
Abstract
Porcine circovirus type 2 (PCV2) has been confirmed as the primary cause of postweaning multisystemic wasting syndrome (PMWS). However, in the field, PMWS is seen only in a small percentage of pigs infected with PCV2. The overall objective of the study reported here was to determine whether host genetic differences in the susceptibility to PCV2-associated disease exist among selected breeds of pigs. This study included Duroc (n = 23), Landrace (n = 19), and Large White (n = 21) pigs. The pigs were infected intranasally and intramuscularly at 5-7 weeks of age with PCV2. A portion of the pigs (31/63; 30.2%) had low passively acquired PCV2 antibodies at the time of infection. There were no differences in mean weight gain, rectal temperature, or respiratory score. Clinical disease compatible with PMWS was observed only in the Landrace pigs. Most of the PCV2-infected pigs had enlarged lymph nodes, and individual Duroc and Landrace pigs had mottled tan lungs. PCV2-associated lymphoid depletion and granulomatous inflammation were observed in pigs of all breeds. Three of 19 Landrace pigs and none of the Duroc or Large White pigs developed severe lymphoid lesions associated with large amounts of intralesional PCV2 antigen typical of PMWS. Compared with seronegative Landrace pigs, Landrace pigs that had low maternal antibodies at the time of PCV2 inoculation had significantly (P < 0.05) less-severe PCV2-associated lesions. The results suggest a predisposition of the Landrace pigs of this study to PCV2-induced disease and lesions, and that low levels of passively acquired antibodies are protective.
Keywords
Porcine circovirus (PCV) was first detected as a contaminant of a continuous porcine kidney cell line in 1974 70 and was further described in 1982. 69 In the late 1990s, PCV was associated with postweaning multisystemic wasting syndrome (PMWS). 18 Further analysis of the virus documented two distinct types of PCV: nonpathogenic type 1 (PCV1) and pathogenic and PMWS-associated type 2 (PCV2). 6, 46, 50 Subsequently, PCV2 has been associated with several other swine diseases including porcine respiratory disease complex, 31 reproductive disorders characterized by abortions with mummified fetuses, 73 enteritis, 38 and porcine dermatitis and nephropathy syndrome. 63 PMWS was first observed in a Canadian herd in 1991, 28 and is now recognized worldwide. 1 The syndrome is characterized by a clinical history of wasting or poor performance in weaned pigs and by severe lymphoid depletion and histiocytic replacement of follicles in lymphoid tissues. 66 Detection of PCV2 antigen or nucleic acids within characteristic microscopic lesions is required for the diagnosis of PMWS. 66
Serologic surveys indicate high prevalence of PCV2 in the swine population 16, 17, 34, 44, 72 ; however, the prevalence of PCV2-associated disease seems to vary considerably from farm to farm and country to country. Morbidity attributable to PMWS reportedly varies from as little as 4% to as high as 30%, and mortality in affected pigs is typically 70–80%. 1, 28, 43, 61 However, most reports to date lack important additional information regarding breed, production style, and environment of the farm, or description of other diseases present in the pigs that may have influenced expression and outcome of PCV2-associated diseases. Genetic analyses to date have not been able to document major differences between PCV2 isolates recovered from affected pigs and those from nonaffected pigs. 12, 42 Analysis and comparison of PCV2 isolates from severely affected countries like Spain or France with those from countries that are less affected like the United States 20, 45 have failed to explain differences in clinical manifestations of PCV2-associated diseases. All isolates sequenced and compared so far indicated nucleic acid sequence homology >94%. 20, 45
Experimental PCV2 infection models indicate that PCV2 is an opportunist, depending on immunostimulation 3, 39 or coinfecting agents like porcine parvovirus (PPV), 7, 36, 40, 52 porcine reproductive and respiratory syndrome virus (PRRSV), 2, 32, 64 or Mycoplasma hyopneumoniae 54 for PCV2 infection to progress to clinical PMWS. Although most research groups were not able to induce clinical disease in pigs inoculated singularly with PCV2, some groups have succeeded in this regard. 8, 32 PMWS was recently induced in conventional colostrum-fed pigs singularly infected with PCV2. 41 The exact reasons for the different outcomes in experimental PCV2 infection models are not known, but there are several possible explanations including differences in environmental conditions and stressors, age of the pigs at infection, or differences in type and dose and routes of the PCV2 inoculation.
One other aspect that, to the authors' knowledge, has not been investigated is the influence of host genetics on PCV2 infection. Differences in genetic susceptibility of populations or breeds to parasites, bacteria, and viruses have been reported. 9, 10, 30, 68 Evidence exists for pig breed-dependent differences in disease resistance for a variety of pathogens including pseudorabies virus, 59 PRRSV, 27 Escherichia coli, 15, 49 and Sarcocystis miescheriana. 60 Pigs of all breeds seem to be susceptible to PCV2 infection, and PMWS has been observed within a large variety of purebred and crossbred pigs submitted to a US diagnostic laboratory (P.G. Halbur, unpublished data). Recently, PMWS has been reported in Eurasian wild boars in Canada 19 and Spain. 71 Retrospective studies indicate that naturally acquired PCV2 infections in pig herds date back for at least 20 years prior to the first observation of PMWS 25, 44, 65, 72 ; however, PCV2-associated diseases emerged in the late 1990s. Allan et al. 5 reported experimental induction of PMWS in colostrum-deprived Northern Ireland Landrace/Large White crossbred pigs inoculated with a PCV2 isolate recovered in 1993 from a subclinically infected Swedish Yorkshire pig, implying host and/or environmental dependant factors associated with the development of PMWS. A recent in vitro study investigating the replication patterns of PCV2 in pulmonary alveolar macrophages indicated clear differences among macrophages derived from different conventional crossbred pigs that were suggestive of differences in susceptibility to PCV2-associated disease. 47
The overall objective of the study reported here was to determine whether there is evidence for host genetic differences in susceptibility to PCV2-associated disease among selected breeds including Duroc, Landrace, and Large White. These breeds were selected because they could be purchased from a single source.
Materials and Methods
Animal source
Seventy-five purebred (n = 26 Duroc; n = 25 Landrace; and n = 24 Large White), colostrum-fed, female pigs raised under identical conditions on the same farm and in the same barn were purchased from a herd that is routinely tested for major swine pathogens and is free of PRRSV and M. hyopneumoniae. There was no evidence of PCV2-associated disease in the source herd or in the multiplier herds supplied by this source. The source herd is routinely vaccinated against PPV infection and leptospirosis. The pigs were born within a range of 23 days, randomly selected on farm after blocking by sire and litter, weaned at 10–12 days of age, and transported in two groups of 39 (n = 12 Duroc; n = 14 Landrace; and n = 13 Large White) and 36 (n = 14 Duroc; n = 11 Landrace; and n = 11 Large White) pigs 2 weeks apart to the Livestock Infectious Disease Isolation Facility at Iowa State University, Ames, Iowa. The pigs of the various breeds were housed in two rooms until inoculation.
Serologic testing
Blood samples were collected at arrival of the pigs, followed by blood collections at 2-week intervals until the day of PCV2 inoculation. Thereafter, blood was collected on postinoculation day (PID) 7, 14, 21, 28, and 35. Preinoculation sera (from arrival up to PID 0) and serum samples obtained on PID 21 and 35 were tested by a PCV2-enzyme linked immunosorbent assay (ELISA) that is based on the recombinant ORF2 capsid protein of PCV2. 51 Results were considered positive if the calculated sample-to-positive ratio (S ° P) was ≥0.2.
The serum samples obtained on arrival of the pigs at the research facility and the serum samples obtained at necropsy from 11 randomly selected PCV2-infected pigs of each of the three breeds were tested for the presence of antibodies to PRRSV by use of a PRRSV-ELISA (IDEXX Laboratories, Inc. Westbrook, MA), and for the presence of antibodies to PPV by use of a hemagglutination-inhibition assay. 48
Experimental design and PCV2 inoculation
Twelve pigs (three Duroc, six Landrace, and three Large White pigs) were kept as sham-inoculated, control pigs and were housed together in a separate room. These pigs were procedural controls, selected on the basis of presence of maternal antibodies to PCV2. Experimentally induced PCV2 in the remaining 63 pigs (23 Duroc, 19 Landrace, and 21 Large White pigs) was initiated after waning of the PCV2 maternal antibody levels below the negative cutoff (S ° P <0.2). All of the PCV2-infected pigs were housed in three rooms and were randomly distributed into two to seven pens of five to six pigs per pen. The procedural controls were housed separately in a fourth room. To account for and minimize sire and litter effects, a minimum of four sires were represented within each breed and two to five piglets were chosen from each dam. The sire and dam distribution in the PCV2-inoculated pigs was as follows: Duroc pigs (n = 23) represented four sires and eight dams, and the Landrace (n = 19) and the Large White (n = 21) pigs represented four sires and six dams, respectively.
PCV2 isolate ISU-40895 20 was obtained via direct transfection of PK-15 cells with an infective clone of PCV2 as described. 21 Passage 4 of the virus at a titer of 104.2 50% tissue culture-infective doses was used for inoculation of the pigs. The inoculum tested negative for the presence of porcine parvovirus and PCV1 nucleic acids on the basis of results of polymerase chain reaction (PCR) analysis. 20, 37 After production, the PCV2 inoculum was stored in individual 50-ml tubes at −80°C and thawed only once immediately prior to inoculation. To better ensure infection of all pigs at the time of inoculation, each pig received 1 ml of the PCV2 inoculum intranasally and 0.5 ml intramuscularly in the right side of the neck as described. 22, 23 On the day of PCV2 inoculation, the pigs were 5–8 weeks old. Mean ± SE age was 43.5 ± 1.8 days for the Duroc, 48.6 ± 2.0 days for the Landrace, and 46.0 ± 2.0 days for the Large White pigs.
Clinical evaluation
After PCV2 inoculation, pigs were monitored daily and scored for severity of clinical respiratory disease, with scores ranging from 0 to 6 (0 = normal and 6 = severe dyspnea and abdominal breathing). 26 In addition, pigs were evaluated daily for clinical signs of disease including sneezing (score range from 0 [no sneezing] to 3 [severe persistent sneezing]), and jaundice. Rectal temperature, wasting, and behavioral changes such as lethargy were recorded daily. The pigs were weighed on the day of PCV2 inoculation and on PID 7, 14, 21, 28, and 35.
Quantification of PCV2
Extraction of DNA from sera obtained on the day of PCV2 inoculation and on PID 7, 14, 21, 28, and 35 was performed using the QIAamp DNA Mini Kit (Qiagen, Valencia, CA). The DNA extracts were used for quantification of the amount of PCV2 genomic DNA by use of real-time PCR analysis. 55
Necropsy
Necropsy was performed on half of the PCV2-infected pigs of each breed at PID 21 (n = 12 Duroc, n = 11 Landrace, n = 11 Large White) and on the remaining pigs at PID 35 (n = 11 Duroc, n = eight Landrace, n = 10 Large White). In addition, 7 of 12 control pigs (two Duroc, three Landrace, and two Large White pigs) were necropsied on PID 21, and 5 of 12 (one Duroc, three Landrace, and one Large White pig) were necropsied on PID 35. The total amount of macroscopic lung lesions (0–100% of the lung affected), 26 and size of lymph nodes (score range from 0 to 3; 0 [normal], 1 [two times the normal size], 2 [three times the normal size], 3 [four times the normal size]) were estimated.
Lungs were insufflated with fixative as described. 26 Sections of lymph nodes (superficial inguinal, external iliac mediastinal, tracheobronchial, mesenteric), tonsil, thymus, ileum, kidney, colon, spleen, and liver were collected at necropsy and fixed in neutral-buffered 10% formalin and processed in routine manner for histologic examination.
Histologic examination and immunohistochemical analysis
Microscopic lesions were evaluated in a blinded manner. Lung sections were scored for presence and severity of interstitial pneumonia ranging from 0 to 6 (0 = normal; 6 = severe diffuse). 26 Sections of heart, liver, kidney, ileum, and colon were evaluated for the presence of lymphohistiocytic inflammation and were scored from 0 = none to 3 = severe. Lymphoid tissues including lymph nodes, tonsil, and spleen were evaluated for the presence of lymphoid depletion ranging from 0 = normal to 3 = severe histiocytic inflammation and replacement of follicles ranging from 0 = normal to 3 = severe. 54
Immunohistochemical detection of PCV2-specific antigen was performed on formalin-fixed and paraffin-embedded lymphoid tissue sections (lymph nodes [superficial inguinal, external iliac mediastinal, tracheobronchial, and mesenteric], tonsil, spleen, and thymus) of all pigs and sections of intestine from selected pigs using a rabbit polyclonal antiserum. 67 Scoring for PCV2 antigen was done in blinded manner, and scores ranged from 0 = no signal to 3 = >50% of the lymphoid follicles containing cells with PCV2 antigen staining. 54 Mean group score was determined for each tissue and was compared among groups.
The overall PCV2-associated lesions were evaluated as described. 54 A combined scoring system for each lymphoid tissue that ranged from 0 to 9 (lymphoid depletion score 0–3; granulomatous inflammation score 0–3; PCV2-IHC score 0–3) was used. The scores (lesions and PCV2-IHC) for the seven lymphoid tissues ([lymph node pool multiplied by five] + [spleen] + [tonsil]) were added together and divided by seven. The lymph node pool consisted of one section each of tracheobronchial, superficial inguinal, external iliac, mediastinal, and mesenteric lymph nodes. Pigs were grouped into four categories on the basis of overall microscopic lymphoid lesion scores: normal (score = 0), mild (score = 1–3), moderate (score = 4–6) and severe (score = 7–9). The mean treatment group lymphoid score was calculated and compared among groups. A pig was diagnosed as having microscopic lesions typical of PMWS if the mean lymphoid microscopic lesion severity score was severe (score 7–9) and if there was evidence of lymphohistiocytic inflammation in other organ systems.
Statistical analysis
The main outcome of interest in this study was evaluation differences in PCV2 susceptibility among breeds. Therefore, owing to the low number of sham-inoculated control pigs, statistical analyses were limited to PCV2-infected pigs. Summary statistics were calculated for all PCV2-infected pigs within each breed to assess the overall quality of the data. Analysis of variance (ANOVA) was used for cross-sectional assessment of the average daily weight gain and nonrepeated continuous measures. The significance level was P < 0.05 followed by pairwise testing using the Tukey-Kramer adjustment to identify the groups that were different. Effect of sire or dam on the outcome was tested by using the model: y = breeds + sires (breeds) + dams (sires, breeds). To summarize and simplify the clinical observations, response feature analysis and chi-square testing was used. The clinical scores for each pig were reduced to one weekly mean score, and the resulting values were subjected to statistical analysis. Daily rectal temperature data were analyzed with multivariate ANOVA. Nonrepeated measures of necropsy and histopathologic data were assessed using nonparametric Kruskal-Wallis ANOVA. If a nonparametric ANOVA result was significant (P < 0.05), Wilcoxon tests were used to assess the differences of pairs of groups. Differences in incidence were evaluated by using Fisher's exact test.
A portion of the pigs within each breed had low maternal antibodies at the time of PCV2 infection. To evaluate influence of the antibody status on the results regarding PCV2-associated lesions, maternal antibody-negative and maternal antibody-positive pigs were compared separately among breeds, and pairwise Wilcoxon tests on outcomes of antibody-negative (S ° P <0.2) and antibody-positive (S ° P ≥0.2) pigs within each breed were done.
Results
Clinical disease
From PID 7 to 14, PCV2 infection was clinically characterized by mild respiratory distress and, occasionally, by sneezing. Mean respiratory scores were higher in Landrace pigs for PID 7–14, but were not significantly different (P > 0.05) among breeds. Individual pigs of all three breeds had periodically increased temperature; however, there was no difference during the 21 days (P = 0.189) or from PID 21 to PID 35 (P = 0.372) in mean body temperature among breeds nor was there a time-by-group interaction.
From the day of PCV2 inoculation onward to PID 35, mean ± SE weight gain was 633.7 ± 32.8 g for Duroc pigs, 763.8 ± 49.5 g for Landrace pigs, and 746.4 ± 36.8 g for Large White pigs. The weekly weight gain (data not shown) was not significantly different (P > 0.05) among the breeds.
One Landrace pig developed persistent fever >41.1°C from PID 14 onward and had to be euthanized at PID 21, although it had been randomly selected for the PID 35 necropsy group. This pig gained only 6 g between PID 14 and 21, which is consistent with wasting and clinical PMWS in a growing pig. One other Landrace pig had decreased average daily weight gain, compared with that in the other Landrace pigs.
Serologic test results
At arrival, the majority of the pigs (65/75) had maternal antibodies to PCV2, with mean PCV2-ELISA S ° P of 0.46 ± 0.08 for Duroc pigs, 0.57 ± 0.08 for Landrace pigs, and 0.43 ± 0.07 for Large White pigs. In the majority of the pigs (18/23 Duroc, 10/19 Landrace, 16/21 Large White), maternal antibodies waned below the ELISA cutoff (S ° P <0.2) prior to PCV2 inoculation (PID-0 seronegative pigs). At the time of PCV2 inoculation, 5 of 23 Duroc, 9 of 19 Landrace, and 5 of 21 Large White pigs had low maternal antibody levels (PID-0 seropositive pigs) with mean S ° P of 0.22 ± 0.00 for Duroc, 0.23 ± 0.01 for Landrace, and 0.24 ± 0.01 for Large White pigs. As expected, 56 the passively acquired antibodies in the noninfected control pigs decayed over time. None of the control pigs seroconverted to PCV2 during the study. Most of the PCV2-infected pigs seroconverted to PCV2 between PID 21 and PID 35. At PID 35, 7 of 11 Duroc, 6 of 8 Landrace, and 6 of 10 Large White pigs were positive for PCV2-specific antibodies.
All pigs tested negative for PRRSV-specific antibodies. The pigs had maternal antibodies to PPV that decayed over time as expected. 58 Indications of seroconversion to PRRSV or PPV were lacking.
PCV2 viremia duration and genomic copy numbers
All serum samples obtained from the noninfected, control pigs were negative for PCV2-specific nucleic acids, as tested by real-time PCR analysis. In the PCV2-infected pigs, PCV2 viremia was detected first at PID 7 in 13 of 23 Duroc, 14 of 19 Landrace, and in 9 of 21 Large White pigs. Landrace pigs had significantly (P < 0.05) higher incidence of viremic pigs at PID 7, compared with that for the Large White pigs. The majority of the PCV2-infected pigs were viremic by PID 14 (21/23 Duroc, 17/19 Landrace, and 21/21 Large White pigs). The duration of PCV2 viremia was based on results from the PCV2-infected pigs (n = 11 Duroc, n = eight Landrace, and n = 10 Large White pigs) that remained in the study for the entire 35 days. PCV2 viremia was determined to be 4.6 ± 0.2 weeks in Duroc pigs (n = 11), 4.3 ± 0.6 weeks in Landrace pigs (n = eight), and 4.2 ± 0.4 weeks in Large White pigs (n = 10), and was not significantly different (P = 0.669) among breeds. Mean copy number of PCV2 genomic DNA in serum was not different (P > 0.05) among the breeds at PID 7, 14, 21, 28, and 35.
Macroscopic lesions
There were no gross lesions in the noninfected control pigs at the PID-21 or PID-35 necropsy dates. At PID 21, 9 of 12 PCV2-inoculated Duroc, 10 of 11 PCV2-inoculated Landrace, and 11 of 11 PCV2-inoculated Large White pigs had gross lymph node enlargement scores of 1.4 ± 0.3, 2.0 ± 0.3, and 1.6 ± 0.2, respectively. In addition, three Landrace pigs and one Duroc pig had mottled-tan lungs that failed to collapse. The Landrace pig that had to be euthanized at PID 21 (prior to scheduled necropsy at PID 35) was emaciated, had a pale liver, and had moderate edema throughout the stomach wall. At PID 35, gross lesions in the PCV2-inoculated pigs were limited to enlargement of lymph nodes in 9 of 11 Duroc (mean 1.8 ± 0.3), 5 of 8 Landrace (mean 1.4 ± 0.4), and 5 of 10 Large White (mean 0.9 ± 0.3) pigs.
Microscopic lesions and amount of PCV2-antigen
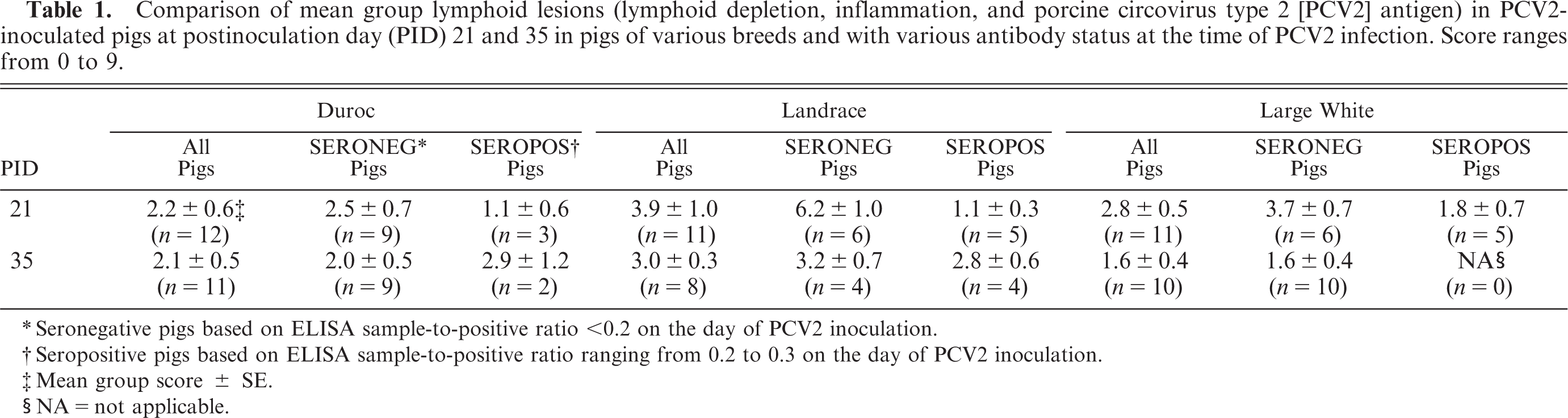
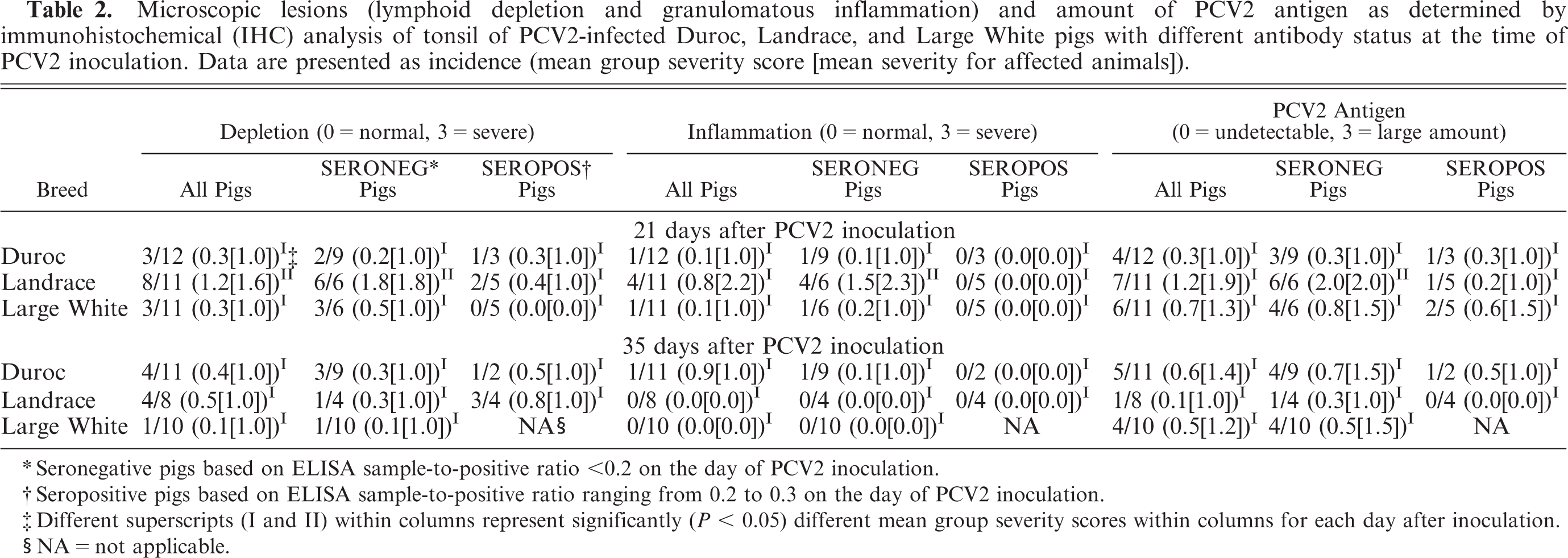
There were no microscopic lesions in any of the control pigs (data not shown). A total of 7 of 63 PCV2-infected pigs did not have PCV2-associated lymphoid lesions (overall score, 0) and 35 of 63 had mild PCV2-associated lymphoid lesions (overall score, 1–3). In 18 of 63 PCV2-infected pigs, lymphoid lesions were moderate (overall score, 4–6), and 3 of 63 pigs had severe lymphoid lesions consistent with PMWS (overall score, 7–9) (Table 1). If present, PCV2-associated lesions were characterized by mild-to-severe lymphoid depletion and mild-to-severe granulomatous inflammation in lymphoid tissues associated with low to high amounts of PCV2-antigen (Tables 2, 3). Landrace pigs had significantly (P < 0.05) more severe lymphoid depletion scores in tonsil at PID 21 (Table 2) and in the lymph node pool at PID 35, compared with those for Duroc and Large White pigs (Table 3).
Comparison of mean group lymphoid lesions (lymphoid depletion, inflammation, and porcine circovirus type 2 [PCV2] antigen) in PCV2-inoculated pigs at postinoculation day (PID) 21 and 35 in pigs of various breeds and with various antibody status at the time of PCV2 infection. Score ranges from 0 to 9.

Seronegative pigs based on ELISA sample-to-positive ratio <0.2 on the day of PCV2 inoculation.
Seropositive pigs based on ELISA sample-to-positive ratio ranging from 0.2 to 0.3 on the day of PCV2 inoculation.
Mean group score ± SE.
NA = not applicable.
At PID 21, lymphohistiocytic hepatitis was mild to moderate in 3 of 12 Duroc and 6 of 11 Landrace pigs, and was mild in 6 of 11 Large White pigs. Lymphohistiocytic myocarditis was moderate in 1 of 12 Duroc, mild to moderate in 4 of 11 Landrace, and mild in 2 of 11 Large White pigs. Lymphohistiocytic nephritis was mild in 3 of 12 Duroc pigs and was mild to moderate in 3 of 11 Landrace and 3 of 11 Large White pigs.
At PID 35, lymphohistiocytic hepatitis was mild to moderate in 4 of 11 Duroc pigs, and was mild in 4 of 8 Landrace and 1 of 10 Large White pigs. Lymphohistiocytic myocarditis was mild in 1 of 11 Duroc, 2 of 8 Landrace, and 0 of 10 Large White pigs. Mild lymphohistiocytic nephritis was observed in 5 of 11 Duroc, 3 of 8 Landrace, and 3 of 10 Large White pigs.
Lung lesions were characterized by mild, focal-to-diffuse, lymphohistiocytic interstitial pneumonia with type-2 pneumocyte hypertrophy and hyperplasia in 5 of 12 Duroc and 4 of 11 Large White pigs. Lymphohistiocytic interstitial pneumonia was mild to moderate and focal to diffuse in 8 of 11 Landrace pigs at PID 21. At PID 35, mild, focal-to-diffuse, lymphohistiocytic interstitial pneumonia was present in 6 of 11 Duroc, 5 of 8 Landrace, and 5 of 10 Large White pigs.
Three Landrace pigs had microscopic lesions consistent with PMWS, with severe lymphoid depletion characterized by complete loss of the follicular structure of all lymphoid tissues evaluated, severe histiocytic replacement of lymphoid follicles, and high amounts of PCV2 antigen associated with the lesions. These pigs also had mild to moderate lymphohistiocytic inflammation in other organ systems including heart, kidney, liver, and thymus. There also was moderate to severe granulomatous enteritis with moderate numbers of multinucleated giant cells present in Peyer's patches of the ileum associated with moderate to high amounts of PCV2-antigen.
Evaluation of the influence of passively acquired antibodies on PCV2-associated lesions and viremia among breeds
There was a total of 5 of 23 PID-0 seropositive Duroc, 9 of 19 PID-0 seropositive Landrace, and 5 of 21 PID-0 seropositive Large White pigs on the day of PCV2 inoculation. To delineate any possible influence of low amounts of passively acquired anti-PCV2 antibodies, the outcomes of the PID-0 seronegative pigs of all breeds were compared with each other. Similarly, the outcomes of the PID-0 seropositive pigs among all breeds were compared with each other. Separation of the weekly weight gain data into PID-0 seronegative and PID-0 seropositive pigs did not result in major changes, compared with the weight gain data that included PID-0 seronegative and PID-0 seropositive pigs and was not significantly different (P > 0.05) among the breeds (data not shown). Mean PCV2 genomic copy number for the PID-0 seronegative pigs was not significantly (P > 0.5) different among the breeds at PID 7, 14, 21, 28, and 35. Likewise, no significant difference (P > 0.05) was observed among breeds in mean PCV2 copy number for the PID-0 seropositive pigs. At PID 21, PID-0 seronegative Landrace pigs had a significantly (P = 0.01) higher mean group overall lymphoid severity score, compared with that for PID-0 seronegative Duroc pigs (Table 1). The PID-0 seronegative Landrace pigs had significantly (P < 0.5) more severe depletion of the lymph nodes at PID 21 and 35 (Table 3), and significantly (P < 0.5) more severe lymphoid depletion, inflammation, and PCV2 antigen associated with these lesions in tonsils at PID 21, compared with that in PID-0 seronegative Duroc and Large White pigs (Table 2). There was a higher incidence of microscopic lung, liver, heart, and kidney lesions in PID-0 seronegative, compared with PID-0 seropositive pigs across breeds (data not shown), but this difference was not significant (P > 0.5).
Microscopic lesions (lymphoid depletion and granulomatous inflammation) and amount of PCV2 antigen as determined by immunohistochemical (IHC) analysis of tonsil of PCV2-infected Duroc, Landrace, and Large White pigs with different antibody status at the time of PCV2 inoculation. Data are presented as incidence (mean group severity score [mean severity for affected animals]).
Seronegative pigs based on ELISA sample-to-positive ratio <0.2 on the day of PCV2 inoculation.
Seropositive pigs based on ELISA sample-to-positive ratio ranging from 0.2 to 0.3 on the day of PCV2 inoculation.
Different superscripts (I and II) within columns represent significantly (P < 0.05) different mean group severity scores within columns for each day after inoculation.
NA = not applicable.
Microscopic lesions (lymphoid depletion and granulomatous inflammation) and amount of PCV2 antigen as determined by IHC analysis in the lymph node pool (tracheobronchial, superficial inguinal, external iliac, mediastinal, and mesenteric lymph nodes) in PCV2-infected Duroc, Landrace, and Large White pigs with different antibody status at the time of PCV2 inoculation. Data are presented as incidence (mean group severity score [mean severity for affected animals]).
Seronegative pigs based on ELISA sample-to-positive ratio <0.2 on the day of PCV2 inoculation.
Seropositive pigs based on ELISA sample-to-positive ratio ranging from 0.2 to 0.3 on the day of PCV2 inoculation.
Different superscripts (I and II) within columns represent significantly (P < 0.05) different mean group severity scores within columns for each day after inoculation.
NA = not applicable.
In this study, the total incidence of microscopic lesions consistent with PMWS was 15.8% (3 of 19) for Landrace pigs, whereas none of the Duroc and Large White pigs developed lesions consistent with PMWS. When separated into PID-0 seronegative and PID-0 seropositive, the PMWS incidence increased to 30% (3 of 10) in PID-0 seronegative Landrace pigs.
Within-breed influence of passively acquired antibodies on PCV2-associated lesions and viremia
To further explore the possible influence of presence of the amounts of passively acquired PCV2 antibodies at the time of PCV2 inoculation on PCV2 infection, outcomes of PID-0 seropositive and PID-0 seronegative pigs within each breed were compared with each other. At PID 14 and 21, PID-0 seronegative Landrace pigs (n = 10) had significantly (P < 0.01) higher amounts of PCV2 nucleic acids in serum, compared with those in PID-0 seropositive Landrace pigs (n = 9). At PID-21 necropsy, mean overall lymphoid severity score was significantly (P < 0.002) higher in PID-0 seronegative Landrace, compared with PID-0 seropositive Landrace pigs (mean score: 6.2 ± 1.0 in PID-0 seronegative pigs [n = 6] versus 1.1 ± 0.3 in PID-0 seropositive pigs [n = 5]) (Table 1). There were no significant differences in Duroc or Large White breeds between PID-0 seronegative and PID-0 seropositive pigs, although PID-0 seronegative pigs had higher values (Table 1).
Evaluation of littermates of the PMWS-affected pigs
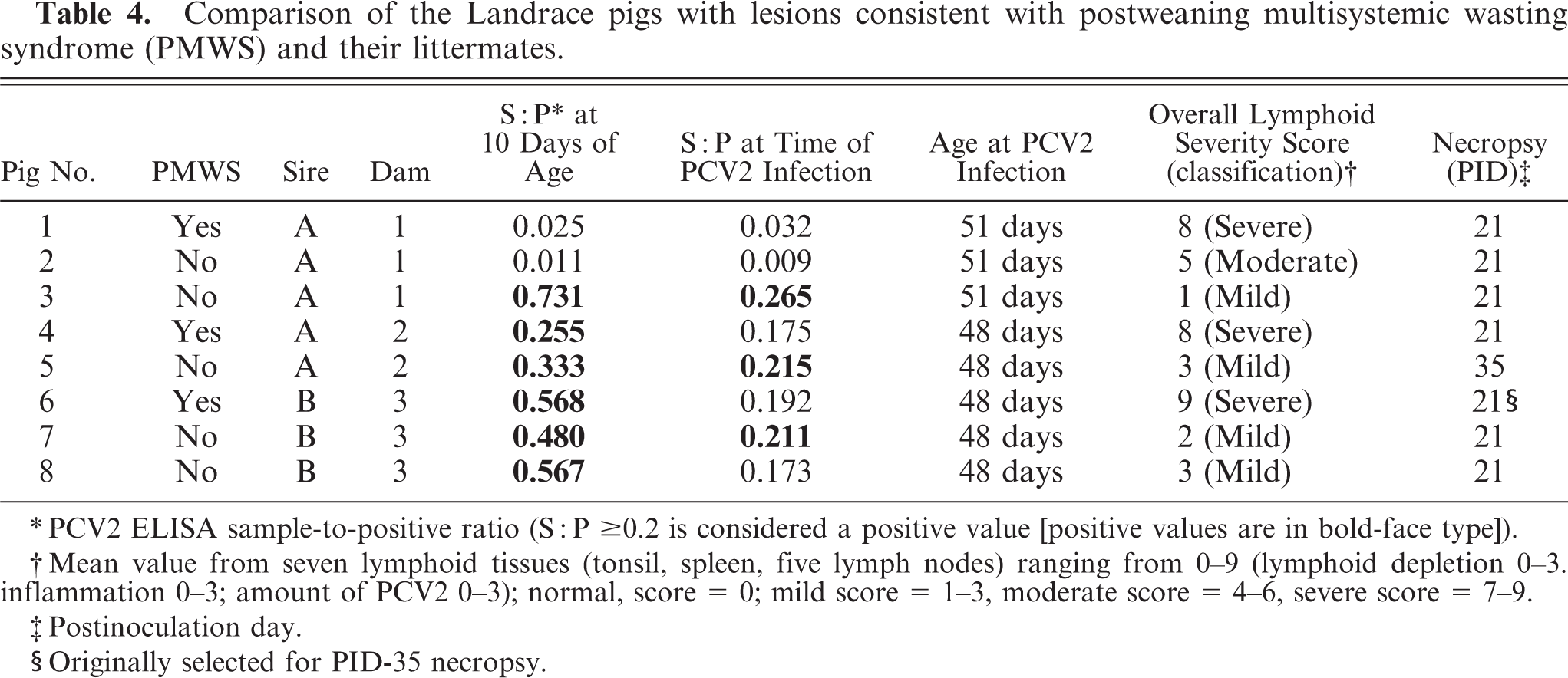
There was no litter or sire effect on the development of PCV2-associated lesions. The three Landrace pigs with microscopic lesions typical of PMWS were from three litters, and two sires and were housed in two rooms and three pens. Comparison of data (serologic, overall lymphoid lesions score) among the three Landrace pigs with lesions consistent with PMWS and their littermates is summarized in Table 4.
Comparison of the Landrace pigs with lesions consistent with postweaning multisystemic wasting syndrome (PMWS) and their littermates.
PCV2 ELISA sample-to-positive ratio (S: P ≥ 0.2 is considered a positive value [positive values are in bold-face type]).
Mean value from seven lymphoid tissues (tonsil, spleen, five lymph nodes) ranging from 0–9 (lymphoid depletion 0–3. inflammation 0–3; amount of PCV2 0–3); normal, score = 0; mild score = 1–3, moderate score = 4–6, severe score = 7–9.
Postinoculation day.
Originally selected for PID-35 necropsy.
Discussion
The overall goal of this study was to conduct a controlled experiment to investigate the influence of host genetics on susceptibility to PCV2. We studied Duroc, Landrace, and Large White pigs because they were available from a single source. Today, all three of these breeds are used extensively worldwide in purebred and crossbred breeding programs.
A limited number of peer-reviewed manuscripts have compared disease susceptibility in various pig breeds. The muscle load of Sarcocystis miescheriana bradyzoites after oral exposure was found to be 20 times higher in Pietrain, compared with Meishan pigs. 60 Compared with Meishan pigs, Large White pigs were found to be more susceptible to infection with pseudorabies virus 59 or K88+ Escherichia coli. 15 Compared with Duroc and Meishan pigs, Hampshire pigs developed significantly less severe macroscopic lung lesions after experimental PRRSV inoculation. 27 Meishan pigs were found to be highly resistant to K88ac+ E. coli-induced disease, whereas there were both susceptible and disease-resistant pigs within the Fengjjing and Minzhu breeds. 49
A cohort study was conducted on four PMWS-affected farms to investigate a suspected decreased susceptibility to PCV2-associated disease in Pietrain pigs by manipulating the genetics via artificial insemination. Half of the sows were inseminated with Pietrain semen, whereas the remaining sows received the semen that was typically used on these farms. 62 The PCV2-associated disease in the Pietrain offspring did not differ from that observed in other pigs on these farms in terms of PCV2 seroconversion, morbidity, and mortality. We did not include Pietrain pigs in the study reported here because the farm from which we purchased our purebred pigs did not raise Pietrain pigs and we believed it important to minimize confounding factors (differences in environment, production style, coinfection status) typical in pigs derived from various sources.
All breeds of this study appeared to be equally susceptible to PCV2 infection. There were no significant differences in viremia duration, amount of PCV2 genomic DNA copies, or anti-PCV2 antibody production among the breeds. However, clinical disease and microscopic lesions consistent with PMWS were only observed in the Landrace pigs. The results indicate that the Landrace pigs of this study were predisposed to progress from subclinical PCV2 infection to clinical PMWS.
To the best of our knowledge, all other confounding factors were similar across breeds in this study. In the field, PMWS often resembles an end-stage disease complex, and multiple pathogens in addition to PCV2 often are detected. 57 Experimental coinfection of pigs with PCV2 and other viruses such as PPV 7, 36, 40, 52 or PRRSV, 2, 32, 64 or bacteria such as M. hyopneumoniae 54 has been documented to enhance the amount of PCV2 genomic copies and PCV2-associated lesions and to increase the incidence of PMWS. There may be a variety of other known and unknown pathogenic and nonpathogenic organisms that are able to trigger progression of PCV2 infection to PMWS. To account for this, we believed it important that all pigs came from the same farm, were farrowed in the same barn, and were transported together to our research facilities, thereby equalizing the microbiological flora across breeds as much as possible.
Sex can be a confounding factor as well. There are at least two studies that concluded male pigs were more susceptible to development of PMWS than were female pigs, and the authors thought this might be attributable to castration and associated secondary infections. 11, 61 Thus, in our study, we used only female pigs to control for sex.
To our knowledge, all breeding herds are infected with PCV2. We usually derive PCV2-negative crossbred pigs for our experimental PCV2 inoculation trials by screening the entire batch of dams scheduled to farrow during the desired timeframe. We then select piglets from these sows in the batch that have the lowest anti-PCV2 antibody levels. 56 Owing to litter and sire restrictions and to the low number of purebred sows within the same farrowing batch, such was not possible in this study. We, therefore, did not screen sows prior to purchase of the pigs, and a portion of the pigs had high levels of maternal antibodies at time of arrival. Compared with that in the other two breeds, the Landrace pigs of this study had higher levels of PCV2 antibodies at weaning and at the time of PCV2 inoculation, but the difference was not statistically significant (P > 0.5). Because of the higher incidence and titer of maternal PCV2 antibodies, we increased the number of procedural control Landrace pigs (six rather than three), which decreased the number of experimentally infected pigs within this breed. Because of budget and room restrictions, experimental infection of the pigs was started prior to complete waning of the maternal antibody titer in these pigs.
We were also concerned about a possible but unknown host age effect on PCV2 infection. At 5 to 8 weeks of age, 19 of 63 pigs (30.1%) still had low levels of passively acquired antibodies to PCV2 at the time of PCV2 inoculation. There were 47.4% (9 of 19) of the experimentally infected Landrace pigs that had low amounts of PCV2-specific antibodies at the time of PCV2 inoculation, compared with 21.8% (5 of 23) of the Duroc and 23.8% (5 of 21) of the Large White pigs. Landrace piglets that had low but measurable maternal antibodies at the time of PCV2 inoculation had significantly (P < 0.05) reduced overall PCV2-associated lesions, compared with Landrace piglets that were seronegative at the time of PCV2 inoculation. All three Landrace pigs that developed lesions consistent with PMWS were seronegative at the time of PCV2 inoculation, as were the 18 Duroc, 16 Large White, and seven additional Landrace pigs that did not develop PMWS. The presence of anti-PCV2 antibodies has been documented to protect pigs from developing severe PCV2-associated lesions. 4, 22 Our experimental results support this conclusion and indicate further that only a portion of the anti–PCV2-negative pigs are susceptible to PCV2-associated diseases. It has been suggested that PCV2-ELISA S ° P of 0.6 prevents PCV2 replication and development of PMWS. 33 On the basis of the results of our study, even lower levels of passively acquired antibodies are protective against development of severe PCV2-associated lesions. This suggests that PCV2 vaccination of breeding herds resulting in high passively transferred antibody titer may be one way to control PCV2-associated disease in growing pigs.
We were able to induce lesions consistent with PMWS in 3 of 19 (15.8%) Landrace pigs, whereas we were not able to induce PMWS in any of the Duroc or Large White pigs. This is comparable to what is seen in commercial crossbred swine production in the field where the incidence of PMWS in affected herds usually peaks with around 10% of the herd affected, but may vary from as little as 4% to as high as 30% in individual farms. 1, 28, 43, 61
In this study, we used a genetically well-characterized PCV2 isolate 20 that has been used extensively in pig models. Previously, singular PCV2 infection with the same isolate at a similar cell culture passage and in approximately the same dose as used in the purebred pigs of this current study did not result in expression of clinical disease or presence of microscopic lesions consistent with PMWS in 264 crossbred pigs that were part of nine studies. 21– 24, 35, 52– 55 This further supports our conclusion that the purebred Landrace pigs used in this study were more susceptible to PCV2-associated diseases.
It appears to be a common belief among some producers and veterinarians that Landrace pigs are more susceptible to infectious diseases in general; however, reports describing experimental results that substantiate this clinical impression are lacking to our knowledge. On the basis of field observations, it has been suggested that Landrace pigs in the UK appeared to be less affected by progressive atrophic rhinitis than are Large White pigs. 13 Landrace pigs are known to be predisposed to type-III congenital tremor, which is attributable to a monogenic sex-linked factor resulting in a deficiency of oligodendrocytes in the spinal cord of male Landrace pigs. 29 Danish Landrace pigs are thought to be the origin of dermatosis vegetans, which is caused by a semilethal autosomal recessive factor. 14
More studies including larger numbers of Landrace pigs from several lines and various geographic locations are warranted to further investigate a possible predisposition to PCV2-associated PMWS in this breed. Genetic resistance may be associated with a single gene, but is usually associated with multiple genes. Another approach may be to further study PMWS-affected pigs and nonaffected littermates (who share at least 50% of their genes in common) by gene array analysis to possibly identify gene loci with differences in expression that may be responsible for increased susceptibility to PCV2-associated diseases. In conclusion, despite its limited size, this work adds to the body of evidence to explain differences in clinical expression of PCV2-associated diseases from farm to farm.