
Abstract
The present study describes the pathogenetic mechanisms of myocarditis in 9 lambs that died in a foot-and-mouth disease outbreak in Samsun, Turkey. In all the heart samples tested, ELISA and sequencing for phylogenetic analyses showed that the virus, namely O/TUR/Samsun/05, was associated with the PanAsia pandemic strain of foot-and-mouth disease virus (FMDV) type O. The lambs had myocardial lesions but no typical vesicular lesions. In situ reverse transcription showed that many cardiomyocytes and some interstitial cells were positive for FMDV type O. Inflammatory infiltration, hyaline degeneration, and necrosis of sheets of myocytes were observed. The cellular infiltrates were mononuclear cells, including many lymphocytes, macrophages, a few plasma cells, and neutrophils. Major histocompatibility complex Class II+ dendritic and mononuclear cells, γδ T cells, CD172A+ and CD14+ macrophages and monocytes, and IgM+ B cells were detected mainly in the infected hearts. Inducible nitric oxide synthetase (iNOS) was seen mostly in areas of inflammation infiltrated by large numbers of cells. Of the 2 α-subunits of integrin known to be used as receptors by FMDV in epithelial tissues, CD49e (integrin α5) was detected in the membranes of cardiac myocytes with intercalated discs, but CD51 (integrin αV) was not detected in cardiac myocytes from infected or normal lambs. Interstitial and inflammatory cells were positive for both integrin subunits. The terminal deoxynucleotidyl transferase mediated dUTP nick-end labeling (TUNEL)-positive signal was detected in the nuclei of both cardiac myocytes and interstitial cells from infected lambs. These findings suggest that the iNOS expressed by inflammatory cells in lesions may have a deleterious effect on cardiac myocytes in these lesions.
Keywords
Foot-and-mouth disease (FMD) is a highly contagious, severe, clinically acute, vesicular disease of the members of the order Arteriodactyla, that is, cloven-hoofed animals, including domesticated ruminants and pigs, and more than 70 wildlife species. FMD is a problem of worldwide concern, as it is enzootic in large areas of Africa, Asia, Europe, and South America (except the Guyanas and Chile). Foot-and-mouth disease virus (FMDV) is classified within the Aphthovirus genus of the Picornaviridae family. The 7 principal antigenic serotypes are the classical A, O, and C types, SAT-1, SAT-2, SAT-3, and Asia-1. 4
FMD is characterized by the formation of vesicles in the mouth and on the feet, teats, and mammary glands. In general, sheep and goats are less susceptible than other ruminants, and the clinical signs are often mild. Lesions are often seen on dental pads and the dorsum of the tongue, but they are also seen on the lips, gums, and cheeks, and sometimes on the hard palate. 2, 4 The disease is not notable for high mortality except in suckling animals; however, morbidity is very high. 4 The mortality in young animals, in particular lambs and piglets, may be due to acute myocarditis. Myocarditis is considered a fatal form of FMD that occurs without vesiculation in young animals. 2, 4 The residual lesions in older cattle qualify as myocarditis with a significant inflammatory response. 39 However, the significance of acute myocarditis in the spread of FMDV has not been studied in detail. 2 The virus can usually be isolated from myocardia in areas containing necrotic myocytes that are infiltrated with mononuclear cells. 13
FMDV binds to cells through a conserved Arg-Gly-Asp (RGD) sequence located in a flexible external loop between the βG and βH strands (G-H loop) of viral protein 1 (VP1). 18, 32 The RGD is a recognition sequence for members of the integrin family of cell surface receptors. 42 FMDV uses 4 members of the αV subgroup of integrins (αVβ1, αVβ3, αVβ6, and αVβ8) 14– 16, 24– 26, 33, 34 and 1 member of the α5 subgroup (α5β1) 23 as receptors in vitro and in vivo. However, it is unknown which integrins are used for the infection of cardiomyocytes.
The present report describes the pathogenetic features of lesions found in lambs that died of acute myocarditis in an FMD outbreak. For this purpose, we carried out the immunohistochemical characterization of infiltrating inflammatory cells; the expression of inducible nitric oxide synthase (iNOS); and the expression of the α5 (CD49e) and αV (CD51) integrin subunits in cardiomyocytes, which are receptors known to be used by FMDV in the epithelial tissues.
Materials and Methods
Lambs and tissue processing
Tissues were obtained at necropsy from 9 native Karayaka lambs (Nos. 1 through 9). They were between 1 and 2 weeks old and had died during an outbreak of FMD in Samsun Province in northern Turkey. Myocardial tissue specimens were collected from the interventricular, atrial, and ventricular parts of hearts and then fixed in buffered formalin. Tissues were embedded in paraffin, sectioned at 5 μm, and stained with hematoxylin and eosin (HE). Additional paraffin sections were used for in situ reverse transcription (RT) to detect FMDV mRNA. Tissues were frozen for immunohistochemistry (IHC). Cryostat sections were cut at 5 μm, mounted on organosilane (3-aminopropyl) triethoxysilane-coated glass slides, and immediately fixed for 8 minutes in ice-cold absolute ethanol. Additional unfixed myocardial specimens from the infected lambs were used for virus isolation and gene sequence analysis.
The myocardial tissues were obtained from 4 apparently healthy Karayaka lambs, approximately 1 to 2 weeks old, from a local sheep flock; they were euthanatized under deep anesthesia in accordance with specifications of Ondokuzmayis University's Animal Ethic Committee and used as controls. In addition, 2 fetal hearts at approximately 4 months of gestation, and heart, skeletal muscle, skin, cheek, tongue, and lymph node samples from 3 adult 3- to 3.5-year-old sheep were obtained from a local slaughterhouse in Samsun province. The fetal hearts and adult tissues were only used to test CD49e and CD51 immunoreactivities and compare changes in age-associated integrin expression with control and infected lambs.
Virus isolation and identification
The virus was propagated in a monolayer baby hamster kidney (BHK)-21 cell culture from an inoculum prepared from the myocardia of FMDV-infected lambs. Serotype identification of the field virus strain was made by the indirect sandwich enzyme-linked immunosorbent assay (ELISA) method. 17, 40 Microtiter plates (NUNC A/S, Denmark) were coated with specific rabbit hyperimmune sera raised against the FMDV types (A, O, Asia 1) and kept at 4°C overnight. The following day, the plates were washed with phosphate-buffered saline (PBS) and dried, and cell culture samples were added to each well. The plates were then incubated at 37°C for 1 hour. After blocking with 10% bovine serum (containing 5% rabbit serum and 0.1% Tween 20) in PBS, 50 μl of 1/1000 diluted anti-FMDV specific polyclonal guinea pig sera were added to each well. The plates were incubated at 37°C for an additional hour. After washing with PBS, 50 μl of diluted horseradish peroxidase–conjugated rabbit anti–guinea pig immunoglobulin G (IgG) (Dako, Denmark) was added to each well. The color was developed with orthophenylene diamine chromogen (OPD) (Sigma-Aldrich, USA). The optical density (OD) of each well was measured at 492 nm with a microplate reader (Molecular Device, USA). The mean OD value of at least 3 wells showing a value up to 0.1 was considered positive for type-specific FMDV. Negative and positive (with both weak and strong reaction) controls were used for each virus type.
Partial 1D gene sequence analysis
The sequence analysis method used in this study has been previously described. 29 Viral RNA was extracted using TRItidy-G reagent (Applichem Inc., Germany) from cell culture supernatant (BHK-21), according to the manufacturer's instructions.
For cDNA synthesis, a mixture of 25 pmol natural killer (NK) 61 primer (5′ GAC ATG TCC TCC TGC ATC TG 3′), 1 μg RNA, 50 mM Tris-HCl (pH = 8.3), 3 mM MgCl2, 75 mM KCl, 0.5 mM deoxynucleotide triphosphates (dNTPs), 4U ribonuclease inhibitor (Fermentas Inc., Germany), 10 mM dithiothreitol, and 0.5 μl 100U Moloney Murine Leukemia Virus (M-MLV) reverse transcriptase (Fermentas Inc., Germany) was prepared. Twenty-five microliters of this mixture was incubated in a thermal cycler (Eppendorf, Germany) at 42°C for 60 minutes and subsequently at 70°C for 10 minutes.
The polymerase chain reaction (PCR) mixture was composed of 3 μl 25 mM MgCl2, 1 μl 10 mM dNTPs, 5 μl 10× buffer, 1 μl 25 pmol reverse primer NK61 (5′ GAC ATG TCC TCC TGC ATC TG 3′), 1 μl 25 pmol forward primer ARS4 (5′ ACC AAC CTC CTT GAT GTG GCT 3′), 0.5 μl Taq DNA polymerase (5 U/μl), and 5 μl RT product (cDNA). This mixture was bulked to 50 μl DEPC-H2O. Amplification of the PCR was achieved by 1 cycle of 4 minutes at 94°C; 30 cycles of 1 minute at 94°C, 55 °C, and 72 °C, and finally by 1 cycle of 5 minutes at 72°C. Identification of PCR products was done in 2% agarose gel (including 0.5 μl/ml ethidium bromide) at 100V. The PCR products (1,031 bases) were extracted and then purified by removing dNTP, oligonucleotides, and various enzymes, using the NucleoSpin Extract kit, according to the instructions of the manufacturer (Macherey-Nagel GmbH&Co, KG, Germany). The NK72 (5′ GAA GGG CCC AGG GTT GGA CTC 3′) primer was used for the nucleotide sequence reaction, using the Silver Sequencing System kit, according to the manufacturer's instructions (Promega Corp, USA). One hundred and sixty five bases were read at the 3′ end of the 1D gene encoding the VP1 capsid protein. Sequences from the various viruses (Table 1) were aligned and analyzed using ClustalX software 43 and GeneDoc software. 35
Summary of type O viruses used in the present study.
In situ RT for FMDV mRNA
Formalin-fixed, paraffin-embedded tissues were cut into 4- to 5-μm thick sections, placed on superfrost slides, and then deparaffinized: first in xylene for 15 minutes and then in 100% ethanol for 15 minutes. Next, tissues were washed in DEPC-H2O and digested with proteinase K (5 μl/ml) (Sigma, USA) for 60 minutes at 37°C. After digestion, sections were again washed with both DEPC-H2O and 100% ethanol for 2 minutes, and then air dried. DNAase (TACS-Nuclease, Trevigen, Inc., USA) treatment was carried out overnight at 37°C in a humidified chamber. After washing in DEPC-H2O, cDNA was synthesized with M-MLV RT on the sections. An RT mixture of 2 μl×4,200 mM dNTPs, (Fermentas Inc., Germany), 2 μl of 40 mM 11-biotin-dUTP (Fermentas Inc., Germany), 4 μl of 1× buffer and 3 μl of 25 pmol reverse and forward primers was used in sequence analyses. This mixture was incubated at 37°C for 5 minutes. Then 1 μl M-MLV reverse transcriptase (200 U/μl) (Fermentas Inc., Germany) was added. The final volume of the mixture was brought to 100 μl with DEPC-H2O.
The sections were incubated with the mixture in the thermal cycler at 42°C for 60 minutes and then at 70°C for 10 minutes. For inactivation of the endogenous peroxidase, sections were incubated in 0.3% H2O2/methanol for 60 minutes. FMDV mRNA was visualized by using peroxidase-conjugated mouse anti-biotin monoclonal antibody at room temperature for 1 hour; followed by incubation with diaminobenzidine (DAB) chromogen (DAKO, USA) for 10 minutes. After washing with distilled water, sections were counterstained with hematoxylin. Negative controls were the incubation of the RT mixture: with uninfected tissues, without primers, and with unassociated primers of FMDV-infected heart tissue.
IHC
The monoclonal mouse antibodies used in this study and their specificities were: anti-ovine major histocompatibility complex (MHC) Class I (H58A, MHCl I, diluted 1/50) for nucleated cells, with the exception of neural cells; anti-ovine MHC Class II (TH14B, MHCl II, diluted 1/50) for B cells, activated T cells, and dendritic cells; anti-ovine CD1 (TH97A, diluted 1/25) for thymocytes and a subset of dendritic cells; anti-ovine CD5 (ST1, diluted 1/25) for all T cells and a subset of B cells; anti-ovine CD2 (MUC2A, diluted 1/25) for αβ T cells; anti-ovine CD4 (GC1A, diluted 1/25) for T-helper cells; anti-ovine CD8 (CACT80C, diluted 1/25) for T-cytotoxic cells; anti-ovine WC1-N2 (BAQ4A, diluted 1/50) for detecting a major subset of γδ T cells present in ruminants; anti-B cells (BAQ44A, diluted 1/25 and GB26A, diluted 1/25); anti-ovine CD21 (GB25A, diluted 1/25) for mature B cells and follicular dendritic cells; anti-ovine sIgM (PIG45A2, diluted 1/50) for B cells; anti-ovine CD14 (CAM36A, diluted 1/25) for monocytes and macrophages; and anti-ovine CD172A (DH59B, diluted 1/50) for granulocytes, monocytes, macrophages, and a subset of dendritic cells. The monoclonal antibodies (mAb) used in this study were obtained from the Washington State University Monoclonal Antibody Center, Pullman. The polyclonal antibodies used in this study and their specificities included rabbit anti-nitric oxide synthase, inducible (iNOS) Ab-1 antibody (1/100 diluted, Lab Vision Corp, USA), and rabbit anti-integrin α5 (CD49e) and αV (CD51) subunit antibodies (1/1000 diluted, Chemicon International, USA). They were obtained from commercial sources, as shown inTable 2.
Monoclonal(mouse) and polyclonal (rabbit) antibodies used in the present study.
∗ Rabbit whole serum.
Frozen sections were prepared using a commercial streptavidin-biotin peroxidase system (Zymed Laboratories, Inc., USA). To avoid autolysis of cells, the frozen sections were immediately air-dried by using cold air from a hair dryer, and then preserved at −20°C. All sections were preincubated in 10% goat nonimmune serum (Zymed Laboratories, Inc., USA) at room temperature for 10 minutes to block nonspecific binding of second-step antibody (Zymed Laboratories, Inc., USA). Sections were reacted with primary antibodies overnight at 4°C and then rinsed with PBS of pH 7.4 at room temperature. Following that, the sections were reacted with biotin-conjugated second step antibody (Zymed Laboratories, Inc., USA) for 10 minutes at room temperature, and then rinsed in PBS. To inactivate the endogenous peroxidase, sections were incubated in 0.3% H2O2/methanol for 60 minutes. The sections were again rinsed with PBS and reacted with the streptavidin-biotin-peroxidase complex (SABC) (Zymed Laboratories, Inc., USA) for 10 minutes at room temperature. After another washing with PBS, sections were incubated with 3-amino-9-ethylcarbazole (AEC) (Zymed Laboratories, Inc., USA) or DAB (DAKO, USA) for 15 minutes and then counterstained with Mayer's hematoxylin. Four negative controls were prepared, first by omitting the primary antibodies, and then by replacing them with PBS, normal rabbit serum, and unrelated mouse monoclonal antibodies.
Detecting Apoptosis
To detect DNA fragmentation, formalin-fixed, paraffin-embedded sections of myocardial sections were stained by the terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick-end labeling (TUNEL) method (in situ cell death detection, POD® kit, Roche Diagnostics, GmbH, Germany), according to the manufacturer's instructions. Paraffin-embedded sections were dewaxed, rehydrated and irradiated at 350W in 0.1 M citrate buffer of pH 6.0 in a microwave oven for 5 minutes. After washing twice in PBS, sections were covered with 50 μl of the TUNEL reaction mixture containing terminal deoxynucleotidyl transferase (TdT) and fluorescein–dUTP (2′-deoxyuridine 5′-triphosphate), and then incubated under a coverslip in a humidified chamber for 1 hour at 37°C. The sections were checked by fluorescence microscopy. The reaction was stopped by washing slides in PBS. After that, slides were incubated with anti-fluorescein-POD conjugate diluted 1:3 in 100 mM Tris-HCl, 150 mM NaCl (pH 7.5), and 1% blocking reagent for 40 minutes at room temperature. After washing 3 times for 15 minutes in PBS, the sections were stained by incubation with the chromogenic substrate DAB for 5 to 15 minutes at room temperature. Sections from uninfected lambs served as controls for this procedure.
Results
Pathological findings
The 9 dead lambs subjected to examination had no clinical signs. However, all 9 lambs had myocardial lesions. The myocardial lesions had grayish or yellowish foci with irregular patterns ranging in size from pinpoint to 2 cm in diameter. These were seen mainly in the interventricular septum, but sometimes in the right ventricular free wall and in the papillary muscles (Fig. 1). No typical vesicular lesions were found in the mouths or on the feet. In some lambs, similar lesions were also seen on their epicardia. In the flock, however, some adult animals exhibited clinical signs of typical FMD, characterized by oral lesions, salivation, and mild lameness.
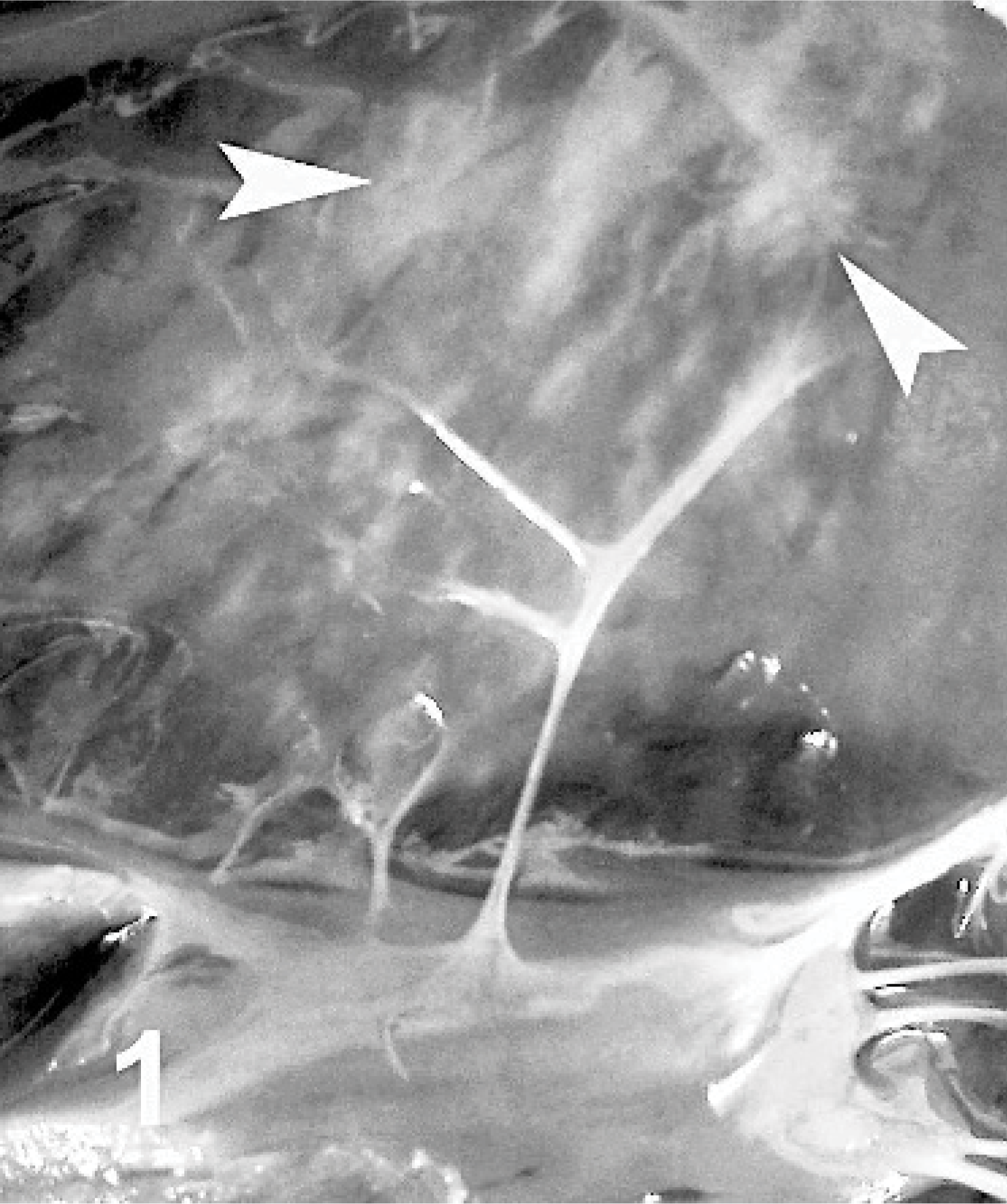
Heart; lamb No. 1. Pale foci (arrowheads) of varying size on the interventricular wall and papillary muscles, with striping and mottled appearance.
Microscopic examination of heart tissues revealed inflammatory infiltration, hyaline degeneration, necrosis of sheets of myocytes with pyknotic nuclei, and occasionally fragmentation of myocytes. Myocardial degeneration to necrosis varied from random and multifocal to confluent in all lambs infected with FMDV. Cellular infiltration and hyaline degeneration did not show any predilection for specific sites in the myocardium. However, endocardial and epicardial sites were more affected in some lambs. Cellular infiltration was composed of mononuclear cells: mainly lymphocytes; macrophages; and, to a lesser extent, plasma cells. In addition, many neutrophils were seen at the periphery of the areas of necrosis and degeneration (Fig. 2).
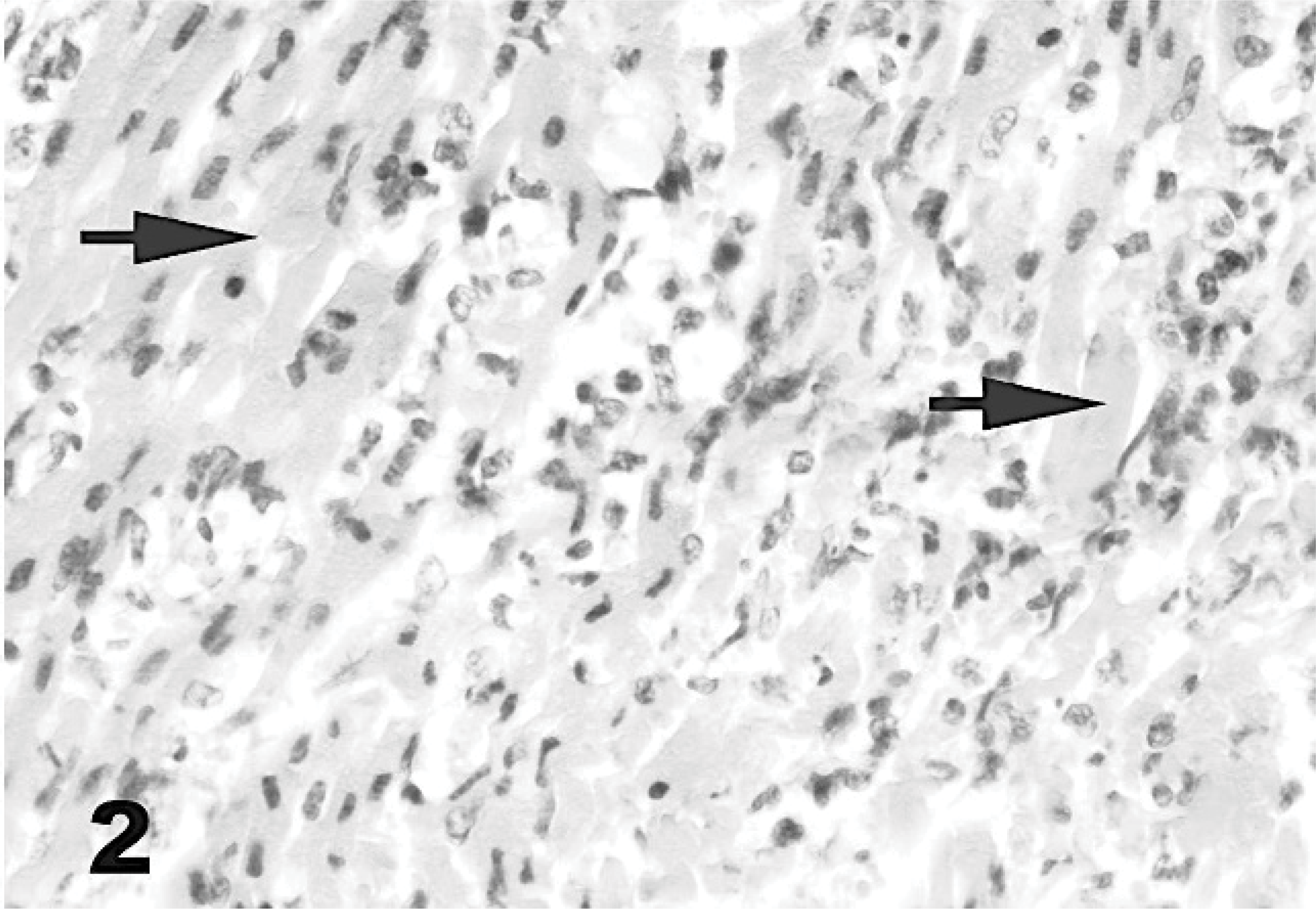
Heart; lamb No. 1. Microscopic appearance of myocardium. Many infiltrating inflammatory cells among the necrotic and/or fragmented cardiac myocytes (arrows). HE.
Virus isolation, identification and 1D gene sequence analysis
All heart samples tested were positive for FMDV type O, as determined by ELISA. Sequencing showed that the virus type isolated in the current study and named O/TUR/Samsun/05 (GenBank accession no. DQ327714) was associated with the PanAsia strain. The isolate was considered closely related to the other type O FMDV, as shown inFig. 3. All controls were negative.

Sequence alignment of 165 nucleotides at the C-terminal of the 1D gene encoding the VP1 capsid protein of viruses used in the present study. Dots indicate nucleotide sites that are identical to that of the sequence of the virus, namely O/TUR/Samsun/05, isolated in the present study.
In situ RT for FMDV mRNA
In situ RT for FMDV mRNA revealed the presence of FMDV in cardiomyocytes (Fig. 4). The intensity of the signal in situ in RT-positive cells increased with the degree of inflammation and myocardial damage seen in the lesions. No FMDV mRNA was detected in the negative controls.
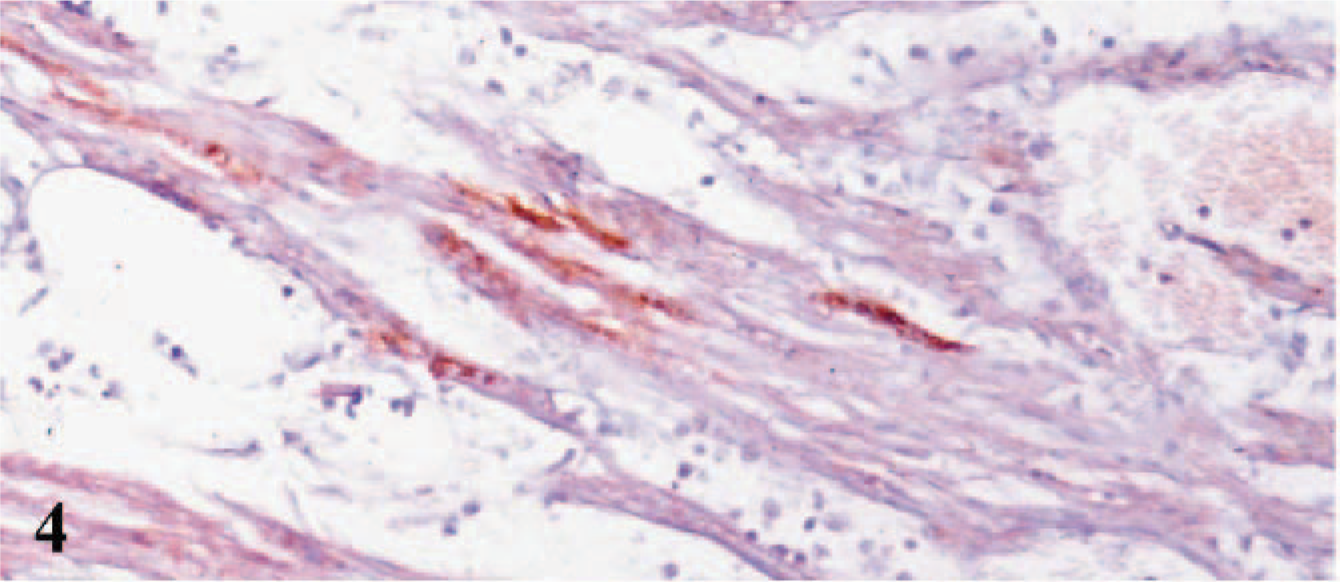
Heart; lamb No. 1. Presence of FMDV mRNA in the cardiomyocytes by combined in situ RT using DAB chromogen and hematoxylin counterstain.
IHC
MHC Class I labeling was localized to the cell membranes of cardiac myocytes, the endothelial cells, and endocardial and epicardial cells of both infected and control hearts. There was also MHC Class I labeling of inflammatory cells in the hearts with myocarditis. In the infected myocardial tissues, MHC Class I expression was stronger than staining noted in the controls.
Some MHC class II+ interstitial dendritic cells were present in the interstitial tissues of hearts in control lambs, especially in the subendocardial, subepicardial, and perivascular areas. The interstitial dendritic cells increased in number, size, distribution, and staining intensity in infected hearts compared with control lambs. MHC Class II expression was found mostly in cells with mononuclear morphology, and to a lesser extent in cells with dendritic morphology in CD4+ and CD8+ rich areas, among both the affected and intact cardiac myocytes (Fig. 5). CD1 labeling was not seen in any sections of hearts from control and affected lambs.
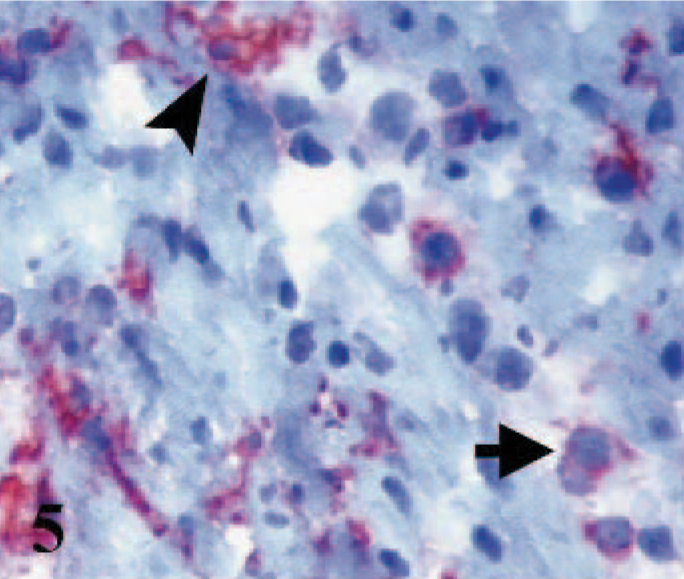
Heart; lamb No. 2. Some MHC Class II expressed dendritic (arrowhead) and mononuclear (arrow) cells. Frozen; SABC/AEC, hematoxylin counterstain.
In the infected myocardial sections, many CD5+ and CD2+ cells were scattered through the interstitial tissues and around the vascular components and subepicardial and subendocardial tissues. In control specimens, these cells were not observed.
Numerous CD4+ and CD8+ positive cells with similar distribution patterns were observed in myocardial tissues infected with FMDV. The frequency of both cell subsets was higher at the periphery of degenerated myocytes, around blood vessels, and in subendocardial and epicardial areas. Some CD8+ cells in the affected myocardium had macrophage morphology.
Many γδ+ T cells (WC1+) were observed in both small and large clusters in areas associated with degenerate to necrotic areas containing other T-cell subsets. In normal heart tissues, few γδ+ T cells were seen in the subendocardial, subepicardial, and perivascular areas.
The number of B cells was less than the number of T cells in the heart tissues of all lambs. Some B cells were scattered among inflammatory cells. CD21 immunostained cells with mononuclear morphology were rare in infected hearts, whereas no CD21+ dendritic cells were observed in any myocardial tissue. Normal cardiac tissues did not reveal B cells when immunostained with CD21 and other B-cell antibodies.
Strong IgM immunostaining was detected among degenerate myocardial elements. Immunostained cells had mononuclear morphology, and there was diffuse interstitial staining of tissues. In some sections, the endothelium of vascular structures had positive labeling in intermediate intensity for sIgM compared with other myocardial elements. The amorphous, eosinophilic material that leaked between the affected cardiomyocytes reacted with the IgM antibody.
Among the myocytes throughout intact and affected areas, CD14 labeling was observed on large cells with pale nuclei and abundant cytoplasm (Fig. 6). Numerous CD14+ cells were dispersed in the perivascular, subendocardial, and epicardial areas and within the clusters of inflammatory infiltrates.
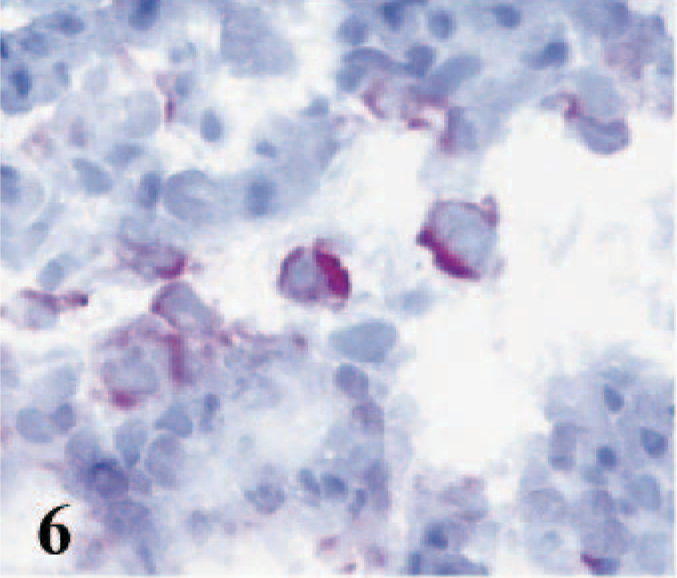
Heart; lamb No. 4. CD14 labeling observed on large cells with pale nuclei and abundant cytoplasm among the myocytes throughout affected areas. Frozen; SABC/AEC, hematoxylin counterstain.
Many CD172A-positive cells were present in the affected areas. Immunostaining was detected in dendritic cells and macrophages and, to a lesser extent, in cells with polymorphonuclear morphology. In severely necrotic areas, immunostaining was more widespread (Fig. 7).
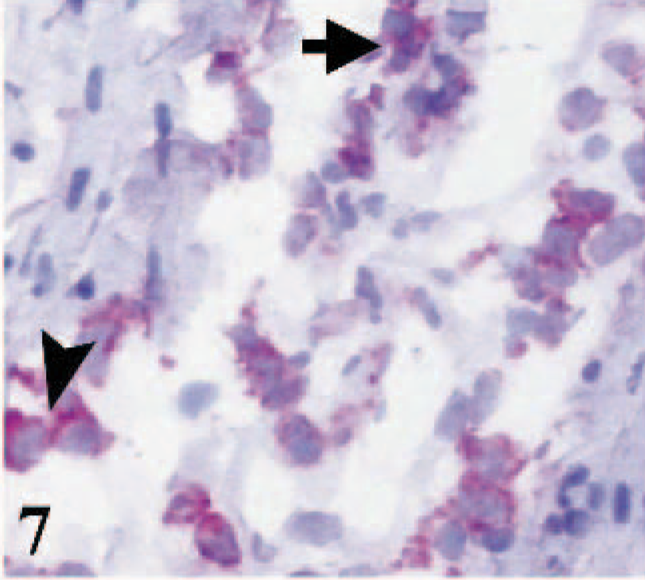
Heart; lamb No. 4. CD172A immunostaining in cells with mononuclear (arrowhead) and polymorphonuclear (arrow) morphology. Frozen; SABC/AEC, hematoxylin counterstain.
iNOS immunostaining was seen mostly in areas that included large numbers of inflammatory cells. Labeling was localized to the membranes of macrophages with pale cytoplasm and nuclei. Small amounts of iNOS labeling was also observed in cardiac myocytes (Fig. 8).
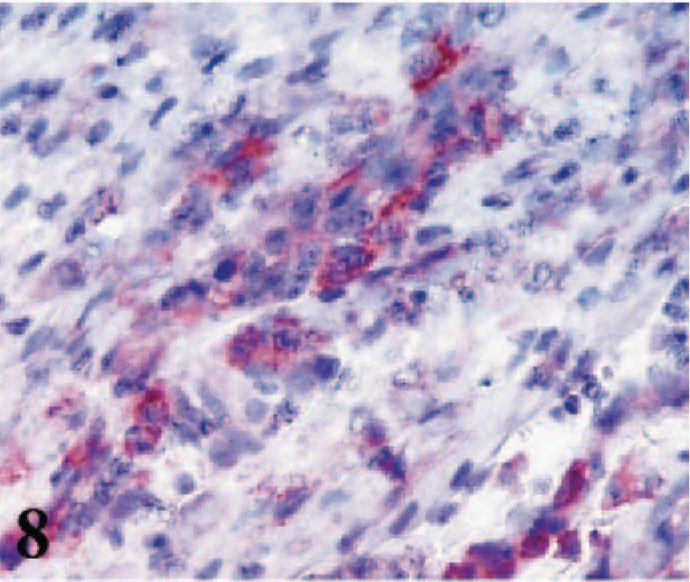
Heart; lamb No. 4. iNOS immunoreactivity in cytoplasm of macrophages infiltrated among degenerated cardiac myocytes. Note weak immunoreactivity in myocytes. Frozen; SABC/AEC, hematoxylin counterstain.
CD49e immunostaining was detected in the membranes and intercalated discs of cardiac myocytes, fibroblasts, macrophages, lymphoid cells, and endothelial cells of the blood vessels of hearts from infected and control lambs, as well as in fetal and adult hearts (Figs. 9– 11).
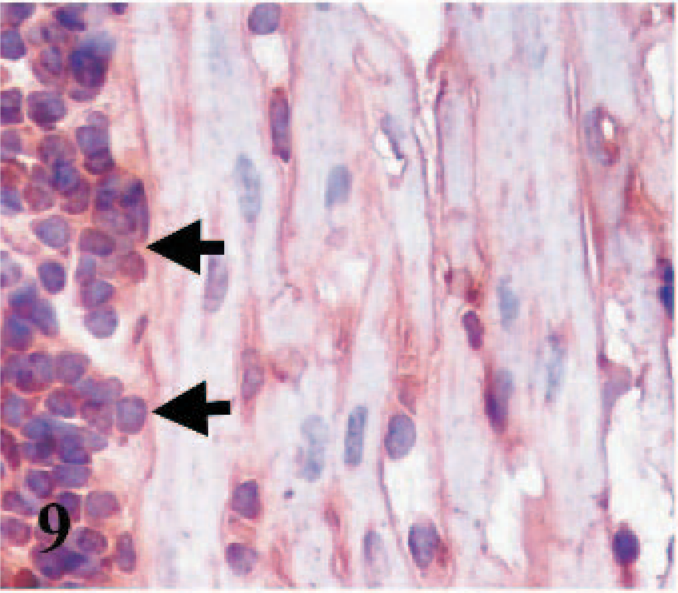
Heart; lamb No. 4. Integrin α5 subunit (CD49e) immunoreactivity in cytoplasmic membranes of cardiomyocytes and inflammatory cells (arrows). Frozen; SABC/DAB, hematoxylin counterstain.
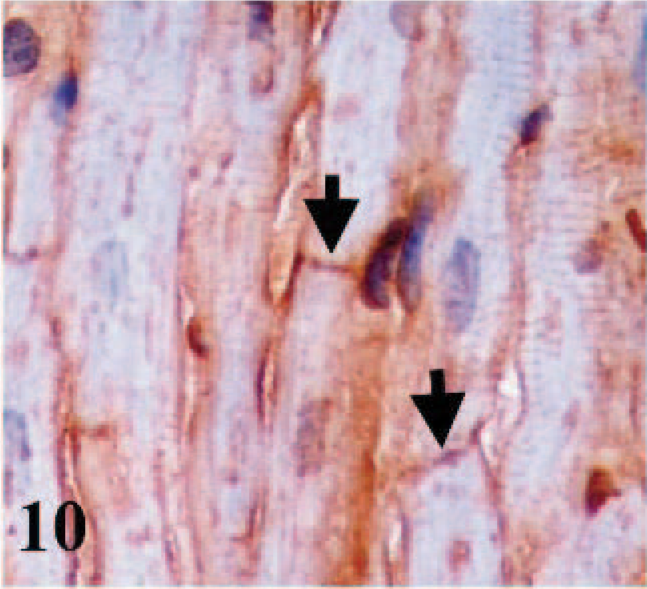
Heart; adult sheep. Immunostaining for integrin α5 subunit (CD49e). Arrows indicate immunostaining of intercalate discs in cardiomyocytes. Frozen; SABC/DAB, hematoxylin counterstain.
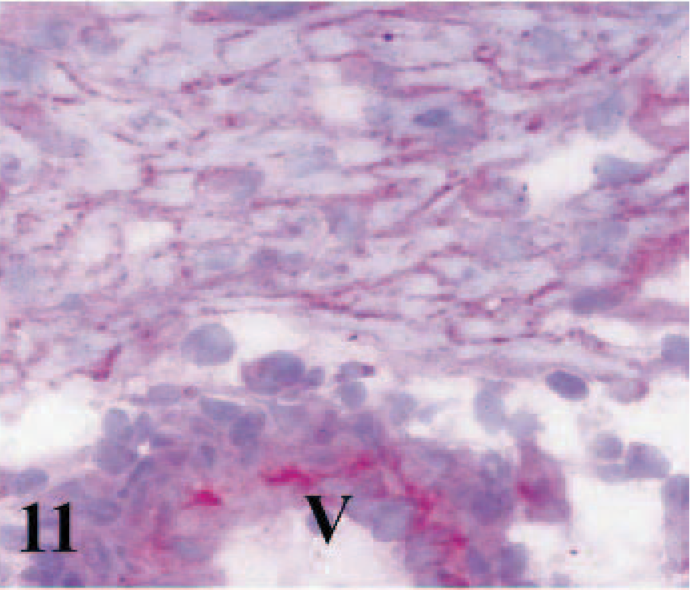
Heart; ovine fetus. Immunostaining of integrin α5 subunit (CD49e) in cytoplasmic membranes of cardiomyocytes and vascular endothelium (V). Frozen; SABC/AEC, hematoxylin counterstain.
CD51 immunostaining was absent from the cardiac myocytes of infected lambs, control lambs, and adult sheep. However, some macrophages, some lymphoid cells, and a few fibroblasts in the interstitial tissues of hearts from all lambs were positive (Figs. 12, 13). Epicardial and endocardial cells and the cardiomyocytes of fetal hearts expressed CD51 antigen (Fig. 14). Weak positive labeling of both CD49e and CD51 was observed in interstitial cells in the skeletal muscles, some adnexal elements of the skin, epithelia of tongues and cheeks, collagen bundles in the subcutaneous tissues, and some sinus macrophages in the lymph nodes from adult sheep.
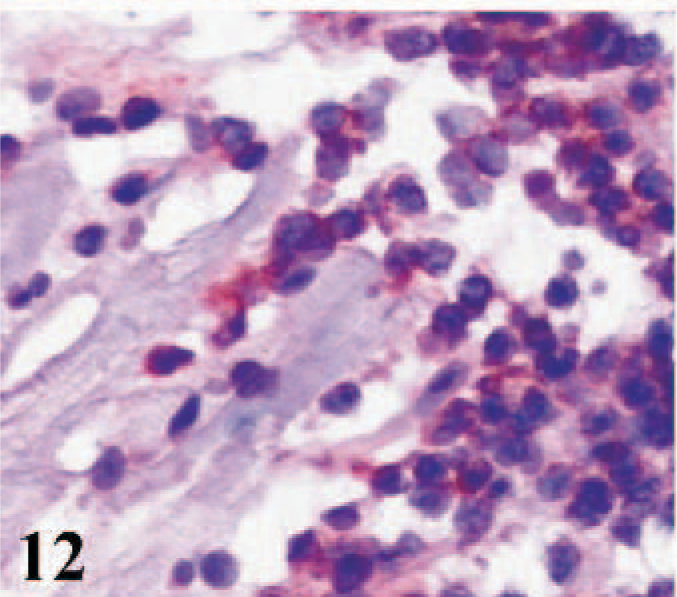
Heart; lamb No. 3. Immunostaining for integrin αV subunit (CD51) in inflammatory and interstitial cells. Frozen; SABC/AEC, hematoxylin counterstain.
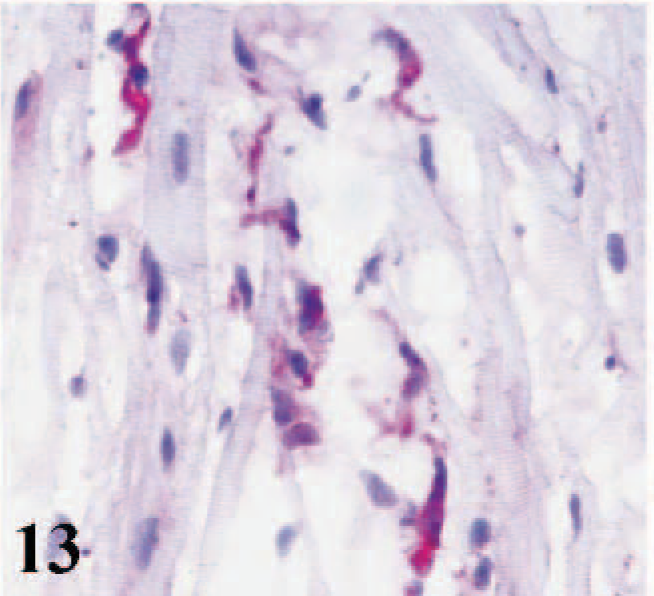
Heart; adult sheep. Immunostaining for integrin αV subunit (CD51) in intersitital cells. Note negative signal of cardiomyocytes. Frozen; SABC/AEC, hematoxylin counterstain.
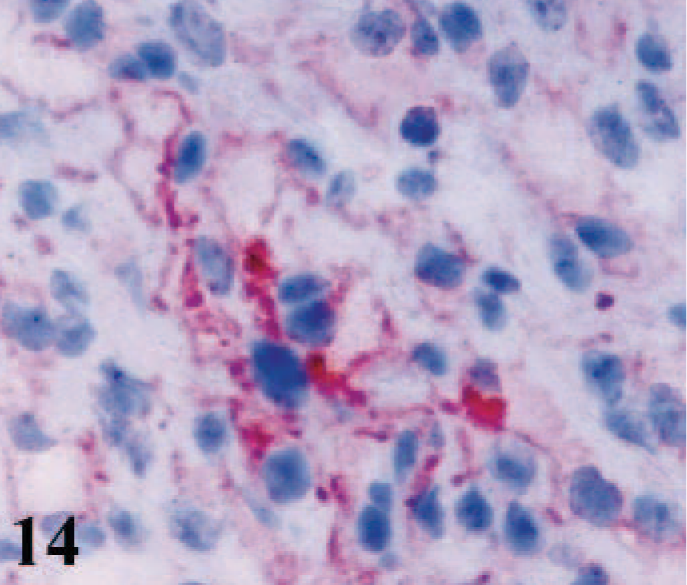
Heart; ovine fetus. Immunoreactivity for integrin αV subunit (CD51) in cytoplasmic membranes of cardomyocytes. Frozen; SABC/AEC, hematoxylin counterstain.
Distribution of apoptotic cells
In myocardia from lambs infected with FMDV, the TUNEL assay indicated that cardiac myocytes, a few endothelial cells, and many interstitial cells (including macrophages, lymphocytes, and fibroblasts) had nuclei that displayed morphologic changes typical of apoptosis. In contrast, the TUNEL-positive signal was not seen in myocardial sections from the control lambs.
Discussion
In the present study, MHC Class II+ dendritic and mononuclear cells, γδ T cells, CD172A+ and CD14+ macrophages and monocytes, and IgM+ B cells, were observed in infected myocardia. Moreover, CD4+ and CD8+ lymphocytes were frequently observed in FMDV-associated lesions in the myocardia of lambs. The role of T cells in immunity to FMDV is still poorly defined compared with that of the humoral response. Phenotypic analysis of lymphocytes from vaccinated and infected cattle showed a small increase in CD8 T cells after infection compared with the frequency observed in vaccinated animals. 19 In the murine model of FMDV infection, B cells appear to play a major role in viral immunity. 5 However, CD8+ T cells are activated in the immune response to FMDV in cattle, 9 as well as CD4+ T cells, which are essential for producing neutralizing antibodies by B cells. CD4+ T cells may also play a role in generating cytotoxic T lymphocyte responses. 9, 10 However, the responses of different organs to FMDV may vary, and the functional interaction of the various cell types in the different organs of sheep remains to be determined.
In the human heart, the number of cardiac dendritic cells (HLA-DR+, CD68+, CD1a negative) increases during the acute phase of myocarditis due mainly to viral infections, and it may exert a destructive effect on the myocytes. 44 In this study, many cells with dendritic morphology were detected in the cardiac tissues of the infected lambs. These results suggest that MHC Class II+ dendritic cells may play an important role in the myocarditis seen in FMDV infection of lambs. Ovine CD1+ dendritic cells in the skin are positive for both MHC Class II and CD21 antibodies, 21 whereas in the present study, MHC Class II+ dendritic cells were negative for both CD1 and CD21 antibodies. Similarly, in the human myocardium, MHC Class II+ cells are negative for CD1a and CD21. 44 In the myocarditis associated with FMDV, myocardial MHC Class II+ dendritic cells may be engaged in viral transport to the myocardium from the primary sites of viral replication, such as the epithelia and lymphoid tissues.
In ruminants, WC1+ γδ T cells are the predominant subset in the peripheral blood, whereas WC1− γδ T cells are more abundant in the spleen. They have a similar abundance in the other secondary lymphoid organs and in the mammary glands, intestine, skin, and uterus. The function of both populations remains to be elucidated. 11 γδ T cells have been purported to play a role in the first line of defense as effector cells, and they may play a unique role in a variety of immune and nonimmune homeostatic processes and during pathological situations. Moreover, damaged tissues, cells, and pathogens can be recognized directly by γδ T cells, without a requirement for antigen processing and presentation. 8 In contrast to other species, the Artiodactyla, including the suborder Ruminantia, contain an extra population of γδ T cells characterized by the expression of the unique molecule WC1 (workshop cluster 1). 1 WC1 is present on the surface of γδ T cells that are CD4−CD8− but express a low level of CD5. WC1+ cells are the most prominent T-cell subset in the skin of sheep at all ages. Although the prominence of γδ T cells in the ruminant immune system has been verified in other tissues, the presence of γδ T cells in the ovine heart has not been previously established. 1, 11, 22 The present study showed γδ T cells in infected heart tissue, and in healthy heart tissue, but at a much lower level. These cells may play an important role in the myocarditis pathogenesis associated with FMDV infection in lambs.
Results from experiments in murine models of myocarditis indicate that, although the immune response to viruses has an important protective role, it may also have deleterious effects on the host. The balance between these protective and deleterious effects may ultimately determine the course of disease after viral infection. 31 In murine myocarditis models, host susceptibility to myocarditis is dependent on the activation of both CD4+ and CD8+ αβ T cells. Both T cells are involved in cardiac tissue damage and mortality from group B coxsackieviruses. 36 There is also significant expression of iNOS, both in infiltrating inflammatory cells 27 and cardiac myocytes 12 Nitric oxide (NO) produced by iNOS has an important role in antimicrobial host defense and immunosurveillance. Low physiologic concentrations of NO can inhibit apoptosis, but higher concentrations may be toxic. High NO concentrations lead to the formation of toxic products like dinitrogen trioxide and peroxynitrite, which induce cell death, if not by apoptosis, then by necrosis. 28 The present study detected iNOS expression in inflammatory cells, mostly in CD14+ and CD172A+ macrophages. This suggested that they were the source of iNOS and, to a lesser extent, the cardiac myocytes undergoing apoptosis and necrosis in the myocardial tissues of FMDV-infected lambs. The occurrence in myocardium of cells with necrotic morphology and apoptotic cells suggests that damage to cardiomyocytes might occur through the release of chemical mediators by inflammatory cells present in the lesions. Further studies are needed to validate this interpretation. The importance of iNOS relative to other factors in FMDV-initiated myocardial injury remains to be determined.
The VP1 of FMDV induces apoptosis of BHK-21 cells via the Akt signaling pathway, after binding to integrins. 37 However, the mechanisms of induction and the role of cardiomyocytic apoptosis in the pathogenesis of FMDV-associated myocarditis remain unclear. Integrins assemble signaling complexes involved in some biological events, including regulation of the cell cycle, proliferation, differentiation, and apoptosis. 20 Previous studies reported that FMDV uses 4 members of the αV subgroup of integrins (αVβ1, αVβ3, αVβ6, and αVβ8) 6, 14– 16, 24– 26, 33, 34 and 1 member of the α5 integrin subunit (α5β1). 23 In the present study, antibodies to α5 and αV integrin subunits were used to determine their immunolocalization in the cardiac myocytes. These integrins are known to serve as receptors for FMDV in vitro and in vivo. The present study showed that α5 is expressed in the cell membrane of cardiac myocytes, as well as in fibroblasts, endothelial cells, and many inflammatory cells. However, αV was expressed only in interstitial cells, including fibroblasts and some inflammatory cells, but not in cardiomyocytes at the level of sensitivity of the immunohistochemical staining. In the heart, age-associated changes in the cardiac matrix and integrins occur. 7 Some integrins are expressed in the embryonic heart and become downregulated postnatally. Expression of α subunits can be temporally modified and developmentally regulated. For example, α1 and α5 integrin subunits are expressed in the embryonic heart, become downregulated postnatally, and can be reinduced by mechanical loading of the heart. αV appears to be uniquely expressed by cardiac fibroblasts, but not by cardiomyocytes. 41 The expression of these integrins in the sheep myocardium remains unknown. This study detected α5 expression in the cardiomyocytes of all animals, whereas the αV subunit was only observed in the cardiomyocytes of fetal hearts. In addition, both integrin subunits were expressed in the interstitial and inflammatory cells. This demonstrated that the α5 integrin subunit does not downregulate with advancing age in the cardiomyocytes of sheep, in contrast to the αV integrin subunit. The current study also detected the expression of 2 integrin subunits and viral mRNA in the cardiomyocytes, as well as in inflammatory cells. Some of these are known to carry infectious FMDV to different sites in the body. 38 However, the present data cannot explain the role of the integrin receptors in the tropism of FMDV to cardiomyocytes and the myocarditis caused by the virus. In older animals infected with FMDV, residual lesions persist after a clinically significant inflammatory response. 39 However, it is difficult to determine whether the residual lesion is the result of infection as a lamb or later in life. Aphthoviruses have flexibility in receptor usage. It is known that different receptors can be used by FMDV and that the potential exists for the virus to use multiple, alternative receptors for entry, even into the same cell type. 3
In conclusion, the present study suggests that the necrosis and apoptosis associated with myocarditis may be attributable to the expression of iNOS and the secretion of cytokines by inflammatory cells present in the myocardial lesions of lambs, thus contributing to the deadly clinical course of FMD in young animals. The current findings should facilitate further studies aimed at elucidating the pathogenetic mechanisms of FMDV-associated myocarditis in greater detail.
Footnotes
Acknowledgements
The authors are particularly grateful to Washington State University's Monoclonal Antibody Center for donating the mAbs for this study and to Gregory T. Sullivan of Ondokuz Mayis University in Samsun, Turkey, for his assistance in editing.