
Abstract
The commonly used preclinical animal model of postmenopausal osteoporosis is the mature ovariectomized rat, whereby cessation of ovarian oestrogen production consequently results in bone volume reduction. The study aim was to precisely define the time course of structural changes resulting from ovariectomy and thereby reduce the time animals have to be treated to judge the effects of osteoporosis treatment. For this purpose, we assessed architectural changes by microcomputed tomography (μCT) during 10 weeks following ovariectomy or sham surgery at two-week intervals. Moreover, the trabecular microarchitecture of the lumbar vertebrae was assessed after necropsy. Besides this, serum biomarkers of bone turnover were determined. These data were in a new approach additionally correlated to femur mRNA expression profiles. We selected the osteoblast marker genes osteocalcin and type I collagen as well as the two osteoclast marker genes cathepsin k and tartrate-resistant acid phosphatase 5. The gene expression analysis suggested an activation of osteoblasts as well as octeoclasts. The significantly induced serum levels of osteocalcin and collagen degradation fragments also revealed this higher rate of bone turnover. Our results indicate that as soon as four weeks after ovariectomy the bone volume fraction exhibited a decline of 30% and 50% of the connectivity density. In addition, significant decreases of trabecular number and thickness as well as of the bone volume fraction were only observed in vertebrae of ovariectomized animals. Interestingly, changes of trabecular morphology were also found in the sham animals as a consequence of senescence.
A focal point of refinement in the use of laboratory animals is to determine the mechanism of interest and to permit that the earliest appropriate biological markers are identified and used. 1 This has especially great importance in slow-going pathophysiological transformation, e.g. osteoporosis. Osteoporosis, a systemic disorder of the skeleton, is characterized by the decline of bone mass and deterioration of the trabecular architecture that lead to an increased bone fragility. 2,3 Postmenopausal women are known to have a high incidence of osteoporotic fractures as a consequence of the cessation of the ovarian function and subsequently induced bone loss.
A frequently used small animal model to mimic the cancellous and endocortical bone loss similar to that found after human menopause is the adult ovariectomized (ovx) rat. 4,5 The bilateral removal of the ovaries in skeletally mature rats leads to an increased bone turnover rate with an imbalance in favour of bone resorption. 6 This model is accepted to be suitable in testing agents for prevention of osteoporosis or treatment as well as for preclinical evaluation of surgical treatment regimens of osteoporotic fractures. 7
Bone densitometry analyses in patients are mostly done by dual-energy X-ray absorptiometry (DXA) allowing a prediction of site-specific fracture risks by assessment of the bone mineral density (BMD). 8 In preclinical osteoporosis research, microcomputed tomography (μCT) allows for an even better determination of trabecular microarchitecture for defining bone quality. 9
The advantage of a longitudinal study design is that individual animals act as their own control as they are scanned several times. Therefore, the number of animals required can be reduced markedly, in accordance to the 3R principles of animal welfare.
The present study aimed at the analysis of the changes of the trabecular microarchitecture at the proximal tibia metaphysis after ovariectomy temporally in order to determine the time point when the bone shows clear osteoporotic features and to follow these changes. Furthermore, we investigated the impact of the isotropic voxel size on the morphological parameters obtained from the live animal scans by performing ex vivo μCT scans of the same bone (Acp5b) region at the end of the study.
Moreover, we analysed the femur mRNA expression profiles of osteocalcin (Oc), type I collagen as well as cathepsin k (Ctsk) and tartrate-resistant acid phosphatase 5 (Acp5b), genes that are used to monitor the activity of osteoblasts and osteoclasts, respectively. 10
Materials and methods
Animals
Pluriparous Wistar rats with an average age of 12 months (Janvier SA, Le Genest St Isle, France) were housed under controlled conditions (20 ± 1°C, 50–80% of relative humidity; 12:12 h light–dark cycles) in groups of four animals per cage with free access to water and a phytoestrogen-free diet (Harlan 2019 Rodent Breeding, Harlan Winkelmann, Borchen, Germany). The animals were subjected either to ovariectomy (ovx, n = 4) or to sham (n = 4) surgery. Anaesthesia was performed by intraperitoneal injection of 90 mg/kg body weight ketamine-hydrochloride (Ketamin 10%, Pharma Partner, Hamburg, Germany) and 10 mg/kg body weight xylazine (Xylariem®, Pharma Partner).
Animals were sacrificed using CO2 inhalation subsequent to a light O2/CO2 anaesthesia including blood collection during necropsy from the hepatic vein at 10 weeks postsurgery. In order to verify the hormonal status, the uterine wet weights were determined. Left and right tibiae as well as four lumbar vertebrae (LV3–LV6) were dissected and stored in 70% ethanol for ex vivo CT scanning. Left femora were cleaned from adjacent tissue and snap-frozen in liquid nitrogen.
The whole experimental set-up was planned and conducted to adhere closely to the 3R principles of animal welfare as well as to avoid enclosure of unnecessary additional experimental groups. Besides this all animal handling and all experimental conditions were ethically reviewed, licensed and carried out according to the Institutional Animal Care and Use Committee guidelines as regulated by the German Federal Law governing animal welfare.
In vivo microcomputed tomography and tibia analysis
In vivo μCT (vivaCT 75, SCANCO Medical, Brüttisellen, Switzerland) on the left proximal tibia of each animal was performed under anaesthesia at week 0 prior to surgery. Animals were anaesthetized as described above. Subsequently, two randomly selected animals of each group were subjected to in vivo imaging at two-week intervals up to week 8 postsurgery. The final μCT measurement of the excised tibiae and the four lumbar vertebrae in 70% ethanol took place (10 weeks postsurgery) after necropsy. The isotropic voxel size was 41 μm for in vivo scans (45 kV, 88 μA, 150 ms, 250 projections) and 20 μm for CT measurements of the excised tibiae and lumbar vertebrae, respectively (70 kV, 110 μA, 220 ms, 1000 projections). The estimated in toto radiation dose of the in vivo measurements was 400 mGy.
Calibration of the scanner took place weekly using hydroxyapatite phantoms.
The 1.4 mm thick region of interest on which the morphological analysis of the tibiae was performed was located 1 mm distal to the tibial growth plate. Contours were manually drawn adjacent to the endocortical boundary. For the analysis, Image Programming Language (IPL) software from SCANCO Medical was used.
The data-sets of the trabecular region were Gaussian filtered (Gauss support: 1; Gauss σ: 0.6) and globally thresholded. The threshold chosen for the data-sets obtained from the scans with the lower voxel size was 380, while 220 was chosen for the higher resolution.
Four lumbar vertebrae per animal were morphologically analysed. In the caudal half of the vertebrae, a region of interest (1.1 mm for LV6 and LV5, 0.78 mm for LV4 and 0.68 mm for LV3) 0.5 mm proximal to the growth plate was semi-automated contoured and a threshold of 230 was used.
Bone volume fraction (BV/TV), trabecular number (Tb.N), trabecular thickness (Tb.Th), trabecular separation (Tb.Sp) and connectivity density (Conn.D) were assessed for each time point. 9
RNA isolation, reverse transcription and realtime PCR
Sequences of primer pairs and amplicon sizes
1A: cytochrome-c-oxidase subunit 1A; Acp5b: acid phosphatase 5, tartrate resistant; Col1a1: collagen 1 a1; Ctsk: cathepsin k; Oc: osteocalcin; fwd: forward; rev: reverse
Quantification of bone markers
At the time of necropsy, blood samples were collected and serum was subsequently separated and stored at −20°C. Enzyme immunoassays (EIAs) were performed to determine the serum concentration of the bone formation marker osteocalcin (Rat-MID™ osteocalcin EIA, Immuno Diagnostic Systems GmbH, Frankfurt am Main, Germany) and degradation fragments of type I collagen (RatLaps™ EIA, Immuno Diagnostic Systems GmbH) to assess bone resorption.
Statistical analysis
The data are presented as arithmetic means ± standard deviation. Statistical analysis included one-way analysis of variance (ANOVA) followed by Bonferroni post hoc test in order to determine significant differences. Paired t-test was used to test for changes in body weight relative to the baseline. Statistical significance of results was considered at P ≤ 0.05.
Results
Body and uterus weights
As shown in Figure 1a, ovariectomy induced hormone ablation led to a decrease of the relative uterine wet weight being 4.5-fold lower compared with the wet weight in the sham-operated group. At the time of surgery, the mean body weights of the sham and the ovx groups were not significantly different (376.0 ± 33.8 and 374.3 ± 30.2 g, n = 4). Weekly measurements following surgery showed a significant increase in the ovx group up to 16% at the end of the study compared with baseline (Figure 1b). Ten weeks postsurgery, the mean mass of the sham-operated animals did not differ significantly from the baseline (375.8 ± 39.9 g).
(a) Relative uterine wet weights (UWW) of Wistar rats 10 weeks after bilateral ovariectomy (n = 4). Box plots show the 10% and 90% percentiles, the median (+) and the minimum and maximum values. +
P< 0.05 denotes statistical significance, calculated by one-way analysis of variance (ANOVA) followed by Bonferroni post hoc test. (b) Percent change of the group average body weight relative to the baseline over a period of 10 weeks (n = 4). *P< 0.05 denotes the first time point when body weight was significantly different from the baseline (paired t-test). +
P< 0.05 represents a statistical significance with respect to the corresponding value of the sham group, calculated by one-way ANOVA followed by Bonferroni post hoc test. ovx: ovariectomized group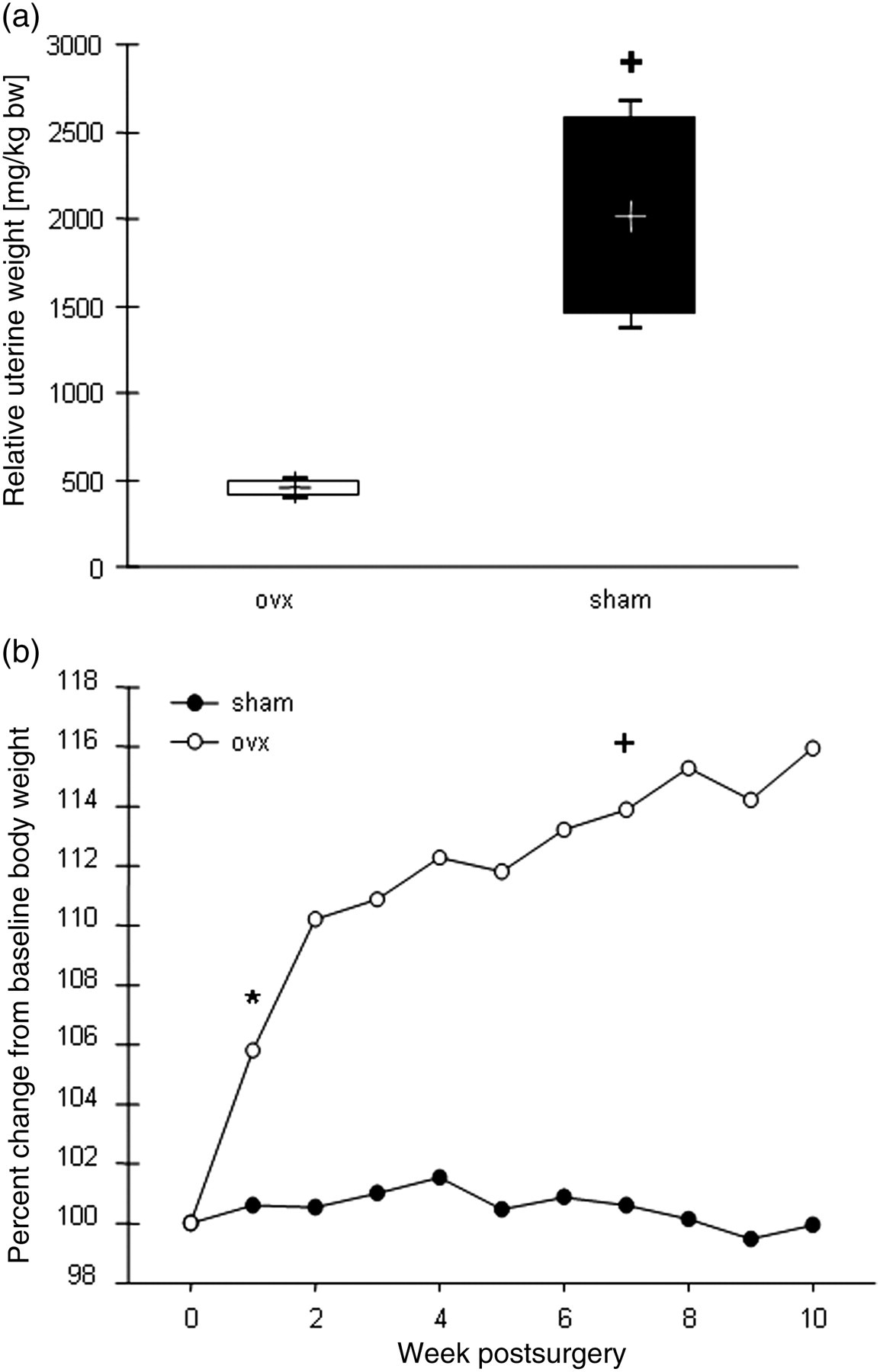
Microarchitectural bone parameters
We investigated the changes of the trabecular micro-architecture in ovx- and sham-operated rats during 10 weeks postsurgery. The average percent change from baseline (100%) of each morphological parameter is shown in Figure 2. In the sham-operated animals, BV/TV as well as Tb.Th and Tb.Sp decreased up to 80% from the baseline at week 10 postsurgery (Figure 2a). By contrast, Tb.N increased slightly. Conn.D increased up to 150% 10 weeks postsurgery.
Percent changes of microarchitectural parameters in the proximal tibia relative to the baseline (100% = week 0) (BV/TV: bone volume fraction; Conn.D: connectivity density; Tb.Sp: trabecular separation; Tb.N: trabecular number and Tb.Th: trabecular thickness). (a) Sham-operated group. (b) Ovariectomized group. Values are the mean of two animals (week 2 up to week 8) or four animals (week 0 and week 10) respectively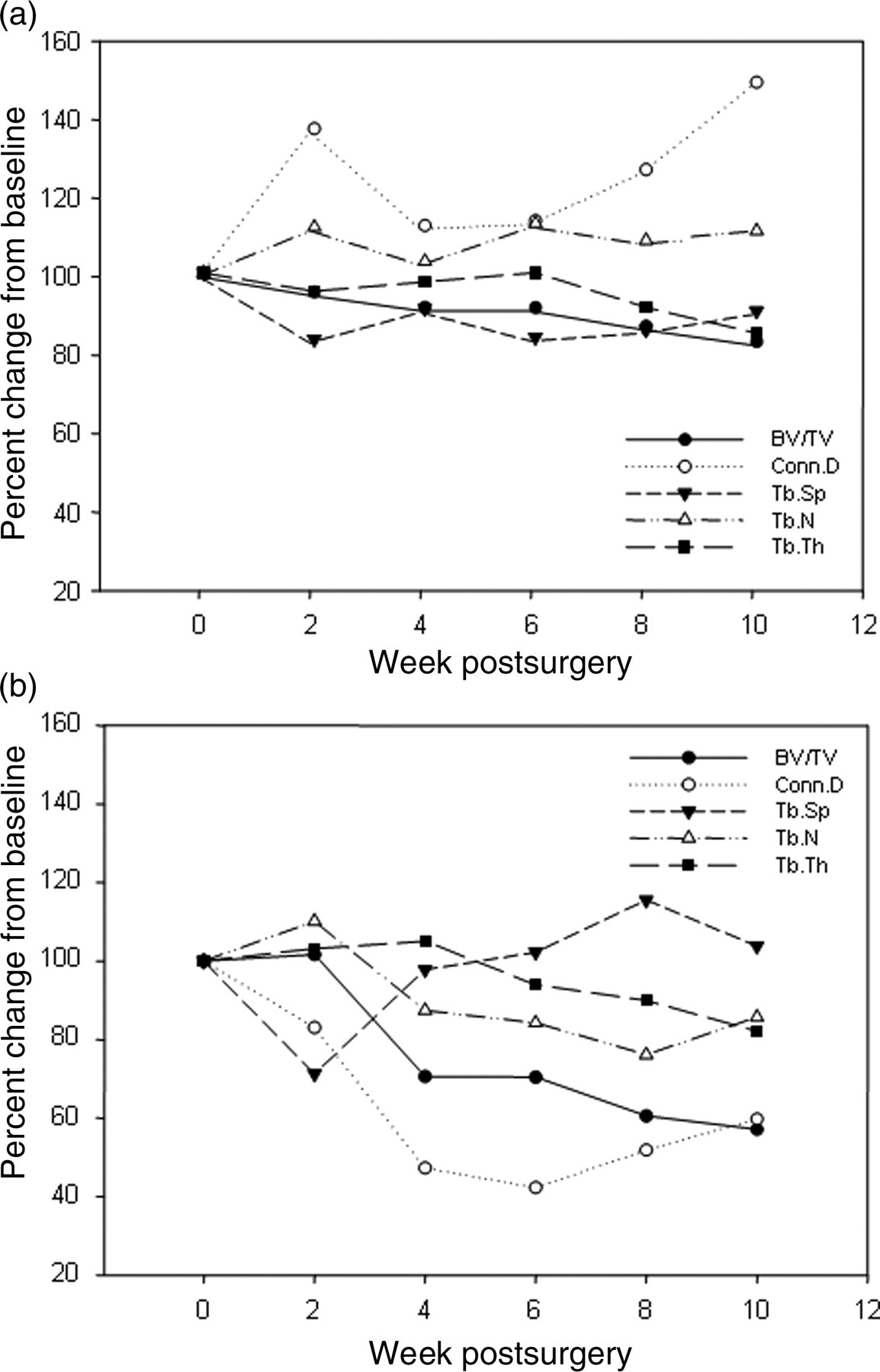
The depletion of endogenous oestrogens led to clear changes of the bone parameters as shown by a 44% decrease of BV/TV and Conn.D up to week 10 (Figure 2b). The comparison of the bone parameters obtained from the animals that were irradiated multiple times and the animals that were scanned only once at the time of the surgery showed no significant differences in bone parameters (data not shown).
Furthermore, we scanned the excised tibiae using a low (41 μm) and a high isotropic resolution (20 μm). Tibiae of both hind limbs of each animal were included in the measurement giving an internal control of non-irradiated samples. μCT images from the two isotropic voxel sizes used are comparatively shown in Figure 3b. The ovx-induced bone loss 10 weeks after castration is exemplary shown in Figure 3a. No significant differences could be observed for Tb.N and Tb.Sp due to the chosen isotropic voxel size (n = 4, Table 2). Moreover, the outcome for BV/TV was not significantly different except for the left tibia in the ovx group. By contrast, the result of the bone parameter Conn.D seemed to be highly dependent on the isotropic voxel size as the values were significantly different.
(a) Trabecular microarchitecture of one animal at the time of ovariectomy (week 0, left-hand) and 10 weeks postovariectomy. Image acquisition was performed using an isotropic voxel size of 41 μm. (b) μCT scans of the proximal tibia of one sham-operated rat resulting from an isotropic resolution of 41 μm (left-hand) and 20 μm (right-hand) Morphological parameters resulting from ex vivo scans with two different isotropic voxel sizes (41 and 20 μm) Data of each hind limb are shown as group means ± SD (n = 4). P values are reported for the one-way analysis of variance representing significant differences of the 20 μm measurement from the 41 μm scan in square brackets and significant differences between the ovx and the sham group-in curly brackets, *+P< 0.05 BV/TV: bone volume fraction; Conn.D: connectivity density; Tb.Sp: trabecular separation; Tb.N: trabecular number; Tb.Th: trabecular thickness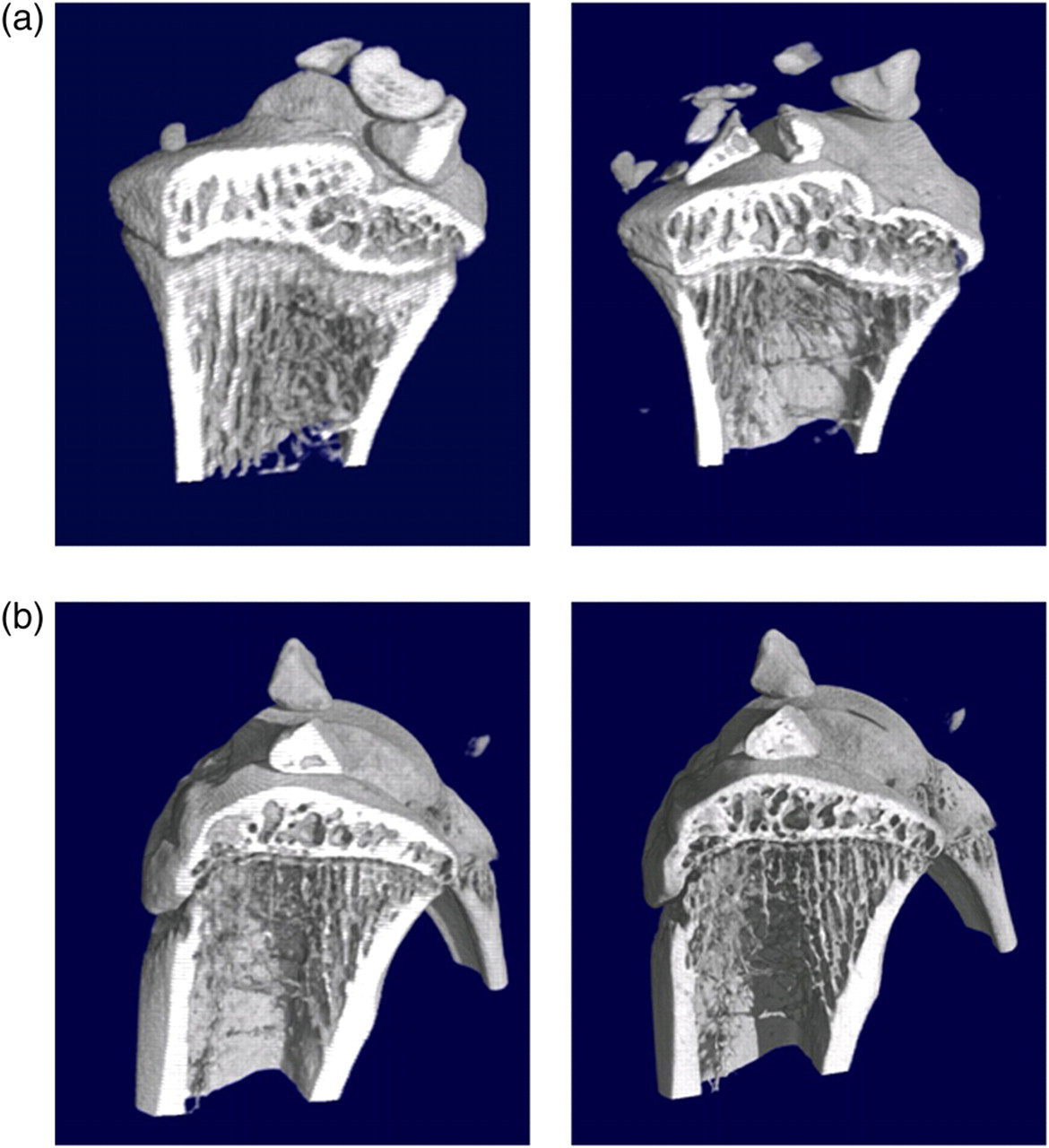
Morphological parameters resulting from μCT scans of the dissected spine (isotropic voxel size: 20 μm)
Data are shown as group means of four animals and four lumbar vertebrae (LV3, LV4, LV5 and LV6) (mean ± SD, n = 16). Statistically significant differences between ovx- and sham-operated animals were tested with one-way analysis of variance, P values are shown in square brackets, *P< 0.05
BV/TV: bone volume fraction; Conn.D: connectivity density; Tb.Sp: trabecular separation; Tb.N: trabecular number; Tb.Th: trabecular thickness
Serum marker
The serum levels of one biochemical marker for bone formation (osteocalcin) and resorption (collagen fragments) have been studied and are shown in Figure 4. In comparison to the sham-operated animals, the concentrations of osteocalcin and of the collagen fragments were significantly elevated in castrated animals at the time of necropsy (1.8- and 1.5-fold respectively, n = 4).
Serum levels of type I collagen degradation fragments (RatLaps™) and osteocalcin 10 weeks after sham surgery and ovariectomy, respectively. Values are expressed as mean ± SD (n = 4). Statistical significance was tested by one-way analysis of variance and Bonferroni post hoc test. + denotes significant differences from sham group, P < 0.05 ovx: ovariectomized group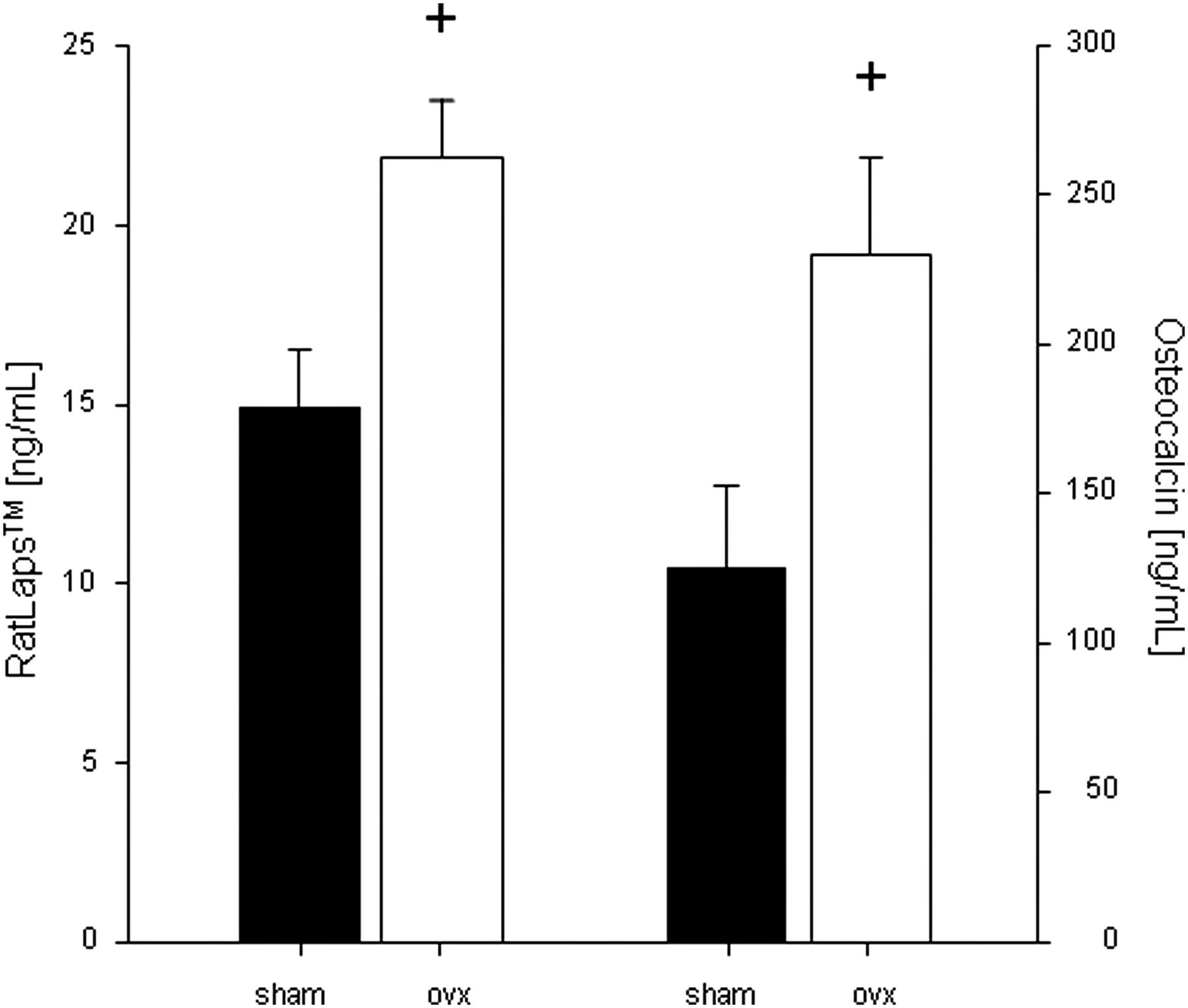
Gene expression
We were interested in the expression pattern of osteoblast and osteoclast marker genes in the femora. The expression levels of Oc and type I collagen (Col1a1), both markers for osteoblast activity in the ovx animals were increased compared with the sham-operated animals (Figures 5a and b). The mean Col1a1 mRNA level of the ovx group was significantly increased though we only detected a not-statistically significant trend to an increased Oc expression level (n = 4). The femoral expression of Ctsk and Acp5b as marker genes for osteoclast activity was slightly higher in the ovx animals (Figures 5c and d). The average Acp5b but not Ctsk mRNA expression level was found to be significantly higher compared with the sham-operated group (n = 4). The mRNA levels of all four genes investigated showed high inter-individual variability within one group, e.g. as the Oc expression among the sham-operated animals differed by a factor of two.
mRNA expression levels of osteocalcin (a), collagen 1a1 (b), cathepsin k (c) and acid phosphatase 5, tartrate resistant (d) relative to the expression of the housekeeping gene 1A. Black and striped bars correspond each with a single animal, white bars are the average mean of one group, respectively (n = 4). Statistical significance was tested by Student's t-test, +
P < 0.05 indicates statistically significant differences between ovariectomized (ovx) and sham groups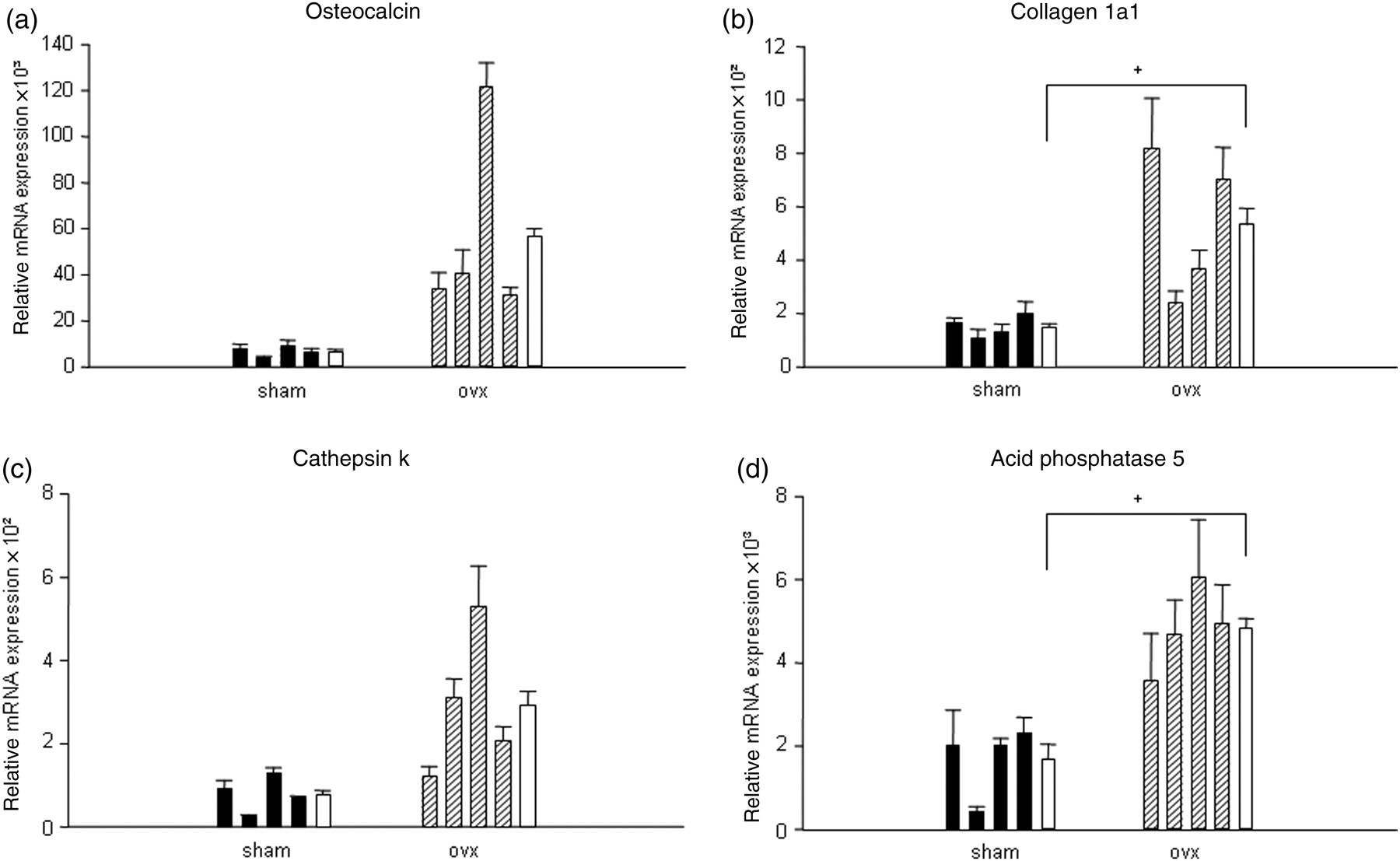
Discussion
The present study aimed at the combination of a temporal analysis of changes in the trabecular microarchitecture at the proximal tibia metaphysis of adult rats after ovariectomy using in vivo μCT together with an ex vivo analysis of both (i) details of osteoporotic structural features in the tibiae and spine and (ii) serum as well as mRNA expression profiles of typical markers for osteoblastic and osteoclastic activities.
The depletion of endogenous oestrogens after bilateral ovariectomy resulted in hyperphagia and a subsequent increase in body weight (Figure 1b). 11,12
Although the possibility of a partial prevention from ovx-induced osteopaenia due to weight gain has been described, 12 we could see clear changes in the trabecular microarchitecture compared with the sham animals (Figure 2a and b). Our findings are in accordance with recent studies where significant effects of oestrogen deficiency have been shown in ovx groups that were also not weight matched to the control group. 13,14
Despite the negative correlation of increased fat mass and fracture risk shown in epidemiological studies, 15 it is unlikely that the moderate ovx-induced weight gain affects the bone microarchitecture in mature rats that had already reached their peak bone mass before surgery. 16
We selected the proximal tibial metaphysis and the lumbar spine as the two skeletal sites for evaluation of morphological changes because they are often used in preclinical research for this purpose. 5,14,17 Moreover, it has been described only recently that they belong to the skeletal sites in older Caucasian women where fractures are likely to occur as a result of osteoporosis. 18
The analysis of the tibial trabecular bone parameters of the sham animals showed a 9% decrease of the BV/TV compared with baseline during the six weeks after surgery. Up to 10 weeks after sham-surgery, BV/TV showed a significant change up to 82% (Figure 2a). A further significant decrease was observed with Tb.Th. Even though these differences compared with baseline were significant, the structural alterations seen in the oestrogen-depleted animals were even more pronounced (Figure 2b). A potential reason for the changes observed in the sham-operated animals could be related to the age of the rats. It has been described that the rodent skeleton undergoes ageing that leads to a decrease in BMD 19–21 after having reached the peak bone mass at around the age of 10 months. 5
However, the structural changes seen in the ovx animals occurred much faster, as a half maximal (29%) decrease of the BV/TV was already observed four weeks after ovariectomy that exacerbated to 60% at the end of the study. Conn.D is the morphological parameter that shows the most pronounced changes as it was already reduced to 50% of baseline four weeks after ovariectomy. This finding is in accordance with others who described early alterations of trabecular connectivity after ovariectomy. 14,22 Reductions in connectivity of the trabecular network can be seen as irreversible changes of the bone microarchitecture, 23 therefore intervention studies aiming at the prevention of bone loss should start immediately after oestrogen depletion. 14
In contrast to previous studies that reported either no significant effect or a decrease of Conn.D in the proximal tibia of sham animals, 14,24 we observed an increase in Conn.D and Tb.N during the period of investigation. These differing observations can be explained by the parameters used during image processing which have a high impact on the outcome of the bone morphometry.
In the current study, Tb.Th in the proximal tibia decreased significantly from baseline in the ovx group whereas the trabeculae in the ovx group were thinner compared with the sham counterparts at the end of the study. In matters of ovx-induced effects on Tb.Th, there are inconsistent data in the reports. Some authors report oestrogen deficiency leading to trabecular thickening while others describe thinning of Tb.Th in the tibia. 14,17,25 Although we also observed a significant decrease of Tb.Th in the sham animals, the values in the ovx group were significantly different compared with the control group at the time of the last scan 10 weeks postsurgery.
In the lumbar vertebrae, we observed a significant thinning of the trabeculae 10 weeks after castration (Table 3). In combination with a decreased Tb.N and an increased distance between the trabeculae, this gave rise to a significantly lower BV/TV.
In contrast to the metaphysis of the tibia, the thinning of trabeculae in the lumbar spine as a response to ovariectomy is consistently described in literature. 17,26,27 Another site-specific response was found in terms of Conn.D after E2 deprivation that was in contrast to the tibia which was significantly increased in the lumbar spine. This observation is in accordance with the findings of Kinney et al. who described a 150% increase in connectivity 120 days after ovariectomy. 27 The reason for this site-specific effect was explained by the more plate-like structure of the rat vertebra versus the more rod-like one in the tibia, giving rise to a differing response.
The comparison of the values obtained from the endpoint measurements using two different isotropic voxel sizes showed, independently from the systemic status (sham-operated or ovx), no significant differences (Table 2). Hence, we could show for the first time that the resolution had no influence on the results for BV/TV, Tb.N and Tb.Sp underlining that the outcome obtained by the in vivo scans with anesthetized animals at a lower-scan resolution is meaningful. This is an important finding as the spatial resolution applied in live animal analysis is limited by the total radiation dose per animal during each scan. 28 Thus, the isotropic resolution applied in longitudinal study designs where individual animals are scanned several times acting as their own control is restricted by the number of scans and the ionizing radiation to which the animal is exposed during one scan.
By contrast, the data for Conn.D and Tb.Th varied significantly in that the lower isotropic resolution resulted in lower values of Tb.Th and higher values of Conn.D.
As expected, the serum markers of bone homeostasis were significantly increased 10 weeks postovariectomy in comparison with the sham-operated animals clearly indicating a higher bone turnover rate (Figures 4a and b). This can be correlated to the mRNA level as the gene expression analysis also revealed elevated mRNA levels of osteoblast-specific marker genes Oc and Col1a1. Additionally, the gene expression of osteoclast marker genes, Ctsk and Acp5b, was increased in the ovx animals. 10 The relative mRNA expression levels have been investigated in each single animal showing high inter-individual differences within one group. This high variability could also be observed in terms of body weight, biochemical serum markers and morphological parameters, probably as a consequence of the usage of pluriparous animals which results in an increase in physiological variation. The high variability could be partially described by the differences in breeding cycles and gestation frequencies. By using an inbred rather than an outbred rat strain, another cause for intra-individual variation could possibly be reduced and thereby the discrepancy found in the current study. Besides this, age-matched nulliparous and pluriparous animals should be compared in future studies in order to further decrease intra-individual variation.
In conclusion, we could demonstrate that E2 deprivation in mature rats induced changes of the tibial micro-architecture resolvable as soon as four weeks after ovariectomy. These morphological alterations included a thinning of the trabeculae, an increase in Tb.Sp and a decline in BV/TV.
As another novelty, we showed that the isotropic resolutions we compared (41 and 20 μm) did not significantly influence the outcome of BV/TV, Tb.N and Tb.Sp.
In addition, significant bone loss in terms of decreased Tb.N, Tb.Th, Tb.Sp and BV/TV was observed 10 weeks after ovariectomy in the spine. These changes of the bone microarchitecture could be correlated to biochemical serum markers of bone homoeostasis as well as to an enhanced mRNA expression of genes used to monitor bone cell activity that indicate an induced bone resorption due to ovariectomy.
In using μCT with a resolution comparable with that used in this longitudinal study (41 μm), our results emphasize that the onset of intervention is crucial. In study designs testing potential bone loss preventing agents, we recommend starting the intervention at the time of E2 depletion as irreversible changes were found to take place just following ovariectomy. A further advantage of this longitudinal study design is in terms of animal welfare, i.e. refinement in which each individual acts as its own control. Moreover, an intervention period of a maximum of eight weeks can be seen as sufficient to see bone sparing effects. Additionally this reduction of time results in a decline of the resources needed.
Footnotes
ACKNOWLEDGEMENTS
The authors thank Iris Käppler for her assistance during the CT scans. This work has been supported by Deutsche Forschungsgemeinschaft DFG TRR67.