2001, 2005). Importantly, tetramerization and enzymatic activation were only seen if heparin was added to the inactive monomers at low pH (<6.5) (Hallgren et al. 2000). In line with this, optimal stabilization of tryptase tetramers requires a slightly acidic pH (Ren et al. 1998). A likely explanation for these findings is that the electrostatic interaction of tryptase with heparin is mediated by His residues, which become positively charged at slightly acidic pH while being deprotonated and thereby uncharged at neutral pH. Indeed, a subsequent study identified a patch of His residues on the surface of mMCP-6 being responsible for the interaction with heparin, leading to tetramerization and gaining of enzymatic activity (Hallgren et al. 2004). The collected findings thus indicate that mast cell tryptases depend on anionic PGs present within mast cell granules for both the gaining and maintaining of enzymatic activity.
CPA3
Differently to chymases and tryptases, there is to date no documentation showing that CPA3 depends on direct contact with GAGs for expression of catalytic activity, for substrate presentation, or for protection against protease inhibitors. On the other hand, there is substantial evidence for an indirect dependence of CPA3 on mast cell PGs, through the finding that the processing of pro-CPA3 into an active enzyme is severely defective in mast cells lacking either serglycin (Henningsson et al. 2006) or N-deacetylase/N-sulfotransferase (NDST)-2 (Henningsson et al. 2002), with NDST-2 being an enzyme critical for sulfation of the heparin chains attached to the serglycin protein core (see also “Expression of GAG-Modifying Enzymes in Mast Cells”). A dissection of the underlying mechanism indicated that the processing of pro-CPA3 into catalytically active enzyme was catalyzed by cathepsin E present within the mast cell granules and that the storage of cathepsin E within granules is dependent on heparin (Henningsson et al. 2005). Thus, although CPA3 shows no direct dependency on PGs for its enzymatic properties, processing of pro-CPA3 into active protease is strongly dependent on proteases that, in turn, rely on serglycin for their presence within mast cell granules.
Mast Cell PGs Regulate the Release of Secretory Granule Compounds
In addition to promoting the storage of a number of compounds in secretory granules, mast cell PGs can also have an impact on the fate of the respective compounds after degranulation. As an important example, there is evidence suggesting that tryptases having high affinity for heparin (mMCP-6) are retained in the close vicinity of the degranulated mast cell, whereas tryptases with low affinity for heparin (mMCP-7) diffuse away from the site of degranulation and may reach the circulation (Ghildyal et al. 1996). Most likely, the retention of mMCP-6 at the site of degranulation is explained by its tight association with granule PGs, even after release to the extracellular space, and binding of exocytosed PG-mMCP-6 complexes to either the mast cell surface or to the surrounding ECM. Conversely, due to its low affinity for granule PGs, diffusion of mMCP-7 away from the site of degranulation is not hindered. In an analogous fashion, chymases with high affinity for GAGs (e.g., human chymase, mMCP-4) and CPA3 tend to attach to the mast cell surface after degranulation (Fig. 2), whereas chymases with low affinity for heparin (mMCP-1) are recovered in the circulation (Schwartz, Riedel, et al. 1981; Tchougounova and Pejler 2001; Brown et al. 2003).
At the acidic pH prevalent within the mast cell granules, histamine is protonated and can thereby engage in electrostatic complexes with heparin (Chuang et al. 2000). When PG-histamine complexes are released into the extracellular space, the rise in pH causes deprotonation of histamine, leading to its detachment from the PG and diffusion away from the site of degranulation and into the circulation (Fig. 2). Deprotonation may also regulate the release of various proteins that depend on His residues for tight binding to mast cell PGs. For example, we have shown that the strong interaction of mMCP-6 with heparin is dependent on a His cluster present on the surface of the molecule (Hallgren et al. 2004). In the acidic milieu of the granules, these His residues are protonated and will mediate strong binding of mMCP-6 to the granule PGs. However, when the mMCP-6:PG complexes are released, their exposure to an elevated pH will cause His deprotonation, leading to weakened interaction of mMCP-6 with the PG and consequent dissociation of the complex. Similarly, it has been demonstrated that granulocyte macrophage colony-stimulating factor (GM-CSF) depends on His residues for interaction with heparin (Sebollela et al. 2005), and it has also been shown that GM-CSF can be produced by mast cells (de Vries et al. 2011). Although not proven, it is thus possible that GM-CSF forms complexes with PGs within acidic compartments of the mast cell (i.e., under conditions where the His residues of GM-CSF are protonated and thereby prone to engage in electrostatic interactions with PGs). When such putative complexes are released from the mast cells, deprotonation of GM-CSF will cause its dissociation from the PG. However, the functional impact of such a mechanism in a mast cell context remains to be examined.
Extracellular Functions of Mast Cell PGs
Although a main function of mast cell secretory granule PGs is to act as carriers for other granule compounds during their storage and exocytosis, it cannot be excluded that granule PGs have additional functions after their release to the extracellular space. One obvious possibility would be that mast cell heparin, being a powerful anticoagulant, has an important role in regulating blood coagulation. However, because no blood clotting abnormalities have been noted in mice lacking mast cell heparin (Forsberg et al. 1999; Humphries et al. 1999), this remains to be demonstrated. Another scenario is that mast cell PGs may interact with various compounds present in the extracellular space under inflammatory conditions. As an important example, many of the chemokine family members are known to interact strongly with anionic GAGs (Proudfoot 2006). It is thus likely that PGs secreted by mast cells will become engaged in productive interactions with chemokines secreted by other inflammatory cells during an inflammatory reaction, and this could have diverse functional consequences (Proudfoot 2006). For example, it has been suggested that mast cell heparin may regulate eosinophil recruitment by binding to CCL11 and thereby protecting the chemokine from proteolytic degradation (Ellyard et al. 2007). It has also been shown that anionic GAGs can affect the rate of diffusion of chemokines within the ECM and can influence the kinetics of chemokine release from the tissue into the circulation (Flaumenhaft et al. 1990; Tanino et al. 2010). Possibly, heparin secreted by mast cells could thus modulate such processes (e.g., by competing with HS/CS PGs anchored within the ECM for binding to chemokines). Other possibilities are that PGs released from mast cells may regulate the complement system (Skliris et al. 2011) or can activate bradykinin (Oschatz et al. 2011).
Mast Cell Secretory Granule PGs Can Promote Apoptosis
Mast cell granules contain remarkably high amounts of preformed, fully active neutral (pH optimum) proteases, several of which (see “Essential Role for Mast Cell PGs in Regulating Granule Storage”) are strongly dependent on serglycin for storage (Åbrink et al. 2004; Braga et al. 2007). If the granule membranes become damaged, large amounts of the granule-contained proteases may thus enter the cytosol (Fig. 2). This may create a hazardous situation, due to the potential proteolytic damage that these proteases may cause to cytosolic proteins. Among the potential ensuing scenarios is the possibility that granule proteases may cause apoptosis, either by proteolytic activation of proapoptotic proteins or by degradation of antiapoptotic compounds. On the basis of this potential scenario, we hypothesized that mast cells are sensitive to apoptosis induced by permeabilization of the secretory granules, and because many of the granule proteases rely on serglycin for storage, we also hypothesized that apoptosis induced along this pathway may be dependent on serglycin. Indeed, we showed that mast cells were highly sensitive to apoptosis induced by granule permeabilization induced by lysosomotropic agents (Melo, Lundequist, et al. 2011; Melo, Waern, et al. 2011) and that mast cells lacking serglycin were markedly less sensitive to apoptosis induced by this pathway than were wild-type cells (Melo, Waern, et al. 2011). In agreement with our hypothesis, the apoptosis-promoting activity of serglycin depended on its complex-bound proteases (Melo, Waern, et al. 2011).
In a subsequent study, the impact of serglycin on cell death in mast cells was examined further. These studies showed that, whereas wild-type cells preferably die by apoptosis in response to numerous different stimuli, mast cells lacking serglycin predominantly undergo necrotic cell death (Melo et al. 2012). The necrotic cell death in serglycin–/– cells was explained by defective degradation of poly(ADP-ribose) polymerase 1 (PARP-1). PARP-1 is a DNA repair enzyme that undergoes proteolytic degradation during the course of apoptotic cell death, leading to impaired DNA repair and the typical DNA fragmentation seen in apoptosis. However, if PARP-1 degradation is impaired (as in serglycin–/– mast cells), sustained DNA repair will lead to extensive energy depletion and consequent necrotic cell death (Ha and Snyder 1999). Hence, serglycin regulates the levels of PARP-1 in response to cellular stress and can thereby be a decisive factor determining whether mast cells die by apoptosis or necrosis.
Expression of GAG-Modifying Enzymes in Mast Cells
The expression by mast cells of the various sulfotransferases and other GAG-modifying enzymes involved in heparin and CS synthesis has been studied to some extent. A crucial step in the sulfation of heparin (and HS) is the deacetylation and subsequent N-sulfation of selected N-acetylglucosamine units. This reaction is catalyzed by the bifunctional NDSTs, an enzyme family composed of four family members (NDST-1–4) (Kjellén 2003). Based on the high expression of NDST-2 in a murine mastocytoma (Kusche-Gullberg et al. 1998), it was hypothesized that this NDST isoform may be of particular importance for heparin biosynthesis in mast cells. Indeed, when NDST-2 was knocked out, it was found that heparin synthesis in mast cells was completely abrogated (Forsberg et al. 1999; Humphries et al. 1999). Moreover, the absence of NDST-2 led to similar storage defects as those seen in mast cells lacking serglycin, that is, markedly defective storage of mMCP-4, -5, -6; CPA3; and histamine in CTMCs (Forsberg et al. 1999; Humphries et al. 1999). However, apart from the remarkable effects on CTMCs, the absence of NDST-2 does not appear to result in any other obvious phenotypic effects, suggesting that NDST-2 function is largely confined to mast cells of the CTMC subclass. This is in sharp contrast to the absence of NDST-1, which is associated with neonatal lethality (Ringvall et al. 2000). The tight association between NDST-2 expression and mast cell phenotype is also underscored by the continuously upregulated expression of the NDST-2 gene during the process of mast cell differentiation from bone marrow precursors, whereas, in contrast, NDST-1 expression is not linked to mast cell maturation (Duelli et al. 2009).
There is evidence that the heparin PGs present in mast cell granules, after completed biosynthesis, may be subject to regulation by endoglycosidases. This was first indicated through the identification of a heparin-degrading enzyme in a murine mastocytoma (Ögren and Lindahl 1971); this enzyme was implicated in degradation of the native heparin chains into fragments of similar sizes as those present in standard, commercially available heparin preparations. Later, the presence of a heparin-degrading enzyme (heparanase) within mast cell granules was demonstrated (Bashkin et al. 1990). After its exocytosis from mast cells, this enzyme was shown to degrade HS present in the ECM. Through the work of Li and colleagues, heparanase-deficient animals have been generated (Zcharia et al. 2009). Interestingly, the absence of heparanase in fetal skin-derived mast cells resulted in increased molecular size of the heparin chains, accompanied by stronger metachromatic staining and increased levels of stored proteases (Wang et al. 2011). These findings indicate that heparanase has a cell-intrinsic homeostatic function in regulating the size and amount of granule-contained heparin PG, thereby having an indirect effect on the storage of PG-associated granule compounds.
A characteristic feature of the CS expressed by BMMCs and mucosal mast cell populations is the preponderance of oversulfated variants containing the GlcUA-GalNAc-4,6-di-O-sulfate disaccharide species. Accordingly, mast cells have been shown to express enzymes catalyzing the 4-O-sulfation of GalNAc (i.e., GalNAc-4-O-sulfotransferases [C4STs]) and the enzyme responsible for the subsequent 6-O-sulfation of 4-O-sulfated GalNAc (i.e., GalNAc(4S)-6-O-sulfotransferase [GalNAc4S6ST]). Of the different isoforms of C4STs, it was found that mature mast cells express predominantly C4ST-1, and it was also noted that the expression levels of C4ST-1 correlated positively with the extent of mast cell maturation. Also, the expression of GalNAc4S6ST was strongly induced during the maturation of mast cells from bone marrow precursors (Ohtake et al. 2008; Duelli et al. 2009). In contrast, the expression of C4ST-2 showed no such correlation, and the expression of chondroitin-6-sulfotransferase (C6ST) was high in bone marrow precursor cells but was strongly downregulated as the cells matured into the mast cell phenotype (Duelli et al. 2009). In accordance with the importance of a high extent of CS sulfation to promote granule protease storage, the ablation of GalNAc4S6ST in mice caused impaired storage of mMCP-6 and CPA3 in BMMCs (Ohtake-Niimi et al. 2010).
Conclusions
Although it has been known for decades that mast cell granules contain highly sulfated PGs, only during recent years has the true function of these PGs started to become unraveled. As discussed in this review, recent studies have revealed a crucial role for granule-contained serglycin PGs in the packaging of secretory granule compounds, as well as a role for the granule PGs in regulating the activities and release of these compounds. To date, most studies delineating functions of mast cell PGs (serglycin) have been carried out using in vitro approaches. However, it will be important for future studies to address the in vivo role of mast cell PGs through studies on mice in which serglycin or enzymes involved in the sulfation of the GAGs attached to the serglycin core protein have been conditionally targeted in mast cells. In particular, considering the strong implication of mast cells in various pathologies, it will be important to evaluate such mice in various models for disease. Moreover, although the importance of serglycin-type PGs for mast cells has been studied extensively, there is remarkably little knowledge of the possible expression and function of PG species other than serglycin in mast cells. We anticipate that future research will address this issue.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors of this article receive support from The Swedish Research Council, The Swedish Cancer Foundation, and King Gustaf V’s 80-year Anniversary Fund.