
Abstract
Arrested physical development and neurologic abnormalities were identified in 3 of 5 Rat Terrier puppies at 9 weeks of age. Bilaterally firm symmetrical masses were palpated in the region of the thyroid glands. Low serum total (T4) and free thyroxine (FT4, by equilibrium dialysis) and markedly elevated thyroid stimulating hormone (TSH) concentrations supported the diagnosis of hypothyroidism. At necropsy, the thyroid gland was grossly enlarged and histologically exhibited severe, diffuse hyperplasia of the follicular epithelium. Gross examination of the central nervous system revealed a myelin deficiency, most evident in the corpus callosum. Regional distribution of hypomyelination was confirmed histologically, affecting the corpus callosum and, to a lesser degree, the corona radiata, the longitudinal fibers of the pons, the pyramids, and the lateral funiculi of the spinal cord. Myelin reduction was paralleled by axon reduction, suggesting that hypomyelination was a consequence of reduced axonal formation. A homozygous nonsense mutation in the thyroid peroxidase gene was identified in the affected puppies. The dam and a clinically normal litter mate were heterozygous for this mutation, confirming simple autosomal recessive inheritance of the disease trait. The same mutation, causing congenital hypothyroidism with a goiter was previously described in the Toy Fox Terrier breed. Given the ongoing practice of introducing the Toy Fox Terrier genetic background into some Rat Terrier breeding programs to obtain a smaller stature and the apparent relative incidence of the disorder in the 2 breeds, it is likely that this mutation crossed into the Rat Terrier breed from Toy Fox Terriers fairly recently.
Canine hypothyroidism is generally regarded as an acquired disease of middle aged to older dogs, with only 3.6% of affected dogs less than 1 year of age. 6 Hypothyroidism in young animals results primarily from congenital disorders, which may include those caused by maternal and neonatal exposure to goitrogens, increased or decreased dietary iodine, and inherited defects of the hypothalamo-pituitary-thyroid axis. The incidence of congenital hypothyroidism (CH) in dogs is unknown, although several case reports have been published. 4– 6, 10, 13 Typically, puppies with CH show signs of disproportionate dwarfism, abnormal hair coat (late development of guard hairs), lethargy, and abnormal mentation. Congenital hypothyroidism has also been infrequently reported in cats. 1, 9
Although the clinical signs of neurologic dysfunction in canine CH have been described, 8 descriptions of the central nervous system (CNS) abnormalities are lacking. In humans and in certain laboratory animal species, neonatal thyroid status is known to have a significant influence on the growth, the development, and the maturation of the CNS. 3, 12 The negative effects of thyroid hormone deficiency are most notable during the “critical period” of CNS development that coincides with the onset of active myelination. 12 Neonatal thyroid deficiency primarily affects the cerebrum, with the cerebellum affected to a lesser degree.
In 2003, Fyfe and others identified an autosomal recessive defect of the enzyme, thyroid peroxidase, in Toy Fox Terriers (TFT) suffering from CH with goiter (CHG), and a genetic test was developed to identify carriers of the causative mutation. 5 Because crossing of TFTs with Rat Terriers (RT) is a recognized practice, it appeared possible that the same mutation is segregating in both breeds. The purpose of this study was to investigate the clinical presentation in RT puppies and the CNS lesions in the most severely affected puppy, and to determine whether the TFT mutation is causing CHG in RTs.
Methods and Materials
Animals
Standard physical and neurologic examinations, routine blood cell and serum biologic testing, a quantitative urinary organic acid screen for inborn errors of metabolism, 7 and radiographic evaluation of bony structures were performed. 14 Evaluation of thyroid and pituitary hormones, including total serum thyroxine level (TT4), free T4 (by equilibrium dialysis), and thyroid stimulating hormone (TSH) concentrations were by standard procedures at a national reference laboratory (Antech Diagnostics, Irvine, CA).
Histologic examinations
After euthanasia, tissues (lung, heart, gastrointestinal tract, liver, kidney, pancreas, adrenal, spleen, thyroid, skeletal muscle, peripheral nerve, brain, and spinal cord) were fixed by immersion in 10% buffered formalin, processed through paraffin, sectioned at 5 microns, and stained with hematoxylin and eosin (HE). Additional stains used on selected brain and spinal cord sections were luxol fast blue (LFB, to demonstrate myelin) and Bielschowski (to demonstrate axons). Immunohistochemical stains were performed in the Autostainer plus from Dakocytomation (Carpinteria, CA) by using the streptavidin-biotin immunoperoxidase technique; the antibodies were to glial fibrillary acidic protein (GFAP) (Dako, 1°3000, no pretreatment) and neurofilament (Dako, 1°300, no pretreatment).
Mutational analysis
Buccal brush samples for deoxyribonucleic acid (DNA) isolation were obtained from 2 affected puppies, one clinically normal litter mate, and their dam. DNA preparation, polymerase chain reaction (PCR), and Mwo I digestion were performed as previously described, 5 except that improved PCR primers were used (5′-GCAGCTTCTGTCTTTTTCCAAAC-3′ and 5′-CAACACCCCCACACTCTCTGATG-3′). Gel purified PCR product of one of the CHG affected RTs was sequenced by dideoxy chain termination cycle sequencing at the University of Michigan DNA sequencing core facility.
Results
Animals
A litter of 5 RT puppies was examined at 9 weeks of age. This was the dam's first litter, and whelping was uncomplicated. The owners initially noticed stunted linear growth and lack of weight gain in 3 (1 female and 2 males) of the 5 puppies. The 3 affected puppies had difficulty eating solid food, would often fall over into the food when attempting to eat, and were still on a liquid milk-replacement formula at the time of initial evaluation.
On physical examination, affected puppies appeared disproportionately small when compared with healthy litter mates (Fig. 1). The 3 affected puppies had an average weight of 0.55 kg, whereas the 2 normal puppies had an average weight of 3.8 kg at the time of presentation. Affected puppies had fuzzy hair coats lacking guard hairs and disproportionately large heads, with protruding tongues; they appeared listless and mentally dull, with an uncoordinated, stiff, and mildly dysmetric gait. Bilateral and symmetrical firm subcutaneous masses were palpated in the mid cervical region on both sides of the trachea in all 3 affected pups.

Affected pups and a normal litter mate at 9 weeks of age. Note the small size of the affected pups compared with the normal litter mate.
Serum TT4, FT4, and TSH levels were analyzed in all affected puppies, 1 unaffected litter mate, and the dam (Table 1). In all affected puppies, serum TT4 concentrations were <0.2 μg/dl, well below the reference range of 1.0–4.0 μg/dl. 2 Serum TSH concentrations of affected puppies averaged 3.26 ng/ml, which is 5 times the high end (0.6 ng/ml) of the normal reference range. Values for the normal puppy and the dam were within the reference range (Table 1). Inborn errors of metabolism were not identified on a quantitative urine organic acid screen. Compared with the unaffected puppy, vertebral and limb radiographs revealed delayed epiphyseal maturation (not shown).
Body weights, and thyroid and pituitary hormone concentrations at original evaluation and after thyroid supplementation.
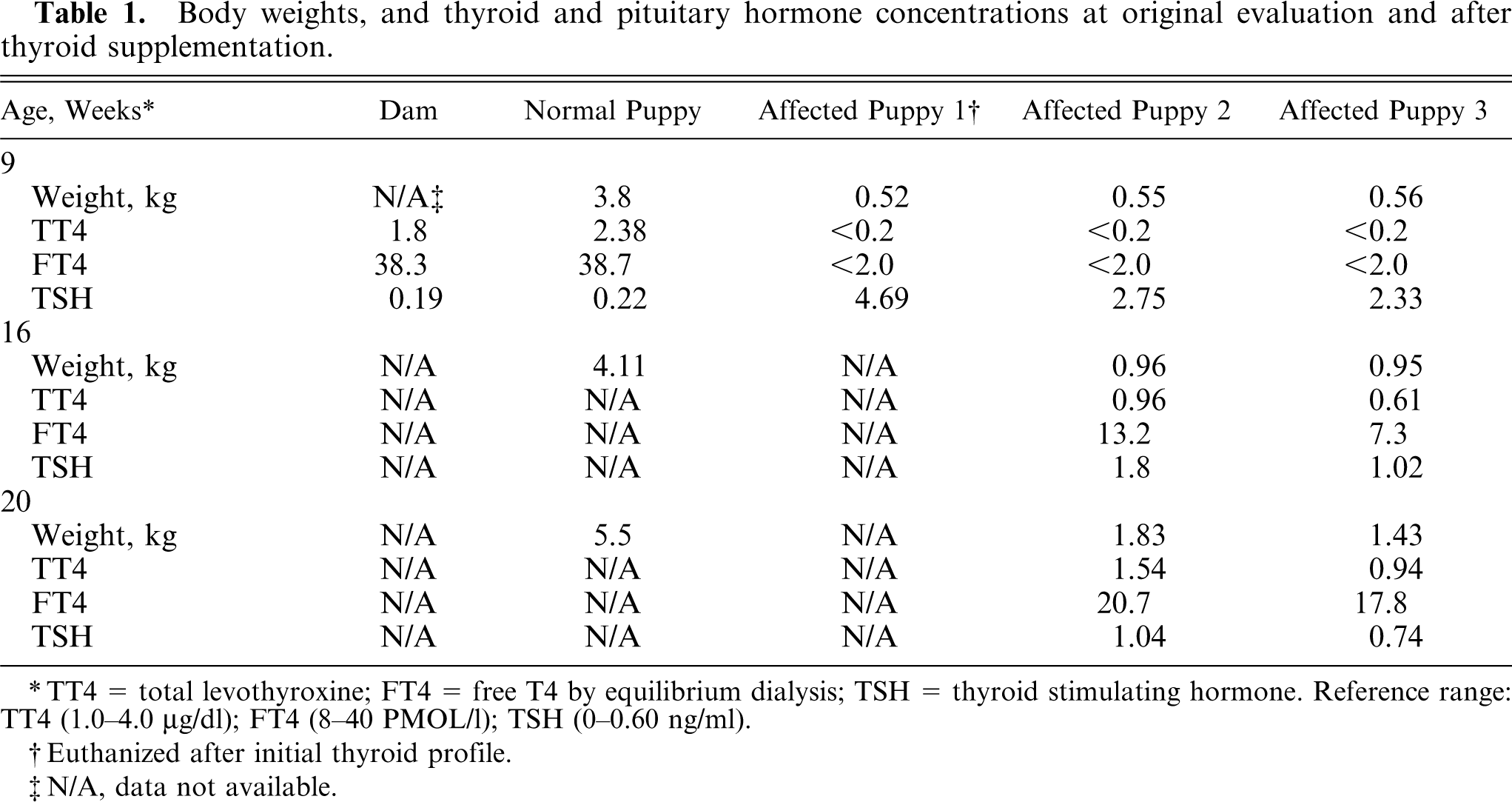
TT4 = total levothyroxine; FT4 = free T4 by equilibrium dialysis; TSH = thyroid stimulating hormone. Reference range: TT4 (1.0–4.0 4;µg/dl); FT4 (8–40 4;PMOL/l); TSH (0–0.60 4;ng/ml).
Euthanized after initial thyroid profile.
N/A, data not available.
Gross and microscopic evaluations
The most severely affected pup was humanely euthanized, and a necropsy was performed. Gross pathologic changes were limited to the thyroid glands, kidney, and brain. The thyroid glands were bilaterally enlarged at 2 × 2 × 1 cm confirming goiter, and the renal cortex was grossly narrow (Fig. 2). In comparison with an age-matched control, transverse sections of the brain revealed that some myelinated tracts were pale and less evident than normal (Fig. 3). Most striking was the corpus callosum, but decreases of white matter were also noted in the periventricular white matter; the terminal corona radiata in the cerebrum; and, to a lesser degree, the cerebellar folial white matter. Other white matter tracts, such as the internal capsule and the crus of the fornix, appeared grossly normal.
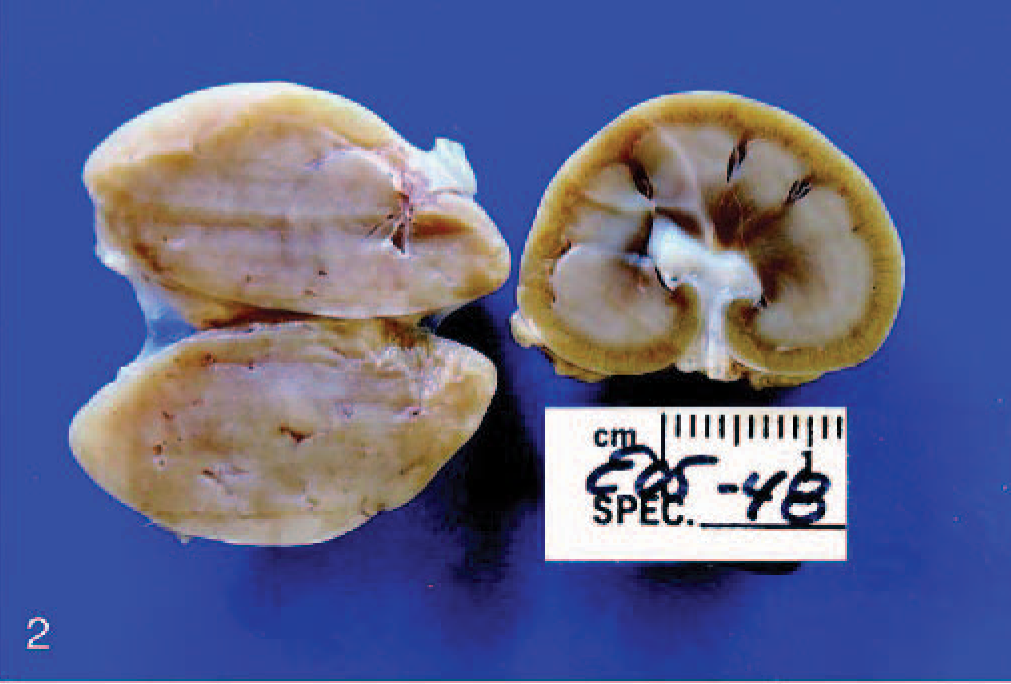
Thyroid and kidney; affected pup euthanized at 9 weeks of age. The peritracheal masses proved to be enlarged thyroid glands (the cut surface is pale tan), which were slightly larger than the kidneys. For a 9-week-old pup, the renal cortex is narrow.
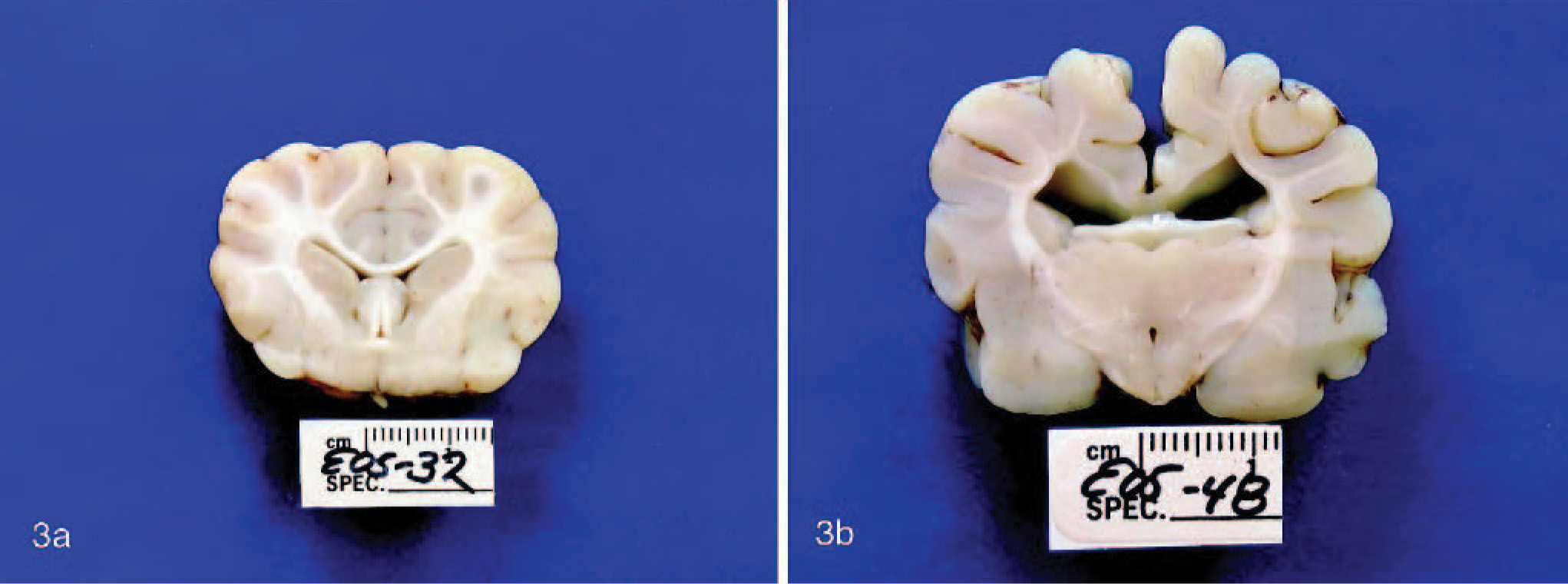
Brain; age-matched control dog (Fig. 3a) and affected rat terrier pup (Fig. 3b). In Fig. 3a, white matter tracts are densely myelinated and well defined. In contrast, in Fig. 3b, the corpus callosum and corona radiata are pale and indistinct because of poor myelination.
No significant microscopic changes were observed in stomach, small intestine, pancreas, adrenal, adipose tissue, spleen, heart, skeletal muscle, peripheral nerve, and spinal ganglion. Hepatocytes had mild cytoplasmic vacuolation, compatible with glycogen storage. The lung was congested, had slightly thickened alveolar septae, and there was amorphous eosinophilic material in alveoli. The renal cortical tubules appeared reduced in number, and glomeruli were seen in unusual proximity to each other, sometimes with 2 side by side (Fig. 4a). The thyroid gland lacked normal colloid-filled follicles and was replaced by hyperplastic follicular epithelium, forming acini lined by cuboidal cells, sometimes several cells thick or thrown into papillae (Fig. 4b, c). Only minimal, faint-staining colloid was found focally, and elsewhere there was none.
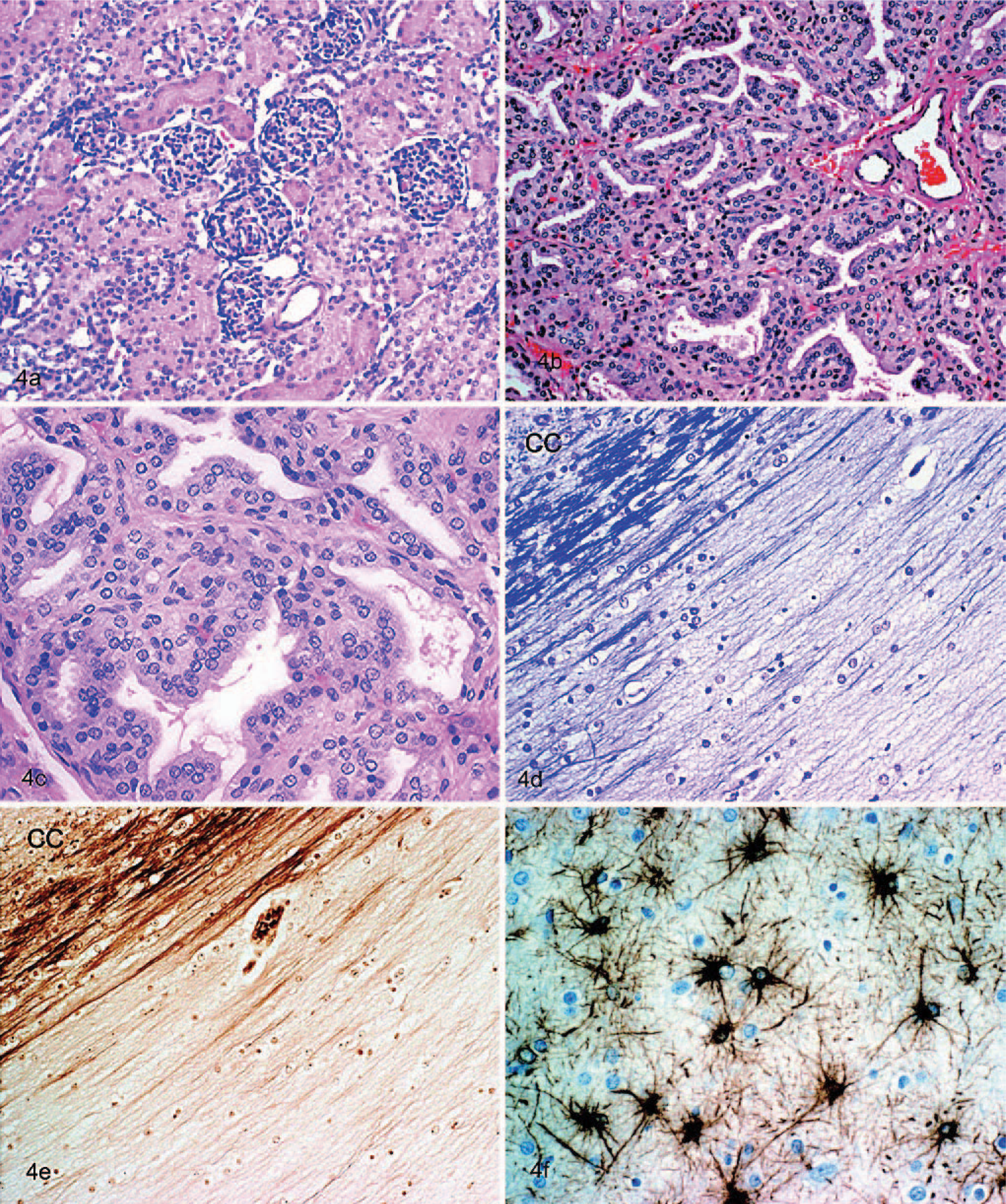
Kidney (Fig. 4a); affected rat terrier pup at 9 weeks of age. The hypoplastic renal cortex revealed unusual proximity of the glomeruli. HE, magnification ×20; Thyroid (Fig. 4b, c). Hyperplastic thyroid follicles were lined by tall cuboidal epithelial cells, which sometimes piled up as blunt papillae. Follicles contained scant strands of colloid or were empty. HE, magnification ×20 for Fig. 4b and ×40 for Fig. 4c.; Brain (Fig. 4d–f). All 3 photos show the corpus callosum and adjacent cerebral cortex (cc). Fig. 4d is a luxol fast blue stain that shows the lack of myelin except for a small bundle of fibers next to the cerebral cortex. Fig. 4e is a Bielschowski silver stain for axons that shows their absence where the myelin is missing in Fig. 4d. Fig. 4f is a GFAP immunohistochemical stain for astrocytes that shows the diffuse development of reactive fibrillary astrocytes. Magnification ×20 for Fig. 4d,e and ×40 for Fig. 4f.
In the CNS, myelin deficiency was most pronounced in the corpus callosum. It was pale with HE stain, and LFB showed it to be almost totally lacking myelin at the midline. In its lateral extensions, the tract contained a few myelinated fibers in the deeper part adjacent to the overlying neocortex (Fig. 4d). Corresponding with the myelin deficit, Bielschowski stain (Fig. 4e) showed a virtual absence of axons, where no myelin was found, and a few bundles of axons, where partial myelin was present. A similar but less marked deficit of myelin and axons was found in the terminal branches of the corona radiata of the cerebrum. Other areas also of mild myelin deficit were in the longitudinal fibers of the pons, the pyramids, and a small area deep in the lateral funiculi of the spinal cord. Generally, the white matter was diffusely gliotic, with prominent open-faced astrocytic nuclei and abundant fine processes on GFAP staining (Fig. 4f). Although the neurofilament staining corresponded with the Bielschowski stain, the latter was more satisfactory for revealing areas of white matter lacking axons.
Mutational analysis
To test the hypothesis that CHG observed in RTs was caused by the same mutation of the thyroid peroxidase gene previously observed to cause the disease in TFT, buccal swabs were obtained for DNA analysis. The C > T mutation of canine thyroid peroxidase that causes CHG in TFTs alters an arginine codon (CGA) to a stop translation codon (TGA), nullifying thyroid peroxidase expression and simultaneously obliterating an Mwo I restriction endonuclease cleavage site. Thus, the 303 bp PCR product amplified from a normal thyroid peroxidase allele is cleaved into fragments of 179 and 124 bp, but the PCR product amplified from a mutant allele is not cleaved. Carriers, clinically normal dogs that are heterozygous for the thyroid peroxidase nonsense mutation, exhibit both the uncleaved PCR product and the Mwo I cleavage fragments. Results of molecular testing for the TFT CHG mutation clearly demonstrated that both of the affected RT puppies that were tested were homozygous for a thyroid peroxidase mutation that abrogated Mwo I cleavage (Fig. 5). Sequencing of the PCR amplification product of one of them demonstrated homozygosity of the previously described C > T nonsense mutation. 5 The dam of the affected puppies and 1 clinically normal litter mate exhibited both cleaved and uncleaved PCR product after Mwo I digestion and are, therefore, CHG carriers. These data also confirmed that CHG is inherited as a simple autosomal recessive trait in RTs, as previously demonstrated in TFTs.
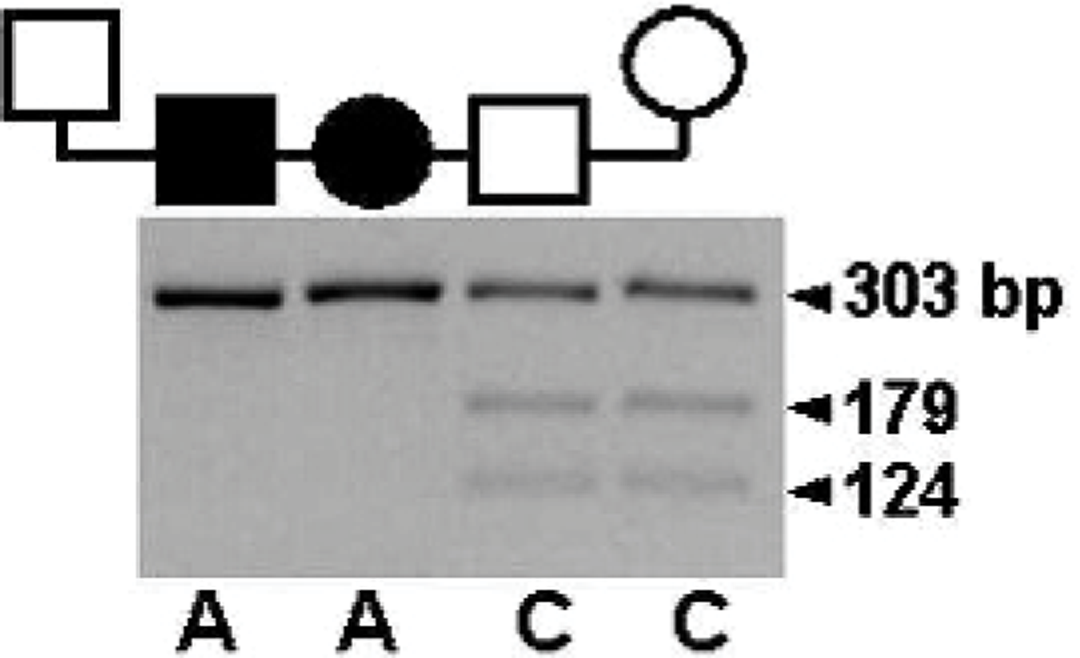
Allele-specific thyroid peroxidase genotyping in a rat terrier family. Males are indicated by squares, females by circles, and filled symbols indicate puppies clinically affected by congenital hypothyroidism with goiter. Genomic DNA flanking the thyroid peroxidase mutation site was amplified by PCR and was subjected to Mwo I digestion. The 303 bp PCR product of the normal allele was cleaved into fragments of 179 and 124 bp, whereas the product of the mutant allele resisted digestion, allowing designation of the tested dogs as genetically normal, carrier (C), or affected (A).
When CHG was confirmed biochemically and histologically, treatment with levothyroxine was initiated at 22 μg/kg PO twice daily. In previous reports, 4, 5 puppies with CHG were severely debilitated, and, after supplementation, skeletal development, hair growth, and increased vigor were noted. After 6 weeks of treatment, the owners reported increased activity and vigor, and the 2 affected RT puppies were reevaluated. Body weights averaged 0.96 kg in the affected puppies compared with 4.11 kg in a normal litter mate. The neurologic examination was essentially unchanged, and the thyroxine dose was increased to 50 μg/kg PO twice daily. Improvement was substantial at 11 weeks after initiation of thyroid supplementation, with an increased activity level and body weights increased dramatically (Table 1). Developing ossification centers and epiphyseal maturation were radiographically visible (not shown).
Concurrent with clinical improvement, the thyroid hormone concentrations increased to within the reference range, with decreased TSH levels by 11 weeks after thyroid supplementation (Table 1). As previously described, the bilateral goiters did not change in size or shape while on thyroid supplementation.
Discussion
Although clinical descriptions of neurologic abnormalities in dogs with CHG are in the literature, there have been no descriptions of CNS abnormalities. Clinical neurologic impairment was evident in the RT pups with CHG and pathologic changes demonstrated histologically. Thyroid hormones are essential for normal neurologic development. By regulating dendrite and axonal growth, synaptogenesis, neuronal migration, and myelination, thyroid hormones are integral in the process of terminal brain differentiation. 3 In studies of hypothyroid rats, retarded development of the neuropil in the cerebral cortex and the cerebellar Purkinje's cells, along with deficiencies in the myelination of the cerebral cortex, visual and auditory cortex, hippocampus and cerebellum were observed. 3 In this litter of CHG puppies, the absence of myelin was most pronounced in the corpus callosum, which serves to associate the 2 cerebral hemispheres. These areas of myelin deficiency appeared to coincide with a concurrent axonal reduction. Thus, both failed axonal and myelin formation may have contributed to the abnormalities found. Much milder involvement of white matter tracts in the pons, the medulla, and the spinal cord may have accounted for the gait deficits. The abnormal sensorium may be explained by the defects in the corona radiate and the corpus callosum.
Because the CHG phenotype in the RT puppies was so similar to that of the previously reported TFT dogs, there was a distinct possibility that the same mutation segregated in both breeds. 5 Molecular testing demonstrated this to be the case. Given the historical and ongoing practice of using TFT breeding stock in some RT breeding programs to obtain a desired smaller stature in their dogs, it is highly likely that the CHG mutation crossed from one breed to the other. This is the first report of CHG in the RT breed. The rather high incidence of CHG in the TFT breed suggests that the mutation is well established and, therefore, flowed from TFT to RT fairly recently. If the mutation predated separation of the populations, one could expect the incidence to be more equal in the 2 breeds. It is possible, however, that a higher incidence of CHG is present in RT but just not recognized or that a popular sire effect in the TFT and the genetic drift in the RT has led to the relatively high and low incidence in the 3 breeds, respectively. Certainly, it is much more likely that the CHG mutation is identical in the 2 breeds by descent from common ancestry rather than each occurring as independent mutations. Nevertheless, in contrast to the TFT breed where the CHG mutation was well spread before the disorder was recognized, the availability of an accurate carrier test should allow RT breeders to rapidly eradicate the carrier state in their dogs. Of utmost importance is that RT breeders use only TFT tested clear of the CHG mutation in future matings.
A broad array of medical conditions has been associated with hypothyroidism of adult onset in the dog, including dermatologic, neuropathic, reproductive, and cardiovascular disorders. 11 However, CH with an inherited basis is uncommon, and typical signs are epiphyseal dysplasia, lethargy or stupor, and failure of haircoat maturation. Examination of the literature indicates that when goiter is present, as in the affected TFT and RT, the signs of CH occur earlier and are more severe than when the goiter is not present. Lesions demonstrated in this report suggest that CHG dogs may be an appropriate model of disturbed neurodevelopment in thyroid hormone deficiency. Retarded growth and disturbed neurodevelopment, together with thyroid enlargement in early life, should alert the clinician to the possibility of thyroid deficiency. This can readily be confirmed with serum analysis, and in the TFT and RT, by demonstration of the disease mutation by DNA-based genetic testing.
Footnotes
Acknowledgements
The authors would like to thank Dr. Duncan Ferguson for helpful discussions, Dr. Ronald Schecter for referral of the dogs, and Dr. Bruce Lindsey for utilization of facilities. This study was supported, in part, by the Muscular Dystrophy Association and the Myasthenia Gravis Foundation of California.