
Abstract
Callithrix jacchus (common marmoset) is regularly used in biomedical research, including for studies involving the skeleton. To support these studies, skeletons of healthy animals that had been euthanized for reasons not interfering with skeletal anatomy were prepared. The marmoset dental formula 2I-1C-3P-2M of each oral quadrant is atypical for New World monkeys which commonly possess a third molar. Seven cervical, 12–13 thoracic, 7–6 lumbar, 2–3 sacral and 26–29 caudal vertebrae are present, the thoracolumbar region always comprising 19 vertebrae. A sigmoid clavicle connects the scapula with the manubrium of the sternum. Depending on the number of thoracic vertebrae, 4–5 sternebrae are located between the manubrium and xiphoid process. Wide interosseous spaces separate the radius from the ulna, and the tibia from the fibula. A small sesamoid bone is inserted in the m. abductor digiti primi longus at the medial border of the carpus, a pair of ovoid sesamoid bones is located at the palmar/plantar sides of the trochleae of each metapodial bone, and round fabellae articulate with the proximal surfaces of the femoral condyles. Male marmosets possess a small penile bone. Both the front and hind feet have five digits. The hallux possesses a flat nail, whereas all other digits present curved claws. Interestingly, a central bone is present in both the carpus and tarsus. This study provides a description and detailed illustrations of the skeleton of the common marmoset as an anatomical guide for further biomedical research.
For osteological studies, the translation of data from rodents to man is hampered by the delayed epiphyseal closure with continuous bone modelling and the absence of cortical bone osteons in the former species. 1 By contrast, the common marmoset (Callithrix jacchus) resembles man in bone structure and remodelling, rendering it an excellent non-human primate animal model for osteological studies. 1,2 In contrast to other non-human primates, common marmosets are small animals, which translates into relatively small housing facilities, with low feeding and housing costs. Together with the easy handling and relatively short reproduction time, marmosets offer very advantageous animal models for biomedical research. 3–6 Osteological studies using marmosets include the research on vitamin D-dependent rickets, osteomalacia, osteopenia (a loss of 10–25% of bone mass) and osteoporosis (a further loss of bone mass). 5
The common marmoset is a New World monkey (Infraorder Platyrrhini, family of the Callithrichidae) with body lengths of 14–18 cm and tail lengths of 25–30 cm. These diurnal, arboreal animals originate from the South American tropical rainforest and weigh between 300 and 450 g. They are omnivorous and feed naturally with tree gum, fruit, nuts, insects and small vertebrates. 7 Both in their natural habitats and in captivity they live in family groups led by a dominant, monogamous parent pair. 8 After a pregnancy of approximately 145 days, twins (or often triplets in captivity) with chimaeric bone marrow due to the fusion of the placental bloodstreams, are born. 9,10 Sexual maturity is reached at the age of 1.5–2 years and the life expectancy of captive animals amounts to 16 years. 1
Marmosets, which have been kept as pets since the French baroque times, were introduced in biomedical research in the early 1960s. 11–13 Since then, the number of studies using common marmosets has progressively increased. In 1992, the European Marmoset Research Group was founded to optimize the research performed on marmosets which are now a frequently studied laboratory species. 11 Despite its increased use, the specific skeletal features of this species are poorly documented. 1 Hershkovitz 14 gives an extended overview of the anatomy of the living New World monkeys, but no detailed information is provided on the anatomy of the postcranial bones of the common marmoset. Wagner and Kirberger, 13 while investigating the radiographic anatomy of C. jacchus, focused on the internal organs and provided only a few descriptive osteological details. Radiographies have also been used for age determination of common marmosets based on tooth development, craniofacial development, and the length and stage of ossification of the long bones. 15–19 By contrast, many authors have investigated the calcium metabolism in Callithrichidae as they require exceptionally large amounts of vitamin D3 due to the end-organ resistance to this metabolite. 20–23
Because of the paucity of specific osteological data, the present article provides a detailed description of the skeleton of C. jacchus to serve as a guide for further osteological and arthrological research. The text of this paper focuses on the main features of the skeleton, while the figures illustrate the various parts of the skeleton in more detail. Common English anatomical terminology is used throughout the text while the figure captions also provide the official Latin terms. 24
Materials and methods
Animals
To reduce the number of laboratory animals, frozen cadavers of 10 adult (2–8 years of age), healthy common marmosets (C. jacchus) of both genders (7 females and 3 males), two one-day-old males and two one-month-old marmosets of each gender were obtained from the Biomedical Primate Research Centre, Rijswijk, The Netherlands where they had been used in other studies. All animals were born and raised at this centre in natural family groups. The adult marmosets used in this study remained in their birth group until they were 18 months old. The breeding facilities were enriched with branches and toys, contained a biofloor and allowed free access to outside pens. Adult marmosets were subsequently moved to experimental units where they were housed as same-sex pairs in spacious cages enriched with branches and toys, and with padded shelter provided on the floor. The daily diet was composed of commercial food pellets for New World monkeys (Sniff, Soest, Germany) supplemented with limited amounts of fresh fruit, arabic gum and slurry enriched with vitamin D. Drinking water was provided ad libitum. All adult animals had been euthanized for various reasons not related to this study. The one-day-old neonates were born as part of triplets and euthanized since marmosets can only feed two babies. The one-month-old animals were euthanized to prevent severe suffering from maternal agalactia. All housing, care and use of animals were in accordance with the Dutch law on animal experimentation, including all permits and approvals.
Preparation of skeletons
After thawing, the soft tissues of the cadavers were first removed manually by anatomical dissection. Subsequently, the soft tissue remnants were digested by Dermestid beetles. Just before complete disintegration of the skeletons they were placed in a solution containing approximately 10% hydrogen peroxide (Univar Benelux, Brussels, Belgium) to bleach the bones and macerate the smaller tissue remnants. The skeletons were finally degreased with methylene chloride (Univar Benelux).
Data collection
The skeletons were macroscopically examined. The young animals were used to describe the deciduous dentition and the cranial sutures. The rostrocaudal dimensions of the skull, the lengths of the various segments of the vertebral column including the tail, the length of the hip bone and the lengths of the appendicular long bones, from the most proximal to the most distal extremities, were determined on the 10 adult animals using digital callipers. To detect potential differences in skeletal dimensions between the three male and seven female common marmosets a Student's t-test was performed in Excel 2011 (Microsoft, Zaventem, Belgium). Differences were regarded as statistically significant when P< 0.05.
Macroscopic pictures of the various skeletal structures were taken with a digital photo camera (Canon EOS 50D, Canon Belgium, Diegem, Belgium) and served as a basis for the anatomical description of the skeleton. To illustrate smaller structures such as the teeth and the ossicles of the middle ear a stereomicroscope (SZX7, Olympus Belgium, Aartselaar, Belgium) linked to a digital camera (ColorView, Olympus Belgium) was used. The topography of the carpal bones was visualized by scanning the carpal joints using an in-house developed microcomputed tomography system. This system consisted of an open-type X-ray system (Feinfocus, Garbsen, Germany) with transmission target and a VHR detector (Photonics Science, Millham, UK). A total of 800 projections were taken. Three-dimensional images were constructed with OCTOPUS, which is an in-house developed software package, 25 and were further processed in MIMICS® (Materialise, Leuven, Belgium).
Results
Skull
Skull lengths and widths, and lengths of the appendicular bones and vertebral regions of the seven female (F1–F7) and three male (M1–M3) common marmosets with the mean value ± standard deviation (SD) (all values in mm)
The neurocranium is much longer than the splanchnocranium. The very large conical orbits are almost completely postorbitally closed as only narrow dorsal and ventral orbital fissures are present. The cranial sutures, particularly visible on the neonatal skull, include the frontal or metopic suture between the frontal bones, the sagittal suture between the parietal bones, the coronal suture between the frontal and parietal bones, and the lambdoid suture between the parietal and occipital bones. A suture between the zygomatic and parietal bones is present in the pterion area. The mandible and in particular its body is relatively large. A prominent mandibular angle can be seen. The symphysis between the left and right mandibles is already closed at birth (Figure 1).
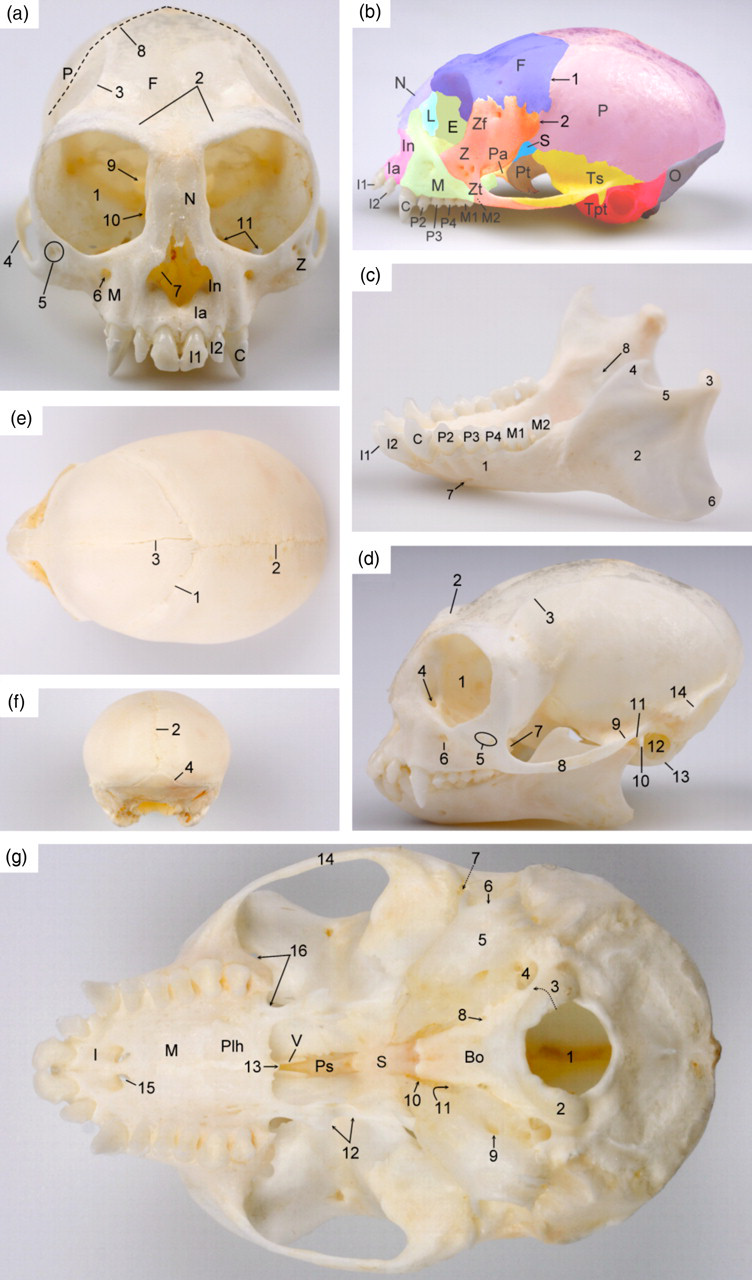
Skull of the common marmoset. (a) Frontal view of the skull of an adult marmoset: 1 = orbit (orbita); 2 = superciliary ridge (arcus supraciliaris); 3 = temporal line (linea temporalis); 4 = zygomatic arch (arcus zygomaticus); 5 = zygomaticofacial foramina (foramina zygomaticofacialia); 6 = infraorbital foramen (foramen infraorbitale); 7 = ventral nasal concha (concha nasalis ventralis); 8 = coronal suture (sutura coronalis); 9 = optic foramen (foramen opticum); 10 = dorsal orbital fissure (fissure orbitalis dorsalis); 11 = ventral orbital fissure (fissura orbitalis ventralis); Ia = alveolar process of the incisive bone (processus alveolaris ossis incisivi); In = nasal process of the incisive bone (processus nasalis ossis incisivi); M = maxilla; Z = zygomatic bone (os zygomaticum); N = nasal bone (os nasale); F = frontal bone (os frontale); P = parietal bone (os parietale). I1 = first incisor (dens incisivus primus); I2 = second incisor (dens incisivus secundus); C = canine tooth (dens caninus). (b) Left lateral view of the skull of an adult marmoset showing the different bones: E = ethmoidal bone (os ethmoidale), F = frontal bone (os frontale); Ia = alveolar process of the incisive bone (processus alveolaris ossis incisivi); In = nasal process of the incisive bone (processus nasalis ossis incisivi); L = lacrimal bone (os lacrimale); M = maxilla; N = nasal bone (os nasale); O = occipital bone (os occipitale); P = parietal bone (os parietale); Pa = palatine bone (os palatinum); S = sphenoidal bone (os sphenoidale); Tpt = petrosal and tympanic parts of the temporal bone (pars petrosa and pars tympanica ossis temporalis); Ts = squamous part of the temporal bone (pars squamosa ossis temporalis); Pt = pterygoid bone (os pterygoideum); Z = zygomatic bone (os zygomaticum); Zf = frontal process of the zygomatic bone (processus frontalis ossis zygomatici); Zt = temporal process of the zygomatic bone (processus temporalis ossis zygomatici); 1 = coronal suture (sutura coronalis); 2 = zygomatic–parietal suture (sutura zygomaticoparietalis). (c) Left lateral view of the mandibles of an adult marmoset: 1 = mandibular body (corpus mandibulae); 2 = mandibular ramus (ramus mandibulae); 3 = condylar process (processus condylaris); 4 = coronoid process (processus coronoideus); 5 = mandibular notch (incisura mandibulae); 6 = mandibular angle (angulus mandibulae); 7 = mental foramen (foramen mentale); 8 = mandibular foramen (foramen mandibulae); I1 = first incisor (dens incisivus primus); I2 = second incisor (dens incisivus secundus); C = canine tooth (dens caninus); P2 = second premolar (dens premolaris secundus); P3 = third premolar (dens premolaris tertius); P4 = fourth premolar (dens premolaris quartus); M1 = first molar (dens molaris primus); M2 = second molar (dens molaris secundus). (d) Left lateral view of the skull including the mandibles of an adult marmoset: 1 = orbit (orbita); 2 = superciliary ridge (arcus supraciliaris); 3 = temporal line (linea temporalis); 4 = lacrimal canal (canalis lacrimalis); 5 = zygomaticofacial foramina (foramina zygomaticofacialia); 6 = infraorbital foramen (foramen infraorbitale); 7 = maxillary tuberosity (tuber maxillae); 8 = zygomatic arch (arcus zygomaticus); 9 = zygomatic process of the temporal bone (processus zygomaticus ossis temporalis); 10 = retroarticular process (processus retroarticularis); 11 = mandibular fossa (fossa mandibularis); 12 = external acoustic pore (porus acusticus externus); 13 = tympanic bulla (bulla tympanica); 14 = crista nuchae (nuchal crest). (e) Dorsal view of the skull of a neonatal marmoset showing the position of the foramen magnum and the different sutures: 1 = coronal suture (sutura coronalis); 2 = sagittal suture (sutura sagittalis); 3 = frontal or metopic suture (sutura frontalis). (f) Caudal view of the skull of a neonatal marmoset showing the different sutures: 2 = sagittal suture (sutura sagittalis); 4 = lambdoid suture (sutura lambdoidea). (g) Ventral view of the base of the skull of an adult marmoset: 1 = foramen magnum; 2 = occipital condyle (condylus occipitalis); 3 = opening for the hypoglossal nerve (canalis nervi hypoglossi); 4 = jugular foramen (foramen jugulare); 5 = tympanic bulla (bulla tympanica); 6 = external acoustic pore (porus acusticus externus); 7 = temporal opening (meatus temporalis); 8 = external opening of the carotid canal (apertura externa canalis carotici); 9 = stylomastoid foramen (foramen stylomastoideum); 10 = oval foramen (foramen ovale); 11 = spinous foramen (foramen spinosum); 12 = pterygoid processes (processus pterygoideus); 13 = caudal nasal spine (spina nasalis caudalis); 14 = zygomatic arch (arcus zygomaticus); 15 = palatine fissure (fissura palatina); 16 = ventral orbital fissure (fissura orbitalis ventralis); Bo = basioccipital bone (pars basilaris ossis occipitalis); S = sphenoidal bone (os sphenoidale); I = incisive bone (os incisivum); M = maxilla; Ps = presphenoidal bone (os presphenoidale); Plh = horizontal plate of the palatine bone (lamina horizontalis ossis palatini); V = vomer (os vomer)
Through the ectotympanic ring, the ossicles of the middle ear, which comprise the malleus, incus and stapes, are readily visible on lateral adspection of the skull. The malleus has no prominent lateral, rostral or muscular processes. A small lenticular bone is present between the incus and the stapes (Figure 2).
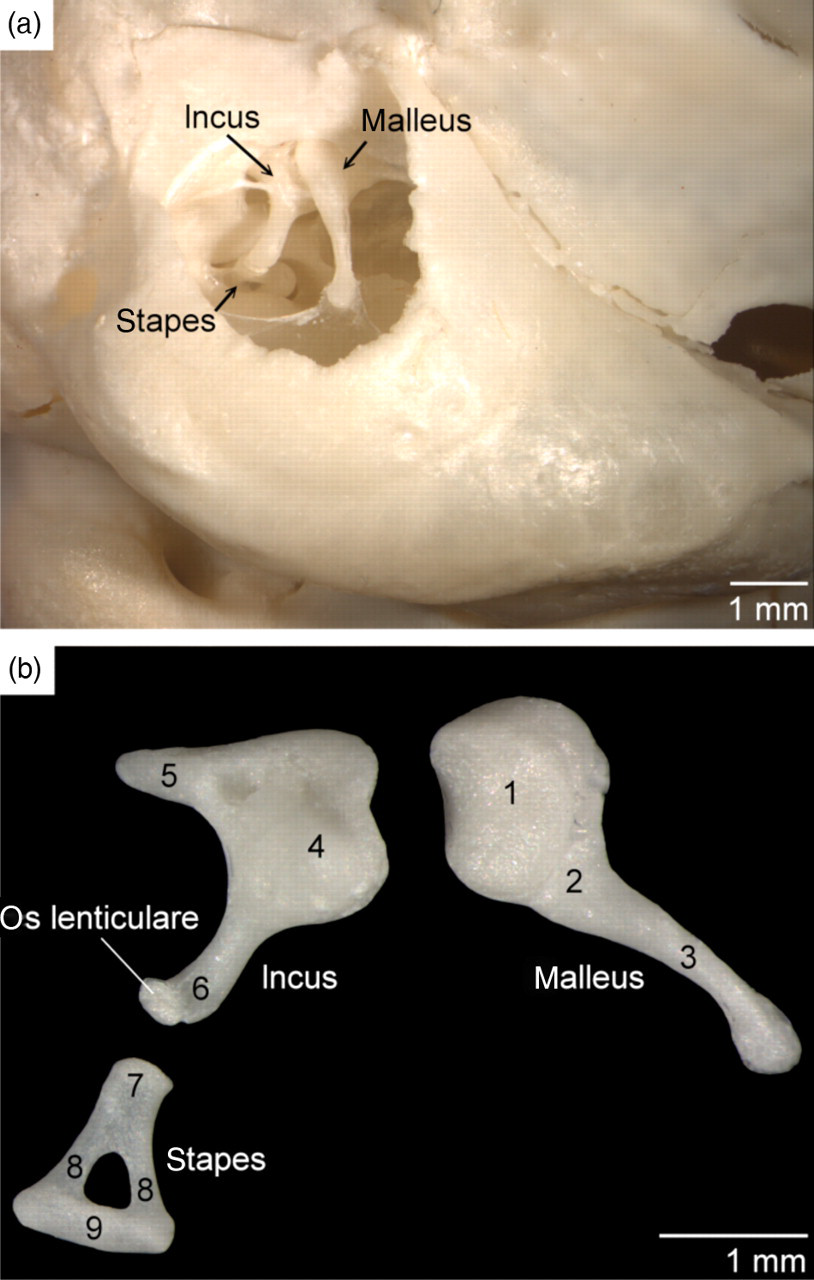
Ossicles of the middle ear of the common marmoset. (a) In situ topography of the right ossicles (lateral view). (b) Larger magnification of the ossicles: 1 = head of the malleus (caput mallei); 2 = neck of the malleus (collum mallei); 3 = manubrium of the malleus (manubrium mallei); 4 = body of the incus (corpus incudis); 5 = short process of the incus (crus breve); 6 = long process of the incus (crus longum or processus lenticularis); 7 = capitulum of the stapes (caput stapedis); 8 = crus of the stapes (crus stapedis); 9 = base of the stapes (basis stapedis)
Dentition
In the adult marmoset, each oral quadrant contains two incisors, a short-tusked canine tooth, three premolars and two molars, which results in a total number of 32 teeth. In the maxilla, the rostral (P2) and middle (P3) premolars have one root, the caudal premolar (P4) presents two roots, and both molars bear three roots (Figure 3). The sizes of the maxillar premolars are rather equal. By contrast, the first molar is markedly larger than the last. Both molars of the mandibles, which do not differ much in size, have two roots. This contrasts with the single roots present in the other mandibular teeth. The number of roots is reflected by the number of tubercles that are visible on the occlusal surfaces of the teeth (Figure 4). At birth, only the first deciduous incisors have erupted while the second deciduous incisors can be observed underneath the gums. The deciduous set of teeth, which is devoid of molars, is complete in the one-month-old marmoset (Figure 3). The number of roots of the upper and lower deciduous premolars is 1-3-2, and 1-1-2, respectively (Figure 4).
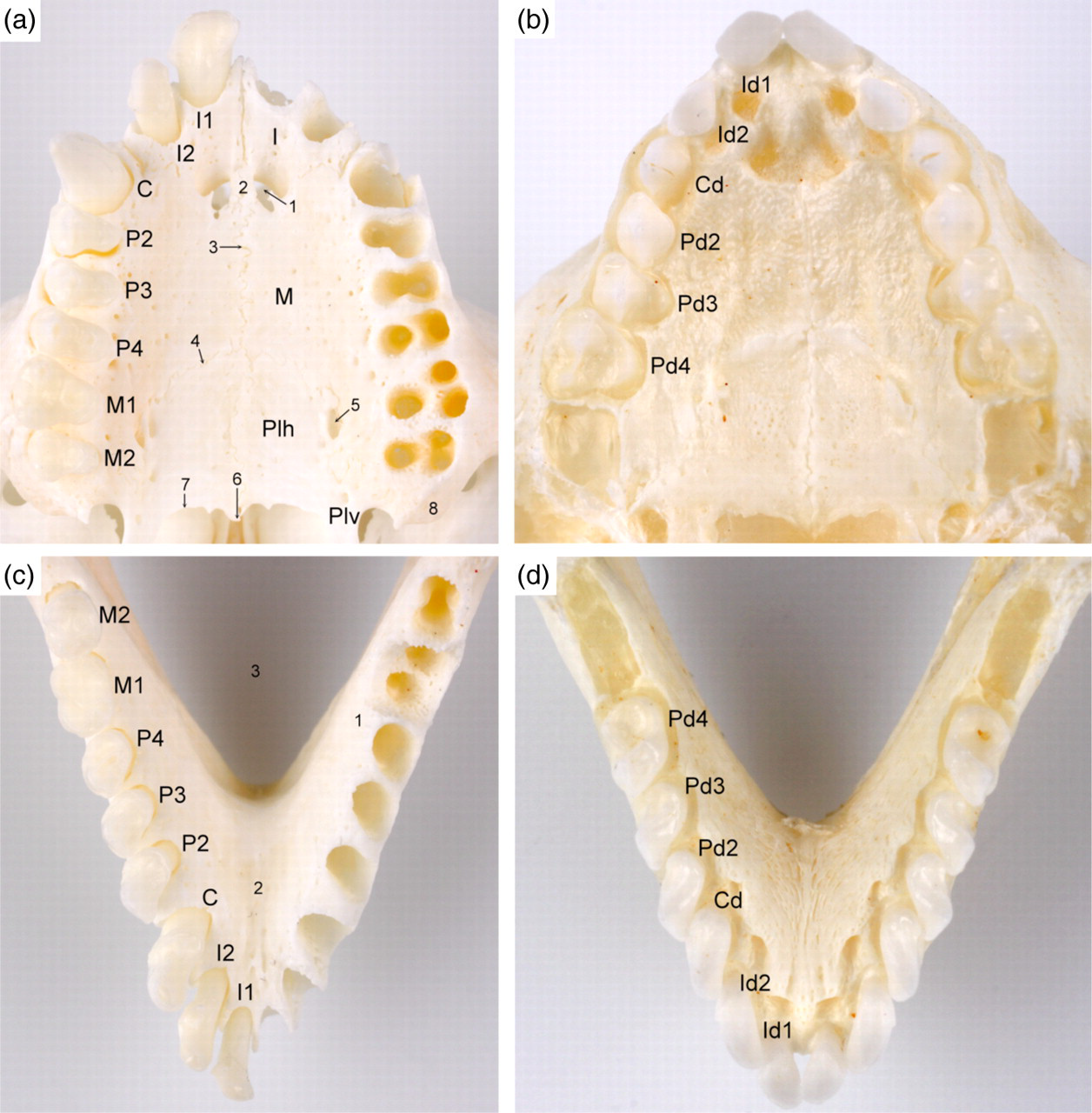
Dentition of the marmoset. (a) Ventral view of the dentition of the upper cheek in the adult marmoset: I1 = first incisor (dens incisivus primus); I2 = second incisor (dens incisivus secundus); C = canine tooth (dens caninus); P2 = second premolar (dens premolaris secundus); P3 = third premolar (dens premolaris tertius); P4 = fourth premolar (dens premolaris quartus); M1 = first molar (dens molaris primus); M2 = second molar (dens molaris secundus); I = incisive bone (os incisivum); M = maxilla; Plh = horizontal plate of the palatine bone (lamina horizontalis ossis palatini); Plv = vertical plate of the palatine bone (lamina perpendicularis ossis palatini); 1 = palatine fissure (fissura palatina); 2 = palatine process of the incisive bone (processus palatinus ossis incisivi); 3 = median palatine suture (sutura palatina mediana); 4 = transverse palatine suture (sutura palatina transversa); 5 = major palatine foramen (foramen palatinum majus); 6 = caudal nasal spine (spina nasalis caudalis); 7 = rim of the choanae; 8 = maxillary tuberosity (tuber maxillae). (b) Ventral view of the dentition of the upper cheek in a one-month-old marmoset: Id1 = first deciduous incisor (dens incisivus deciduus primus); Id2 = second deciduous incisor (dens incisivus deciduus secundus); Cd = deciduous canine tooth (dens caninus deciduus); Pd2 = second deciduous premolar (dens premolaris deciduus secundus); Pd3 = third deciduous premolar (dens premolaris deciduus tertius); Pd4 = fourth deciduous premolar (dens premolaris deciduus quartus). (c) Dorsal view of the dentition of the lower cheek in the adult marmoset: 1 = mandibular body (corpus mandibulae); 2 = intermandibular symphysis (symphysis intermandibularis); 3 = intermandibular space (spatium intermandibulare). The labelling of the dentition is similar to the upper cheek. (d) Dorsal view of the dentition of the lower cheek in a one-month-old marmoset. The labelling of the dentition is similar to the upper cheek
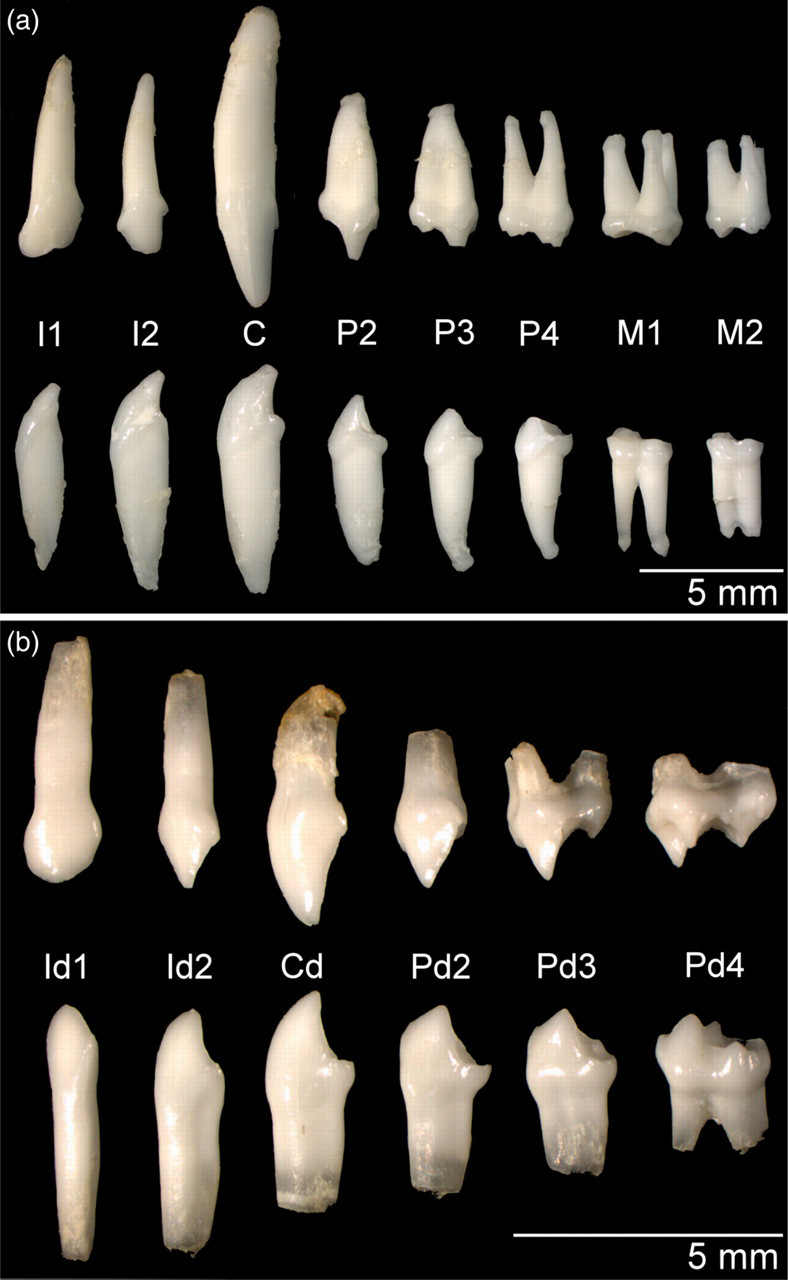
Lateral view of the left-sided teeth of the adult (a) and one-month-old (b) marmoset, respectively. The upper and lower rows represent the teeth of the upper and lower cheeks, respectively. The labelling of the dentition is analogous to Figure 3
Axial skeleton and thorax
The axial skeleton and thorax are presented in Figure 5. The vertebral column consists of seven cervical vertebrae, 12 or 13 thoracic vertebrae, seven or six lumbar vertebrae, two or three sacral vertebrae and 26–29 caudal vertebrae. The sum of the thoracic and lumbar vertebrae always amounts to 19. On the skeletons of the adult marmosets, the cervical, thoracic, lumbar, sacral and caudal regions measured 23 ± 2, 56 ± 3, 56 ± 3, 15 ± 2 and 298 ± 40 mm in length, respectively (Table 1). Any statistically significant difference between the male and female adult common marmosets could not be detected.
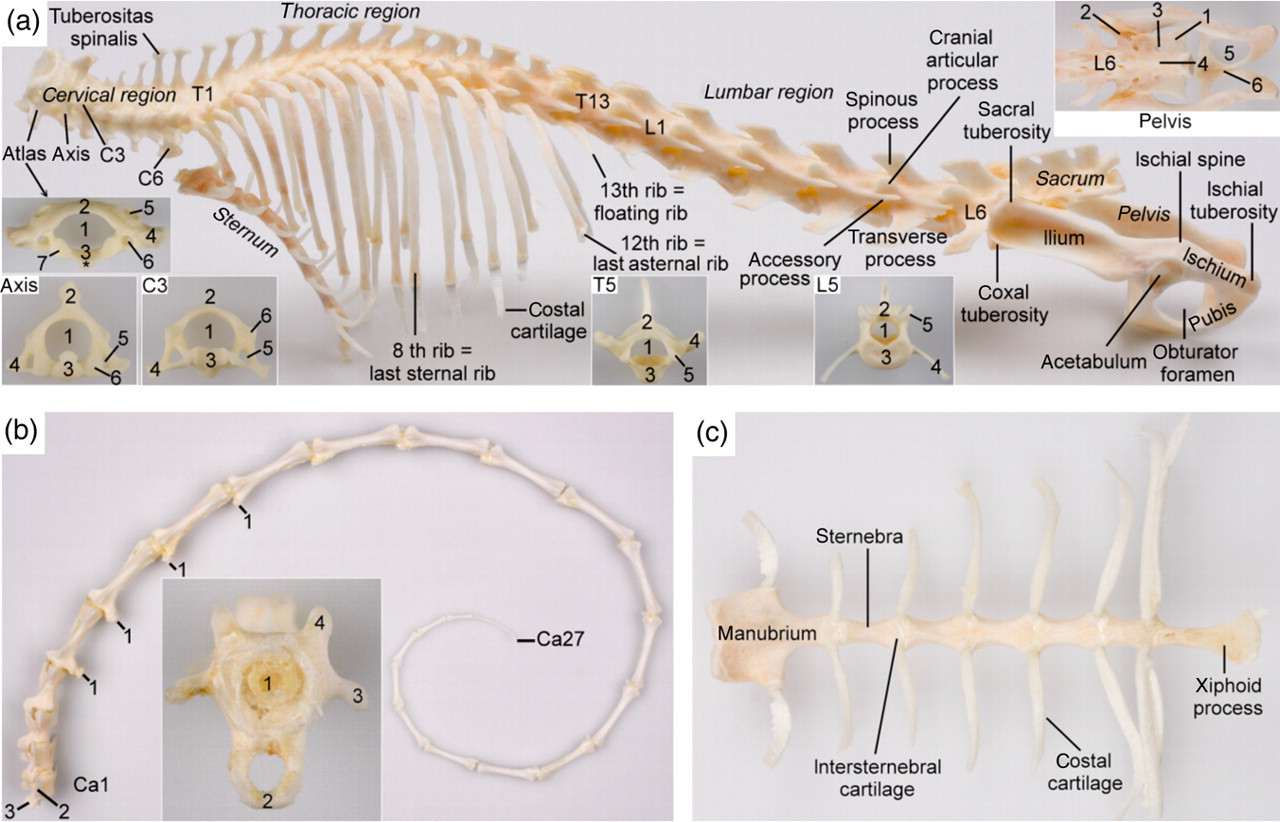
The axial skeleton of the marmoset. (a) Left lateral view of the vertebral column, thorax and pelvis. The cervical, thoracic and lumbar regions of this specimen contain 7, 13 and 6 vertebrae, respectively. The sacrum consists of three fused vertebrae. T1 = first thoracic vertebra; T13 = 13th thoracic vertebra; L1 = first lumbar vertebra; L6 = sixth lumbar vertebra. The inset of the atlas (caudal view) shows: 1 = vertebral foramen (foramen vertebrale); 2 = dorsal arch (arcus dorsalis); 3 = ventral arch (arcus ventralis) with * = ventral tuberosity (tuberculum ventrale); 4 = wing of atlas (ala atlantis); 5 = lateral vertebral foramen (foramen vertebrale laterale); 6 = transverse foramen (foramen transversarium); 7 = caudal articular fovea (fovea articularis caudalis). Inset of the axis (cranial view): 1 = vertebral foramen (foramen vertebrale); 2 = crest of the axis (crista axis); 3 = dens of the axis (dens axis); 4 = transverse process (processus transversus); 5 = transverse foramen (foramen transversarium); 6 = cranial articular surface (facies articularis cranialis). Inset of the third cervical vertebra (C3) (cranial view): 1 = vertebral foramen (foramen vertebrale); 2 = vertebral arch (arcus vertebralis) devoid of spinous process (processus spinosus); 3 = vertebral body (corpus vertebrae); 4 = transverse process (processus transversus); 5 = transverse foramen (foramen transversarium); 6 = cranial articular surface (facies articularis cranialis). The sixth cervical vertebra (C6) can easily be recognized by its ventral lamina (lamina ventralis). Inset of the fifth thoracic vertebra (T5) (cranial view): 1 = vertebral foramen (foramen vertebrale); 2 = vertebral arch (arcus vertebralis) with spinous process (processus spinosus); 3 = vertebral body (corpus vertebrae); 4 = transverse process (processus transversus); 5 = cranial articular surface (facies articularis cranialis). Inset of the fifth lumbar vertebra (L5) (cranial view): legend similar to that of T5. Inset of the pelvis (dorsal view): 1 = sacral wing (ala sacralis); 2 = auricular surface (facies auricularis); 3 = dorsal sacral foramen (foramen sacrale dorsale); 4 = median sacral crest (crista sacralis mediana); 5 = obturator foramen (foramen obturatum); 6 = pelvic symphysis (symphysis pelvina). (b) Vertebrae of the tail (lateral view): Ca1 = first caudal vertebra; Ca27 = last caudal vertebra; 1 = vertebral body (corpus vertebrae); 2 = arcus haemalis (hemal arch); 3 = transverse process (processus transversus); 4 = cranial articular process (processus articularis cranialis). (c) Ventral view of the sternum
The bodies of the cervical vertebrae are short. The atlas is characterized by small, rectangular wings and large lateral vertebral foramina. The crest of the axis is very prominent. The spinous processes increase in length from the third to the seventh cervical vertebra. On the last two cervical vertebrae, the tips of the spinous processes form heavy tuberosities. The sixth cervical vertebra can easily be recognized by its ventral lamina.
The bodies of the thoracic vertebrae elongate towards the lumbar region while the spinous processes shorten and become broader. These processes point in a caudal direction up to the eighth or ninth thoracic vertebra. The spinous processes of the last three or four thoracic vertebrae, as well as those of the lumbar vertebrae, point in a cranial direction. As a result, the anticlinal vertebra is T9 or T10.
Although the number of lumbar vertebrae is only half that of the thoracic vertebrae, the lengths of the thoracic and lumbar regions are similar. The very long lumbar vertebrae possess prominent spinous and cranioventrally inclined transverse processes that become larger towards the sacrum. The last lumbar vertebra is always shorter than the other lumbar vertebrae. Only a narrow lumbosacral interarcual space (spatium interarcuale lumbosacrale) is present between the most caudal lumbar vertebra and the sacrum.
In most cases the sacrum is composed of three fused vertebrae. In one animal, only two sacral vertebrae were present. In another marmoset a transitional vertebra was located in between the last lumbar vertebra and the sacrum that was composed of two fused sacral vertebrae. A transitional vertebra between the second sacral vertebra and the tail was seen once. Although the bodies of the sacral vertebrae are fused, the transverse processes of the first can be recognized individually. These processes, together with the cranial third of the sacral wings that are formed by the transverse processes of the subsequent sacral vertebrae, are connected to the auricular surface of the ilium.
The caudal vertebrae elongate towards the middle of the tail and then start to shorten again. Closed neural arches are present on the first three to four vertebrae while closed hemal arches can be recognized from the fourth to the eighth caudal vertebra.
The number of ribs is dependent on the number of thoracic vertebrae and varies between 12 and 13 pairs. However, a number of 13 thoracic vertebrae are predominant since only one of the seven female marmosets and one of the three male marmosets investigated in this study had 12 thoracic vertebrae. The seventh or eighth pair of ribs, depending on the number of thoracic vertebrae, is the last sternal pair that is directly attached to the sternum by costal cartilage. The caudal pairs are asternal ribs that have indirect connections with the sternum since their costal cartilages are attached to that of the previous rib. The last pair of ribs is very short and floating, thus lacking any connection with the sternum.
The sternum is composed of a broad manubrium, four or five cuboidal sternebrae related to the number of thoracic vertebrae, and a slender xiphoid process. Articular surfaces for the clavicles are located bilaterally on the craniolateral surfaces of the manubrium. Caudal to these, the costal cartilages of the first pair of ribs are attached. Subsequent sternebrae are connected to each other by means of intersternebral cartilages onto which the costal cartilages are attached.
Penile bone
A small penile bone, approximately 2.5 mm in length, is present at the tip of the penis. It has a slender cranial process which extends to the glans of the penis and a more voluminous caudal part (Figure 6).
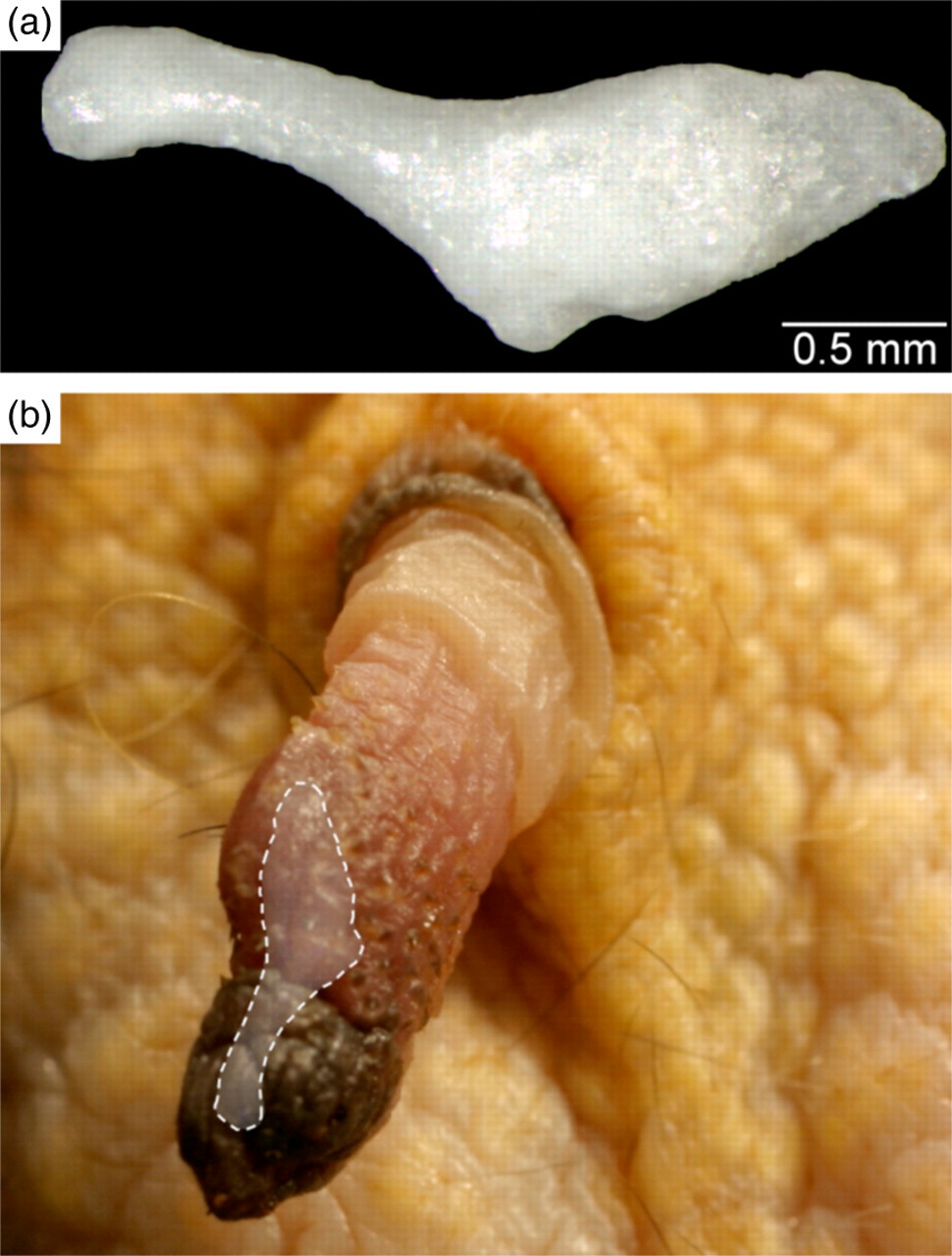
(a) Penile bone of the common marmoset. (b) In situ topography of the penile bone
Front limb
The long bones of the front limb (Figure 7) comprise the scapula, humerus, radius and ulna that measure 29 ± 1, 45 ± 1, 40 ± 1 and 47 ± 1 mm in length, respectively (Table 1). No statistically significant differences between the male and female adult common marmosets were detected. The scapula is characterized by a prominent coracoid process and an acromion that has a connection with the sigmoidal clavicle. The humerus is slender and shows a discrete humeral crest and deltoid tuberosity. By contrast, the medial epicondyle is very well developed and protrudes far medially. No supracondylar or supratrochlear foramina are present. The radius and ulna are separated by a wide interosseous space and are not fused. The carpus contains 10 carpal bones, viz. four in the antebrachial and metacarpal rows, a central carpal bone inserted axially between both rows and a sesamoid bone located in the abductor muscle of the thumb (sesamoid bone of the m. abductor pollicis longus). Axial and abaxial ovoid sesamoid bones articulate with the palmar sides of the distal trochleae of each of the five metacarpal bones. The pollex has only two phalanges, whereas the other four digits contain three phalanges. Each digit has a curved claw bone covered by a sharp, curved claw (tegula). No distal sesamoid bones are present at the palmar sides of the distal interphalangeal joints.
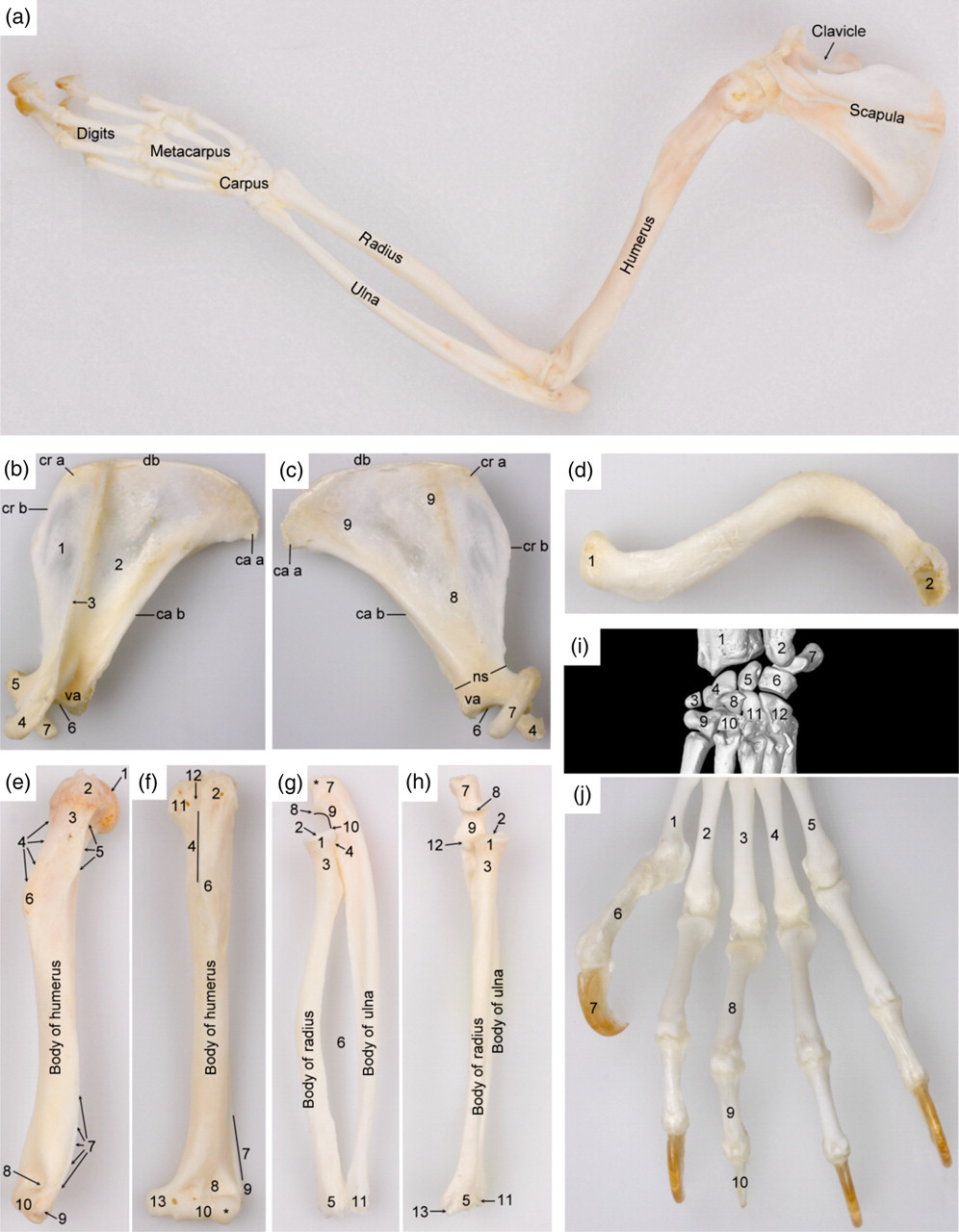
Skeleton of the left thoracic limb. (a) Topography of the skeletal structures of the thoracic limb (lateral view). (b) and (c) Lateral and medial views of the scapula, respectively: db = dorsal border (margo dorsalis); cr b = cranial border (margo cranialis); ca b = caudal border (margo caudalis); cr a = cranial angle (angulus cranialis); ca a = caudal angle (angulus caudalis); va = ventral angle (angulus ventralis); ns = neck of the scapula (collum scapulae); 1 = supraspinous fossa (fossa supraspinata); 2 = infraspinous fossa (fossa infraspinata); 3 = spine of scapula (spina scapulae); 4 = acromion; 5 = supraglenoid tuberosity (tuberculum supraglenoidale); 6 = glenoid cavity (cavitas glenoidalis); 7 = coracoid process (processus coracoideus); 8 = subscapular fossa (fossa subscapularis); 9 = serrate face (facies serrata). (d) Ventral view of the clavicle (clavicula): 1 = face articulating with the sternum (facies articularis sternalis); 2 = face articulating with the acromion of the scapula (facies articularis acromialis). (e) and (f) Lateral and cranial views of the humerus, respectively: 1 = head of the humerus (caput humeri); 2 = greater tubercle (tuberculum majus); 3 = neck of the humerus (collum humeri); 4 = crest of greater tubercle (crista tuberculi majoris); 5 = tricipital line (linea musculi tricipitis); 6 = deltoid tuberosity (tuberositas deltoidea); 7 = crest of the lateral epicondyle (crista supracondylaris lateralis); 8 = radial fossa (fossa radialis); 9 = lateral epicondyle (epicondylus lateralis); 10 = humeral trochlea (trochlea humeri) with capitulum (*); 11 = lesser tubercle (tuberculum minus); 12 = intertubercular groove (sulcus intertubercularis); 13 = medial epicondyle (epicondylus medialis). (g) and (h) Lateral and cranial views of the radius and ulna, respectively: 1 = head of the radius (caput radii); 2 = fovea capitis of the radius (fovea capitis radii); 3 = neck of the radius (collum radii); 4 = articular circumference (circumferentia articularis); 5 = trochlea of the radius (trochlea radii); 6 = interosseous space (spatium interosseum antebrachii); 7 = olecranon of the ulna with olecranon tuberosity (tuber olecrani(*)); 8 = anconeal process (processus anconeus); 9 = trochlear notch (incisura trochlearis); 10 = lateral coronoid process (processus coronoideus lateralis); 11 = lateral styloid process (processus styloideus lateralis); 12 = medial coronoid process (processus coronoideus medialis); 13 = medial styloid process (processus styloideus medialis). (i) Cranial view of the carpus: 1 = radius; 2 = ulna; 3 = sesamoid bone in the m. abductor pollicis longus (os sesamoideum m. abductoris digiti primi (pollicis) longi); 4 = radial carpal bone (os carpi radiale or os scaphoideum); 5 = intermediate carpal bone (os carpi intermedium or os lunatum); 6 = ulnar carpal bone (os carpi ulnare or os triquetrum); 7 = accessory carpal bone (os carpi accessorium or os pisiforme); 8 = central carpal bone (os carpi centrale); 9 = carpal bone I (os carpale primum or os trapezium); 10 = carpal bone II (os carpale secundum or os trapezoideum); 11 = carpal bone III (os carpale tertium or os capitatum); 12 = carpal bone IV (os carpale quartum or os hamatum). (j) Dorsal view of the left front foot: 1 = first metacarpal bone (os metacarpale primum); 2 = second metacarpal bone (os metacarpale secundum); 3 = third metacarpal bone (os metacarpale tertium); 4 = fourth metacarpal bone (os metacarpale quartum); 5 = fifth metacarpal bone (os metacarpale quintum); 6 = proximal phalanx of the first digit (phalanx proximalis digiti primi s. pollicis); 7 = distal phalanx of the first digit (phalanx distalis digiti primi s. pollicis) covered by a sharp, curved claw (tegula); 8 = proximal phalanx of the third digit (phalanx proximalis digiti tertii); 9 = middle phalanx of the third digit (phalanx media digiti tertii); 10 = distal phalanx of the third digit (phalanx distalis digiti tertii)
Hind limb
The hip bone (Figure 5) measures 40 ± 1 mm from the sacral to the ischial tuberosities (Table 1). No statistically significant difference between the male and female adult common marmosets was noticed. It contains a very large obturator foramen. The acetabulum is deep and contains a lunate articular face that is interrupted by an acetabular notch. Dorsal to the acetabulum, a discrete ischial spine is present. The pelvic symphysis remains syndesmotic in adult animals, and the angle between the left and right pubic bones is approximately 70° on craniocaudal view. Sexual dimorphism of the pelvis could not be observed.
The long bones of the hind limb (Figure 8) comprise the femur, tibia and fibula of which the lengths amount to 57 ± 2, 60 ± 2 and 58 ± 2 mm, respectively (Table 1). Statistically significant differences between the male and female adult common marmosets were not present. The femur is very slender and shows a well-delineated head, a distinct neck, and pronounced major and minor trochanters. No third trochanter or any obvious gluteal tuberosity is present. The patella has an ovoid shape. A small, round to cuboidal fabella articulates on the proximal surface of each femoral condyle. Like the radius and ulna, the tibia and fibula are separated by an interosseous space and are not fused. The tarsus contains seven tarsal bones, i.e. the talus and calcaneus in the crural row, the central tarsal bone (forming a reduced middle row) and four bones of the metatarsal row that increase in size from medial to lateral. Five digits are present. The hallux contains two phalanges whereas the other four digits have three phalanges. The flattened, blunt distal phalanx of the hallux is covered by a flat nail (unguis) whereas the claw bones of the other four digits are covered by sharp, curved claws (tegula). Axial and abaxial ovoid sesamoid bones are present at the plantar sides of the distal trochleae of the metatarsal bones. No distal sesamoid bones are present at the plantar sides of the distal interphalangeal joints.
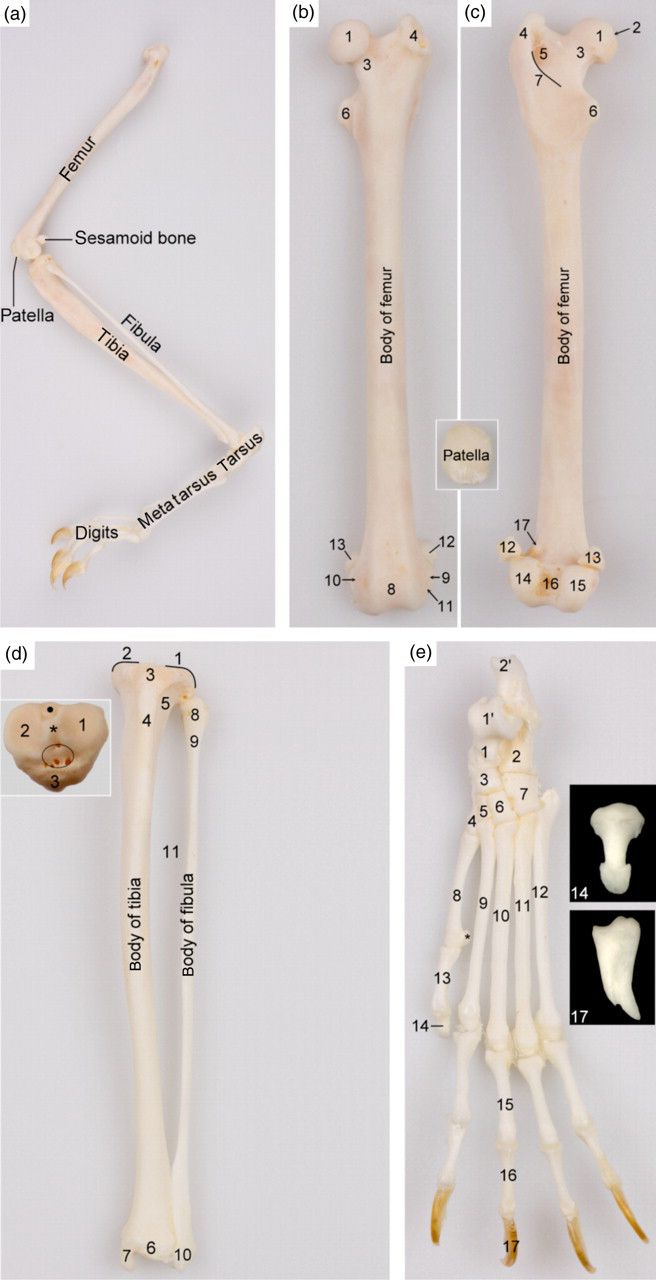
Skeletal structures of the left pelvic limb. (a) Topography of the skeletal structures of the left pelvic limb (lateral view). (b) and (c) Cranial and caudal views of the femur, respectively: 1 = femoral head (caput ossis femoris); 2 = fovea of the femoral head (fovea capitis ossis femoris); 3 = femoral neck (collum ossis femoris); 4 = greater trochanter (trochanter majus); 5 = trochanteric fossa (fossa trochanterica); 6 = lesser trochanter (trochanter minus); 7 = intertrochanteric crest (crista intertrochanterica); 8 = trochlea of the femur (trochlea ossis femoris); 9 = lateral epicondyle (epicondylus lateralis); 10 = medial epicondyle (epicondylus medialis); 11 = extensor fossa (fossa extensoria); 12 = lateral fabella (os sesamoideum musculi gastrocnemii laterale); 13 = medial fabella (os sesamoideum musculi gastrocnemii mediale); 14 = lateral condyle (condylus lateralis); 15 = medial condyle (condylus medialis); 16 = intercondylar fossa (fossa intercondylaris); 17 = lateral supracondylar tuberosity (tuberositas supracondylaris). The inset shows a cranial view of the patella. (d) Cranial view of the tibia and fibula: 1 = lateral condyle of tibia (condylus lateralis); 2 = medial condyle of tibia (condylus medialis); 3 = tibial tuberosity (tuberositas tibiae); 4 = tibial crest (crista tibiae); 5 = extensor groove (sulcus extensorius); 6 = tibial cochlea (cochlea tibiae); 7 = medial malleolus (malleolus medialis); 8 = fibular head (caput fibulae); 9 = fibular neck (collum fibulae); 10 = lateral malleolus (malleolus lateralis); 11 = interosseous space (spatium interosseum cruris). The inset shows a dorsal view of the tibial plateau: 1 = lateral condyle (condylus lateralis); 2 = medial condyle (condylus medialis); 3 = tibial tuberosity (tuberositas tibiae); * = intercondylar eminence (eminentia intercondylaris); encircled area = cranial intercondylar area (area intercondylaris cranialis); dot = caudal intercondylar area (area intercondylaris caudalis). (e) Dorsal view of the skeleton of the left hind foot: 1 = talus; 1′ = trochlea of talus (trochlea tali); 2 = calcaneus; 2′ = calcaneal tuberosity (tuber calcanei); 3 = central tarsal bone (os tarsi centrale or os naviculare); 4 = first tarsal bone (os tarsale primum or os cuneiforme mediale); 5 = second tarsal bone (os tarsale secundum or os cuneiforme intermedium); 6 = third tarsal bone (os tarsale tertium or os cuneiforme laterale); 7 = fourth tarsal bone (os tarsale quartum or os cuboideum); 8 = first metatarsal bone (os metatarsale primum); 9 = second metatarsal bone (os metatarsale secundum); 10 = third metatarsal bone (os metatarsale tertium); 11 = fourth metatarsal bone (os metatarsale quartum); 12 = fifth metatarsal bone (os metatarsale quintum); 13 = proximal phalanx of the first digit (phalanx proximalis digiti primi s. hallucis); 14 = distal phalanx of the first digit (phalanx distalis digiti primi s. hallucis); 15 = proximal phalanx of the third digit (phalanx proximalis digiti tertii); 16 = middle phalanx of the third digit (phalanx media digiti tertii); 17 = distal phalanx of the third digit (phalanx distalis digiti tertii)
No significant difference in bone sizes could be demonstrated between male and female common marmosets.
Discussion
Common marmosets are particularly useful as models in osteological studies since their bone morphology and physiology resemble that of man. In particular, the similarities between the aged marmoset and man in bone structure and remodelling are very valuable for the study of bone and joint diseases such as osteoporosis or rheumatoid arthritis. 1,6 The present morphometric data could, for example, be used as a standard in studies on growth-related disorders potentially evoked by calcium imbalance, or to assess the impact of therapies against osteoporotic skeletal deformation. Unfortunately, rodents cannot easily replace marmosets in such studies since they differ too much in bone characteristics. 1,2 Moreover, the Food and Drug Administration recommends the additional use of a non-rodent species in studies assessing bone quality. 1 Although marmosets are relatively small, they still require much more space compared with laboratory rodents. 7 Other disadvantages include the lack of diversified or genetically-altered strains and their higher demands concerning housing and feeding. 7 Although non-human primates such as the cynomolgus monkey (Macaca fascicularis) and rhesus monkey (Macaca mulatta) might be the best choice for translation of experimental data to man, their use is restricted by their high cost, human safety considerations, training requirements for personnel and limited availability. 1,12 It can therefore be concluded that the common marmoset presents a valuable research model in terms of cost versus benefit.
For osteological studies, it is important that the examined animals are healthy and do not suffer from skeletal diseases such as osteomalacia to which marmosets are very prone due to their impaired binding capacity and affinity of the receptor for the hormone 1α,25(OH)2D3. 7,26 This leads to vitamin D3 resistance which may enhance bone resorption to ensure adequate serum levels of calcium. 1 In addition, the potential reduction of both physical activity and ultraviolet exposure of common marmosets kept in captivity can result in enhanced bone resorption by osteoclasts. 1 Secondary hyperparathyroidism is responsible for deformation, bending or fracturing of the long bones influencing the results of osteological studies. 2 Until the age of 18 months, the animals investigated in the present study were housed in well-enriched facilities and had free access to outside pens in which they enjoyed much physical activity and direct sunlight. To prevent osteomalacia, they were supplemented with vitamin D3. In addition, blood analyses of the adult animals did not demonstrate any metabolic abnormalities.
The main characteristics of the skull of the adult marmoset include the almost complete postorbital closure of the eye sockets, the fused frontal suture and intermandibular symphysis which results in a single mandible, 27 the presence of a zygomaticoparietal suture, a large ectotympanic ring lateral to the tympanic bulla, short-tusked canines and three premolars in each oral quadrant. The presence of only two molars in each oral quadrant is rather atypical for Platyrrhini since most Platyrrhine species possess a third molar. 11,14
According to Wagner and Kirberger, 13 male marmosets often have 13 thoracic vertebrae, whereas females almost equally have 12 or 13. This could not be confirmed in the present study. On the contrary, only one of the seven examined females in our study had 12 thoracic vertebrae. Ongoing studies on more cadavers may clarify this ambiguity. However, the variation in the number of thoracic vertebrae is probably a mere anatomical variation that is also present in the rabbit that can either have 12 or 13 thoracic vertebrae. 28,29 In accordance with literature, 13 our observations confirm that the thoracolumbar vertebral number is always 19, that the last lumbar vertebra is consistently shorter than the other lumbar vertebrae, and that the sacrum consists mainly of three fused segments. However, transitional lumbosacral and sacrocaudal vertebrae have additionally been observed in the present study.
The arboreal life of marmosets is reflected in the very long and slender bones of the limbs, in particular the similar lengths of the stylopodia and zeugopodia of both the thoracic and pelvic limbs, and the tail size that is twice the length of the marmoset's body. Our measurements of the lengths of the long bones show slightly larger values compared with the morphometric data obtained in a previous study. 30 Different genetic background, health status, feeding and environmental enrichment can probably explain this difference. Leutenegger and Larson 30 state that Callithrix females surpass males in bone sizes by about 10%. The latter statement could not statistically be confirmed in the present study which might be due to the limited number of animals, in particular male animals, which were included.
The number and composition of the carpal bones in the marmoset contrast with that of hominoids, 28,31,32 but show similarities with that of the Old World monkeys and small domestic mammals including laboratory animals. A central carpal bone is found in all genera of Old World monkeys, 31 the rabbit, 28 the guineapig 33,34 and the mouse and rat. 35–37 This central carpal bone is a remainder of the primitive autopodium in which a central row of four bones is present. 38 A sesamoid bone in the m. abductor pollicis longus located at the medial side of the wrist is present in marmosets, cats and dogs. 16,39,40 Additional sesamoid bones are located at the femoral condyles and at the trochleae of the metapodial bones. These sesamoid bones are not common in hominoids. 28,31 In contrast to the carpus, the composition of the tarsal bones of C. jacchus is similar to that of hominoids, Old World monkeys, cats and dogs. 28,31,32,39,40
By presenting detailed anatomical data on the skeleton of the common marmoset, this study obviously promotes the education in laboratory animal science. Together with the morphometrical data provided in our study, more insight could be gained in pathological skeletal changes that are potentially induced by experimental protocols and drugs. Skeletal pathologies easily develop in the common marmoset that is characterized by a specific calcium metabolism. 20–23,26 In-depth knowledge of the skeleton of the common marmoset could also contribute to a better evaluation of the potential effect of therapeutic agents or specific diets administered to treat skeletal and arthrological disorders including osteoporosis, rickets and osteoarthritis. 5,6 This will probably result in refinement of research protocols and possibly also in a reduction of experimental animals.
Footnotes
ACKNOWLEDGEMENT
The authors thank P Vervaet for his kind help with the preparation of the skeletons.