
Abstract
The aim of this study was to evaluate HDPR1 expression in esophageal squamous cell carcinoma (ESCC) and the relationship between HDPR1 and beta-catenin by immunohistochemical analysis. The clinical relevance of these proteins was also analyzed. Immunohistochemistry was performed on paraffin-embedded tissue specimens from 184 ESCC patients to detect the expression of HDPR1 and beta-catenin. The correlation between the results of immunoexpression and the clinicopathologic features was processed statistically. Increased cytoplasmic and nuclear HDPR1 expression was noted in 100 (54.3%) and 131 (71.2%) of 184 specimens, respectively. Statistical analysis showed significant associations of cytoplasmic HDPR1 with regional lymph node metastasis (p = 0.021) and P-stage (p = 0.004). The increased nuclear staining was only correlated with P-stage (p = 0.047). Significant associations of coexpression of cytoplasmic and nuclear HDPR1 with regional lymph node metastasis (p = 0.015) or P-stage (p = 0.002) were observed. Enhanced cytoplasmic expression of HDPR1 was positively correlated with increased cytoplasmic but not reduced membranous beta-catenin expression (r = 0.239, p = 0.027 and r = 0.126, p = 0.089, respectively). These finding suggested that cytoplasmic HDPR1 protein expression was associated with tumor malignant progression via beta-catenin accumulation. It implicated that cytoplasmic HDPR1 expression may serve as a potential predictive factor for lymph node metastasis and tumor development in ESCC.
Esophageal squamous cell carcinoma (ESCC), one of the most aggressive carcinomas in China, is associated with poor patient survival despite aggressive treatment (Ke 2002; Lerut et al. 1999; Parkin et al. 2005). The 5-year survival rate of patients with ESCC is less than 35% due to the difficulty of diagnosis at an early stage, local invasion, and lymph node metastasis at an advanced stage (Lerut et al. 1999; Shimada et al. 2003). Thus, identifying a sensitive and representative biological marker is extremely important for diagnosis and evaluating prognosis.
Dapper (Dpr), which was first found as a Dvl interacting protein, plays a crucial role in normal vertebrate development. Depletion of maternal Dpr RNA from Xenopus embryos leads to a loss of notochord and head structures (Cheyette et al. 2002). A Dpr homolog, named Frodo, is essential for normal eye and neural tissue development (Gloy et al. 2002). Hikasa and Sokol (2004) showed that Frodo and Dpr act synergistically during head development and are required for neural development in Xenopus. In zebrafish, orthologs of Dpr and Frodo are expressed during embryogenesis and have an important role in patterning the neural plate and several mesodermal derivatives (Gillhouse et al. 2004). All the reports mentioned above showed that Dpr and its homologs are involved in growth and development via affecting beta-catenin, an important component of the Wnt signaling pathway.
Moreover, the Dpr gene was evolutionally conserved from fish to humans (Fisher et al. 2006; Katoh and Katoh 2003; Waxman et al. 2004; Zhang et al. 2006), implying that it might have important functions. However, little is known about the roles of Dpr in human diseases.
The human homologue of Dpr (HDPR1) was identified and detected in a series of cancers (Katoh and Katoh 2003, 2005). Recently, it was reported that HDPR1 is downregulated in hepatocellular carcinoma (HCC) and non-small-cell lung cancer (NSCLC; Yang et al. 2010; Yau et al. 2005). However, the expression pattern and functional mechanism of HDPR1 in other cancers, such as ESCC, remain unclear. In this study, we examined HDPR1 expression in ESCC and analyzed the relationship between HDPR1 expression and clinicopathological factors of patients with ESCC. The relationship between HDPR1 and beta-catenin expression was also analyzed.
Patients and Methods
Patients and Tissue Specimens
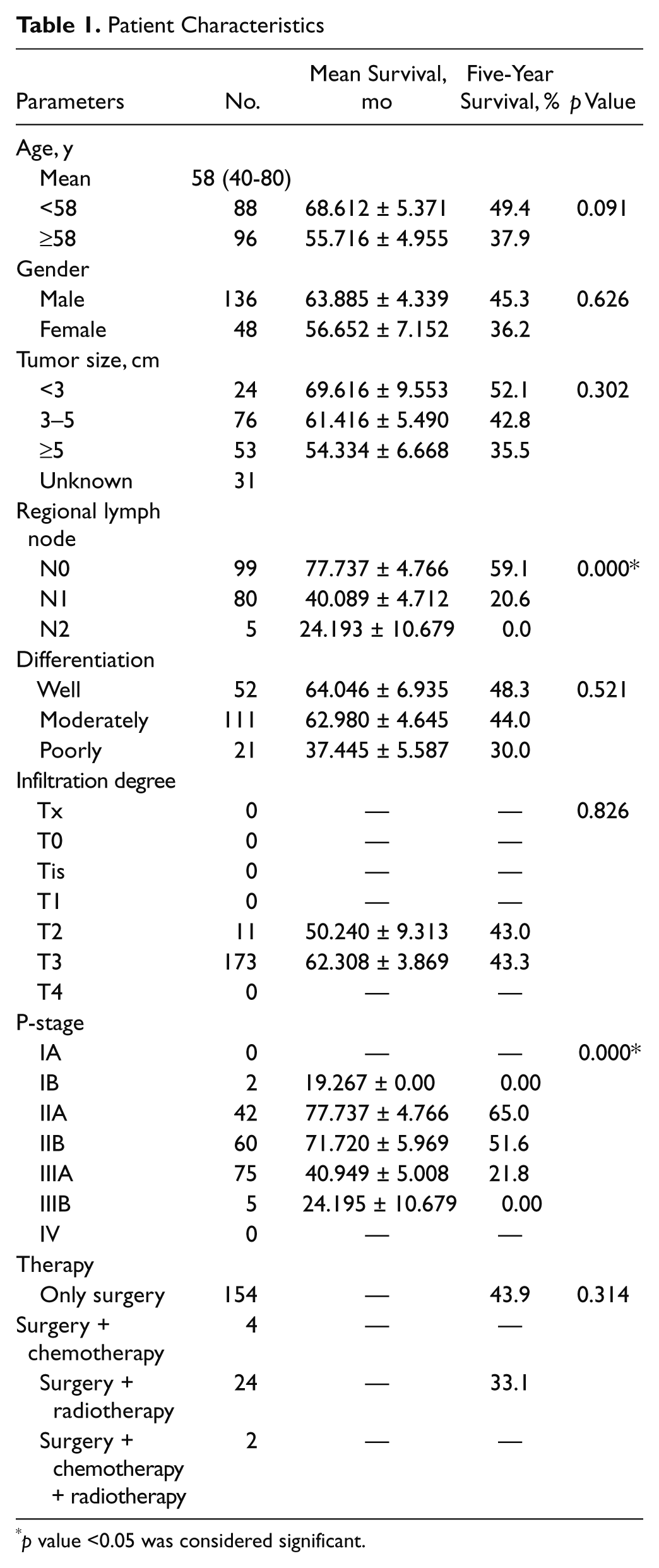
For the retrospective study, tumor specimens were obtained from 184 patients with primary ESCC at Shantou Central Hospital from 2000 to 2006. The patients were 136 men and 48 women (median age, 58 years; range, 40-80). We retrieved information, including sex, age, stage of disease, and histopathological factors, from the patient hospital charts. Patients’ data are summarized in Table 1. The patients who died in the immediate postoperative period from complications or from other tumors or other causes were excluded. The local ethics committee granted ethical approval for the study.
Patient Characteristics
p value <0.05 was considered significant.
TMA Construction
For TMA construction, a hematoxylin and eosin–stained slide from each block was used to define the representative regions of each tissue. Two replicate tissue cores were obtained from each specimen. The 1.8-mm diameter TMA cores ranged in length from 1.0 to 3.0 mm depending on the depth of the tissue in the donor block and were each spaced at 0.8 mm from core center to core center. Each core was precisely arrayed into a new paraffin block. After construction, serial 4-µm-thick tissue sections were cut, and hematoxylin and eosin staining was performed on the initial slide to verify tissue sampling and completeness. Unstained sections were baked overnight at 56C in preparation for immunohistochemistry.
Immunohistochemical Staining
Immunohistochemical staining was performed on 4-µm-thick tissue microarray sections from the TMA blocks. After deparaffinization and rehydration, all sections were treated with microwaves (10 min) in 0.01 mol/L sodium citrate buffer (pH 6.0) for antigen retrieval. To block endogenous peroxidase activity, we incubated the sections with 10% normal goat serum in PBS for 15 min at room temperature. Then all sections were incubated at 4C overnight with rabbit polyclonal antibody to HDPR1 (1:100; Abcam, Cambridge, UK) and mouse monoclonal beta-catenin (1:100; ZSGB-BIO, Beijing, China). Then slides were detected by the application of the SuperPic Ture Polymer Detection kit and Liquid DAB Substrate kit (Zymed/Invitrogen, San Francisco, CA) and counterstained with hematoxylin, dehydrated, and mounted.
Evaluation of Immunostaining
Immunohistochemical staining was assessed by a pathologist who had no knowledge of clinical characteristics of the patients. The immunoreactive score was determined by the intensity and extension of the positive staining. The intensity of positive staining was scored 3 for strong staining, 2 for moderate staining, 1 for weak staining, and 0 if negative. The extent of positivity was estimated on scale of 0 to 4: 0 = positive staining in 0% to 5% of cells, 1 = positive staining in 6% to 25%, 2 = positive staining in 26% to 50%, 3 = positive staining in 51% to 75%, and 4 = positive staining in 76% to 100%. If the positive staining was homogeneous, the final score was achieved by multiplication of the two scores above, producing a total range of 0 to 12. If the staining was heterogeneous, it was scored as follows: each component scored independently and summed for the results. For example, a specimen containing 25% tumor cells with moderate intensity (1 × 2+ = 2), 25% tumor cells with weak intensity (1 × 1+ = 1), and 50% tumor cells without immunoreactivity received a final score of 2 + 1 + 0 = 3.
For HDPR1 scoring, a case with a total score ≤4 was labeled “negative” and >4 was labeled “positive,” according to the scores of normal esophageal epithelium tissues, which were all ≤4. Because the normal esophageal epithelium was positively stained by beta-catenin at the cell membrane but negative in the cytoplasm, the membranous beta-catenin expression was defined as “reduced” if the score was ≤4 and “normal” if the score was >4. Cytoplasmic beta-catenin expression was defined as “increased” if the score was >4 and “normal” if the score was ≤4.
Statistical Analyses
The correlations between immunohistochemical results and clinicopathologic variables were analyzed by Pearson’s χ2 test. Survival curves were estimated by the Kaplan-Meier method and compared by the log-rank test. A p value less than 0.05 was considered statistically significant. All the statistical analyses were performed with SPSS 13.0 software (version 13.0; SPSS, Inc., an IBM Company, Chicago, IL).
Results
Patient Characteristics
Table 1 shows the demographic data and the results of survival analysis in 184 ESCC patients (UICC-TNM classification, 7th edition, 2009). Because statistical analysis showed that there was no survival advantage with the use of radiotherapy or chemotherapy compared with the surgery-alone group, the patients’ survival was analyzed together. The 5-year survival was 52.2%, with a median survival of 46.6 months. Kaplan-Meier method analysis revealed that survival time of the patients was significantly associated with regional lymph node metastasis (p = 0.000) and P-stage (p = 0.000).
Expression of HDPR1 in ESCC
HDPR1 expression was rarely detected in normal squamous cell epithelia (Fig. 1A). Strong immunoreactivity for HDPR1 was observed both in the cytoplasm and/or nucleus of ESCC cells (Fig. 1B,C). Increased cytoplasmic and nuclear HDPR1 expression was noted in 100 (54.3%) and 131 (71.2%) of 184 tumor specimens, respectively.
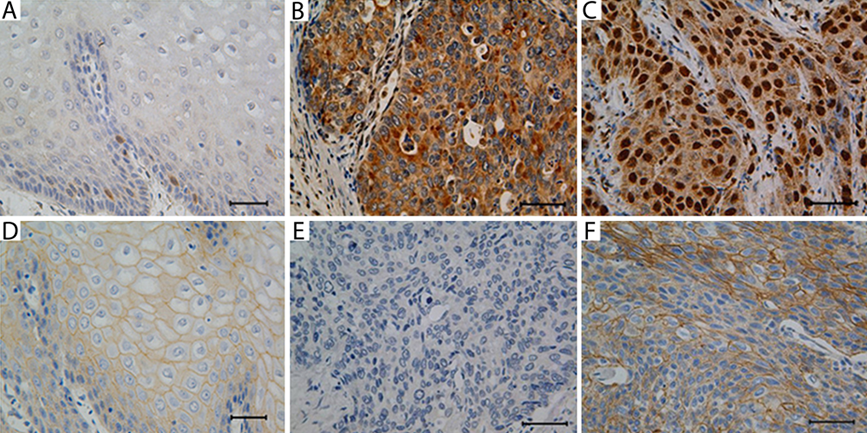
Immunohistochemical analysis of HDPR1 and beta-catenin expression in esophageal squamous cell carcinoma (ESCC). The HDPR1 expression was rarely detected in the normal epithelia (A, ×400). The HDPR1 expression was detected in the cytoplasm (B, ×400) or nucleus (C, ×400) of ESCC tissue. The beta-catenin expression was detected mainly in the cytomembrane and weakly in the cytoplasm in normal epithelia (D, ×400). Reduced expression of beta-catenin was observed in ESCC (E, ×400), although some samples showed enhanced expression (F, ×400). Scale bars: 50 µm.
Statistical analysis showed significant associations of cytoplasmic positive staining of HDPR1 with regional lymph node metastasis (p = 0.021) and P-stage (p = 0.004). For negative cases of the regional lymph node metastasis, the positivity of HDPR1 was close to the negativity (53.5% vs 46.5%). However, for the positive cases of regional lymph node metastasis, the HDPR1 positivity was much higher than the negativity (54% vs 36.5%). A similar tendency was also observed in the P-stage cases. In the stage III group, HDPR1 positivity was 66.2% and HDPR1 negativity was 33.8%. Other factors, such as age, gender, tumor size, differentiation, and infiltration degree, had no statistical associations with cytoplasmic expression of HDPR1.
We also investigated the associations between nuclear expression of HDPR1 and clinicopathological features. The increased nuclear staining of HDPR1 was positively correlated with P-stage (p = 0.047). Enhanced nuclear HDPR1 expression cases were found in 65.8% of stage I/II and 78.8% of stage III cases. No significant association was observed between nuclear HDPR1 expression and other clinicopathological factors (Table 2).
Relationship between HDPR1 Protein Expression and Clinicopathological Features in Esophageal Squamous Cell Carcinoma
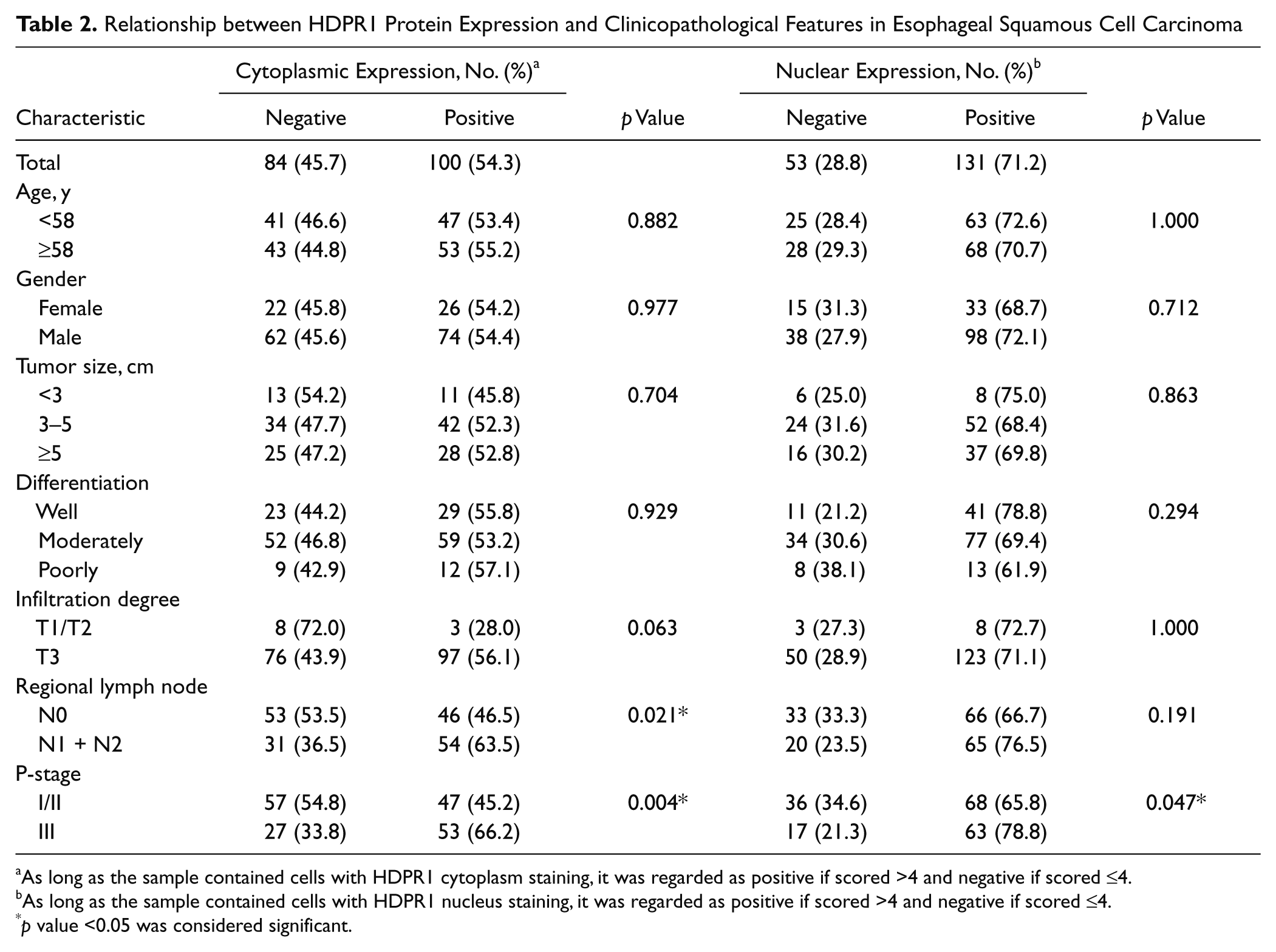
As long as the sample contained cells with HDPR1 cytoplasm staining, it was regarded as positive if scored >4 and negative if scored ≤4.
As long as the sample contained cells with HDPR1 nucleus staining, it was regarded as positive if scored >4 and negative if scored ≤4.
p value <0.05 was considered significant.
Correlation between HDPR1 Subcellular and Clinicopathologic Parameters
To understand the function of HDPR1 in ESCC, we also investigated the relationships between HDPR1 subcellular distribution and the clinicopathologic parameters. Before analysis, all the cases were divided into four groups according to the stained score and the location of HDPR1, including nucleus and cytoplasm positive, only cytoplasm positive, only nucleus positive, and negative. Table 3 shows that HDPR1 subcellular distribution is associated with regional lymph node metastasis (p = 0.015) and P-stage (p = 0.002). The percentage of HDPR1 expression in both cytoplasm and nucleus was 49.4% for the regional lymph node metastasis cases and 52% for the stage III cases, which was significantly higher than other groups. No significant association was observed between HDPR1 subcellular distribution and other clinicopathological factors (Table 3).
Relationship between HDPR1 Protein Distribution and Clinicopathological Features in Esophageal Squamous Cell Carcinoma
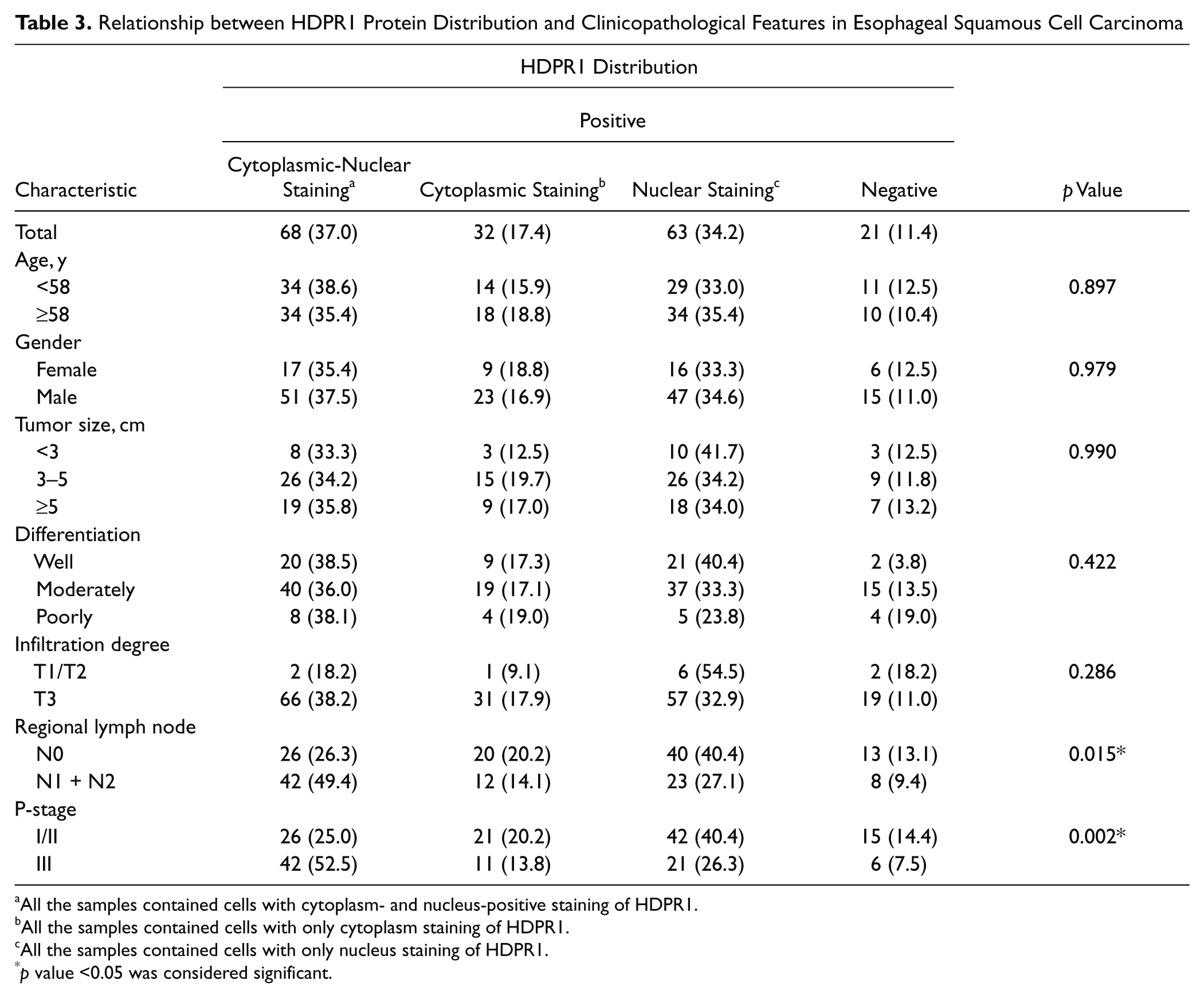
All the samples contained cells with cytoplasm- and nucleus-positive staining of HDPR1.
All the samples contained cells with only cytoplasm staining of HDPR1.
All the samples contained cells with only nucleus staining of HDPR1.
p value <0.05 was considered significant.
Association of HDPR1 Expression and Subcellular Distribution with Survival Rate of ESCC Patients
The association of HDPR1 expression level with survival rates of ESCC patients was evaluated by the Kaplan-Meier method. Unfortunately, no statistical significance was found between cytoplasmic (p = 0.767; Fig. 2A) or nuclear (p = 0.393; Fig. 2B) expression of HDPR1 and survival rate of ESCC patients. Compared with other groups, the 5-year survival rate was highest in patients with HDPR1 expression in the nucleus and cytoplasm (5-year survival = 57.3%); by contrast, the lowest survival rate was in patients with only cytoplasm-positive expression (5-year survival = 24.4%). The survival rates of the other two groups were similar (44.3% for the negative group and 40.2% for nuclear-positive group). However, we observed no statistical significance (p = 0.176; Fig. 2C).
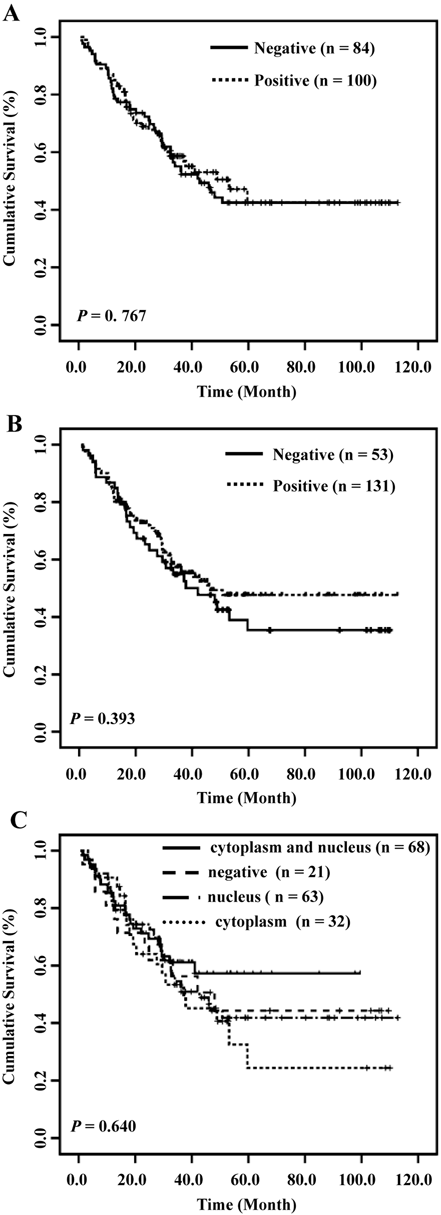
Overall survival of patients with esophageal squamous cell carcinoma (ESCC) in relation to HDPR1. Kaplan-Meier survival curve for (A) cytoplasmic HDPR1, (B) nuclear HDPR1, and (C) subcellular distribution of HDPR1.
Relationship between HDPR1 and Beta-Catenin
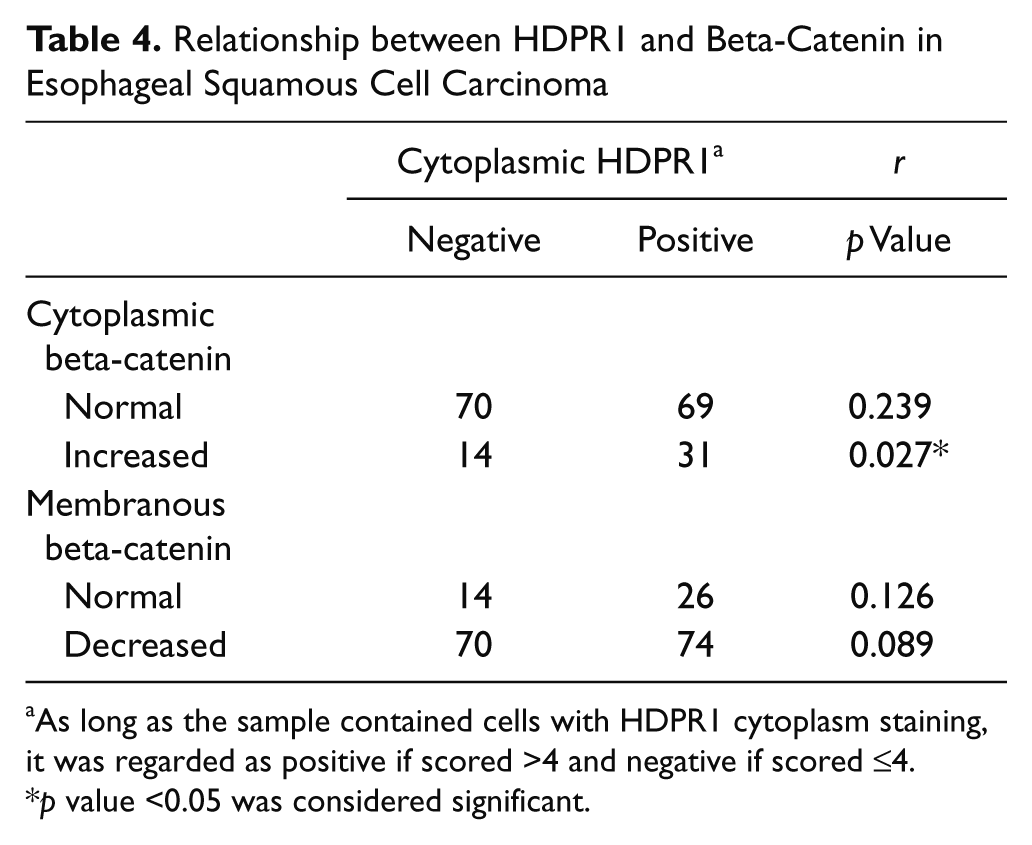
It was reported that HDPR1 could inhibit or promote the Wnt pathway through affecting beta-catenin binding to the promoter (Cheyette et al. 2002; Gao et al. 2008; Gloy et al. 2002; Park et al. 2006). To investigate the association between HDPR1 and beta-catenin in ESCC, we used immunohistochemistry and analyzed their correlation. In normal squamous cell epithelia, beta-catenin expression was detected only in membrane, but reduced membranous (145/184, 71.2%) and enhanced cytoplasmic (43/184, 23.4%) beta-catenin expression was observed in ESCC, whereas the nucleus was negative stained (Fig. 1D–F; Supplemental Table S1). No significant association was observed between beta-catenin expression and clinicopathological features as well as the survival rate in ESCC (Supplemental Table S1 and Supplemental Fig. S1); however, because beta-catenin is an important component of the Wnt pathway in cancer (Ilyas 2005; Nelson and Nusse 2004), we analyzed the association between HDPR1 and beta-catenin. Table 3 shows that enhanced cytoplasmic expression of HDPR1 is positively correlated with increased cytoplasmic (r = 0.239, p=0.027) but not with reduced membranous beta-catenin expression (r = 0.126, p = 0.089).
Discussion
HDPR1, a member of the Dapper family, was reported as an important regulated factor in the Wnt pathway (Cheyette et al. 2002; Gao et al. 2008; Gloy et al. 2002; Park et al. 2006; Zhang et al. 2006). Considering its important effect on the Wnt pathway (Ilyas 2005), it was supposed that HDPR1 might play an important role in cancer, including ESCC. However, research has rarely reported about the functional mechanism of HDPR1 in ESCC.
The recent data suggest that decreased HDPR1 expression is associated with the progression of NSCLC and HCC (Yang et al. 2010; Yau et al. 2005). In our study, however, the expression pattern of HDPR1 in ESCC was totally distinguished from that in other cancers mentioned above. In ESCC, HDPR1 was overexpressed in tissues as well as cell lines (Supplemental Fig. S2) and associated with clinicopathological factors. Immunohistochemistry analysis showed that increased cytoplasmic HDPR1 was positively associated with regional lymph node metastasis (p = 0.021) and P-stage (p = 0.004; Table 2), indicating that cytoplasmic HDPR1 might contribute to tumor metastasis, resulting in malignant progression of ESCC. Unlike cytoplasmic HDPR1, increased expression of nuclear HDPR1 was only correlated with P-stage (p = 0.047; Table 2); however, patients with coexpression of HDPR1 in the cytoplasm and nucleus appeared to have a higher rate of regional lymph node metastasis (p = 0.015) and stage III (p = 0.002) than solo expression (Table 3), suggesting that nuclear HDPR1 might have a connection with cytoplasmic HDPR1, which might facilitate tumor metastasis and progression. However, survival analysis did not show statistical significance (Figure 2). The results above indicate that HDPR1, especially cytoplasmic HDPR1, might be a predictive factor for regional lymph node metastasis and tumor development in ESCC.
HDPR1, as a molecular switch for Wnt signaling, could promote or block beta-catenin degradation (Teran et al. 2009). In our work, we examined the association between HDPR1 and beta-catenin. Our results revealed that enhanced HDPR1 expression was positively correlated with increased beta-catenin expression in the cytoplasm (p = 0.027, r = 0.239; Table 4 and Supplemental Figure S3). Cytoplasmic beta-catenin could transport into the nucleus to activate downstream oncogenes, leading to carcinogenesis (Ilyas 2005; Nelson and Nusse 2004). However, we found that beta-catenin accumulation was observed in the cytoplasm but not in the nucleus, suggesting that new mechanisms mediated by HDPR1 might exist by which tumor malignant progression is regulated in ESCC. Beta-catenin has been reported not only as an activator for gene transcription but also as an important component for the cytoskeleton (Ilyas 2005; Nelson and Nusse 2004). Upregulation of beta-catenin disrupts the organization of the cellular microtubule array because of the interaction with dynein (Ligon et al. 2001). Moreover, interaction of beta-catenin with actin-bundling protein fascin or ezrin, a member of the ERM family, is associated with cell motility (Tao et al. 1996; Hu et al. 2000; Hiscox and Jiang 1999). Our previous works demonstrated that both fascin and ezrin were involved in ESCC invasion; in addition, fascin had an effect on the expression of beta-catenin (Xie et al. 2005; Xie et al. 2009; Xie et al. 2010). According to the results above, it was hypothesized that cytoplasmic HDPR1 could stabilize beta-catenin, resulting in its accumulation in the cytoplasm, which might enhance the interaction with other cytoskeleton-related proteins, leading to reorganization of the cytoskeleton and tumor metastasis. The cytoplasmic beta-catenin functions as a mediator of tumor malignant progression, which works in tandem with several regulatory proteins and downstream genes, thus negating correlation with clinicopathologic factors (Supplemental Table S1). Because there was no staining of beta-catenin in the nucleus, it was proposed that nuclear HDPR1 might play a role by affecting other factors but not beta-catenin in ESCC, such as HDAC1 (Gao et al. 2008). However, more work needs to be done to figure out the distinct involvement of HDPR1 in ESCC.
Relationship between HDPR1 and Beta-Catenin in Esophageal Squamous Cell Carcinoma
As long as the sample contained cells with HDPR1 cytoplasm staining, it was regarded as positive if scored >4 and negative if scored ≤4.
p value <0.05 was considered significant.
In summary, we have shown that cytoplasmic HDPR1 expression is upregulated and associated with tumor metastasis and development via beta-catenin accumulation. There is some connection between cytoplasmic and nuclear HDPR1, which might contribute to tumor malignant progression. Cytoplasmic HDPR1 could be a predictive factor for regional lymph node metastasis and tumor development in ESCC.
Footnotes
The author(s) declared no potential conflicts of interest with respect to the authorship and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: This work was supported by grants from the National Basic Research Program (973 Program No. 2010CB912802), the National High Technology Research and Development Program of China (No. 2006AA02A403), and the Natural Science Foundation of China-Guangdong Joint Fund (No. U0932001).