
Abstract
Bedding influences various parameters in the housing of laboratory mice, such as health, physiology and behaviour (often considered as being integral parts of welfare). Notwithstanding existent studies about bedding preferences of individually tested mice, data about group-housed mice are still lacking. The aim of this study was to find out the structure preference for softwood bedding of group-housed mice. One hundred and eight 8-week-old female mice (C57BL6/JOlaHsd and BALB/cOlaHsd) were housed in groups of three and were given one-week free access to two different bedding structures at a time. In three test combinations, softwood shaving bedding was tested versus softwood chip bedding products of three different particle sizes (fine/medium/coarse-grained). The preference test was performed in a DoubleCage system composed of two Makrolon type IIL cages, connected by a perspex tunnel. This validated system was able to detect the crossings of each individual animal with correct crossing time and direction. On the basis of these data, dwelling times on the particular bedding structures were statistically analysed as a parameter for bedding preferences. In all three test combinations, a highly significant shaving preference was detected. On average, mice spent 70% of their dwelling time on the shavings. This preference was more explicit during the light period and in C57BL/6J mice. The relative ranking of the bedding structures was: shavings≫coarse-grained chips>medium chips = fine chips. By means of these results, a shaving structure as bedding can be recommended for laboratory mice, whereas fine chip structures should be avoided.
In standard housing of laboratory mice and rats, bedding and nesting material are the only substrates animals are in direct and permanent contact with. Thus, bedding can influence all aspects of animal housing and experiments, including occupational safety, 1,2 animals' health 3–7 and behaviour, 8–11 as well as physiology 12–19 and pathology. 12,20,21 Based on this knowledge, a bedding product should be dust-free, absorbent, free of contaminants and pathogens, free of toxic or carcinogenic substances, inedible, indigestible, free of enzyme inducers, not interacting with experimental parameters, soft, atraumatic, insulating, manipulable, adequate for nest-building, autoclavable, storable, easily removable from cages and cheap. 19,21–24
To gain more information about animals' needs, preference tests are a conventional and suitable method and therefore can help to improve housing conditions. 25 Various preference tests have been investigated for laying hens, 26,27 dairy cows, 28 pigs 29 or rats. 30 For laboratory mice, preferences for different caging types, 31 nesting materials 32,33 and environmental temperatures 34 have been studied among others. Regarding bedding preferences, it was found that mice and rats prefer soft, long, large particles. 32,35–38 Furthermore, mice prefer wood bedding to other bedding materials, 39,40 and aspen wood to other types of wood bedding. 9,36,40 However, all animals were individually tested. As mice are highly social animals which should be kept in groups whenever possible, it is important to know which bedding mice prefer when they are group-housed. 41,42
In contrast to the past, 41,43 it is now possible to individually register socially-housed animals during preference tests. According to the validation of this new system (DoubleCage, described in the Animals, materials and methods section) the data registered by an automated system and by manual observation (video analysis) are highly correlated (R 2 = 0.997 and 0.999 for dwelling times in both cages and R 2 = 1.000 for crossing frequency). 44
Furthermore, none of these previous studies clearly distinguished the material from the structural aspect, thus the present study focused on structure-related bedding preferences of group-housed mice using the DoubleCage system. The aim was to find out what kind of bedding structure is recommendable for group-housed mice.
When reviewing all publications in the journal Laboratory Animals in 2010, we found that most EU authors (around 90%) provided wood bedding for rodents. Thus, wood is still the bedding material most commonly used in Europe. The presented data focused first on four softwood products. To avoid possible aggressive interactions, 45 only female mice were tested and other factors potentially affecting outcome, such as the reaction to new environment, test duration, previous experience, cage position, 41 were also considered in the experimental plan.
Animals, materials and methods
A total of 108 (54 BALB/cOlaHsd and 54 C57BL/6JOlaHsd) female specific-pathogen-free (SPF) mice were used. According to the breeder (Harlan, Horst, The Netherlands), animals were raised on cellulose flake bedding (Harlan 7099 TEK-Fresh) with a round PVC tube as enrichment. At the age of three weeks, they were supplied by van in filter boxes. Preference tests were performed at the age of eight weeks (average weight: C57BL/6JOlaHsd: 17.4 ± 2.5 g; BALB/cOlaHsd: 20.2 ± 2.6 g). The animals' SPF status was assured by the breeder's certificate and health monitoring according to current FELASA recommendations 46 at the end of the experiment. After arrival the mice were randomly allotted to stable groups of three, which remained for the entire experimental period.
Macroenvironment
All animals were housed in conventional rooms under positive pressure, at a room temperature of 22 ± 2°C and 55 ± 10% relative humidity with air exchange 10–16 times per hour, on a 12/12 h light/dark cycle (light on at 06:00 h, light off at 18:00 h) and at a light intensity of 61 ± 10 Lux (measured at cage level in the acclimatization room) or 65 ± 5 Lux (measured by a DoubleCage-sensor, see Test system section).
Husbandry
Animals were kept in open cages (Makrolon type II long, 32.5 × 16.5 × 14 cm, Tecniplast, Hohenpeißenberg, Germany), food (10 mm pelleted diet, Altromin No. 1324; Altromin, Lage, Germany) and tap water (in bottles, drinking quality) were provided ad libitum (identical in all cages). Bedding (1.2 L per cage = depth 2 cm) was given according to the particular test combination (see Procedure and experimental design section). Bedding and water were changed weekly.
Acclimatization
During the first three weeks of acclimatization, animal groups were housed in type IIL cages, followed by two weeks adaptation to the DoubleCage test system. During the five weeks of acclimatization, half of the groups received shavings as bedding and half were kept on chips (fine, medium or coarse, depending on the respective test combination as described in Figure 1).
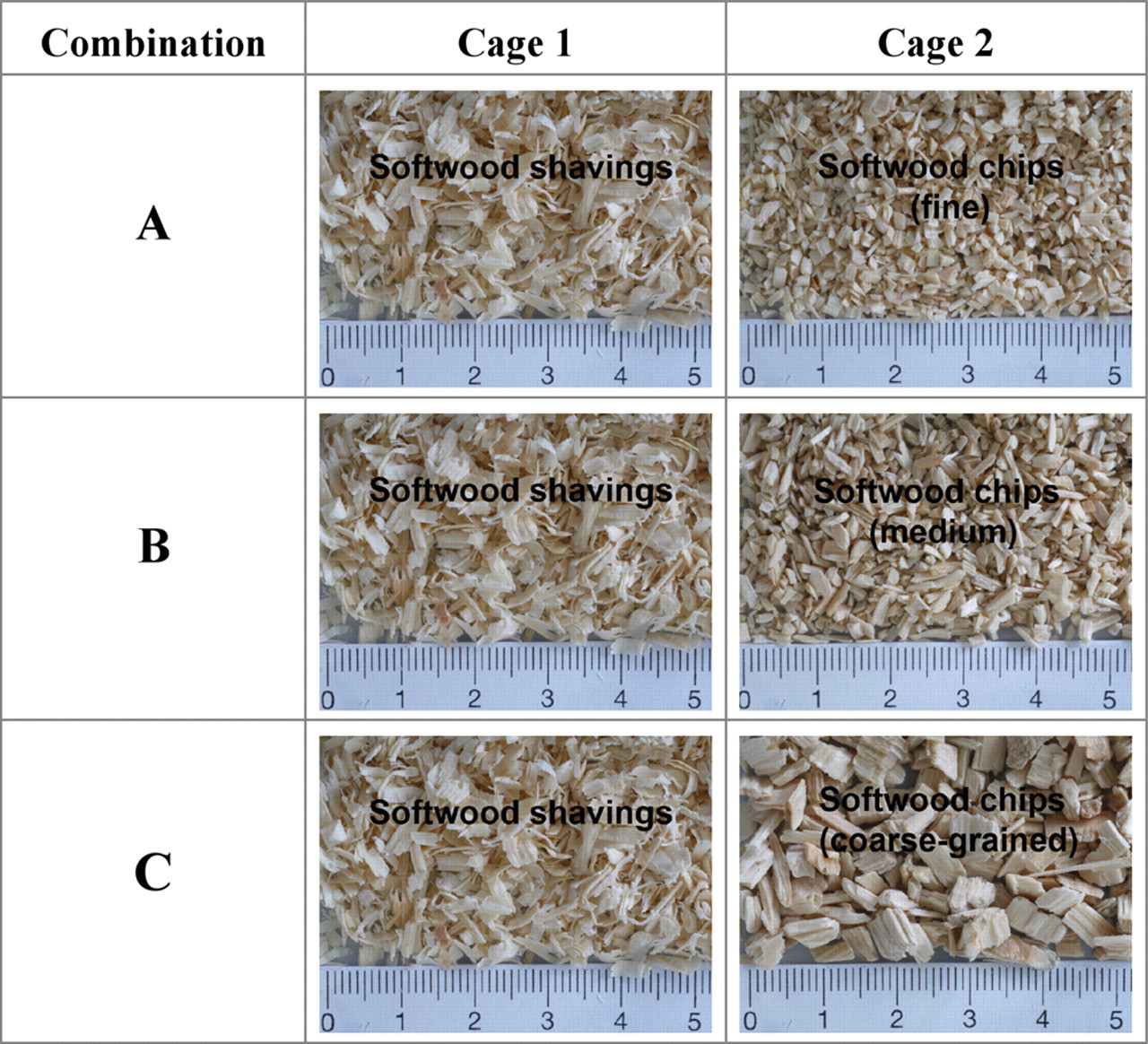
Bedding types in test combinations A–C, softwood shaving bedding was tested against three different chip beddings
Transponders
At six weeks of age, each animal obtained a subcutaneous transponder (Euro ID Identification Systems; Weilerswist, Germany) (short isoflurane anaesthesia), so that each animal could be individually identified.
Test system
The system (DoubleCage) was designed in cooperation with the University of Zurich 44 to meet the requirements of our institute. As described above, the DoubleCage system (Figure 2) was able to identify crossings of individual animals (up to 16 animals, 4 animals per cage) and to record realtime data. Each DoubleCage was composed of two Makrolon cages (type IIL), cage 1 and cage 2, connected by a perspex tunnel (length: 30 cm, inner ∅: 2.4 cm), which had a transponder sensor on either side and was connected to a personal computer.

DoubleCage test unit: cage 1 and cage 2, perspex tunnel with two black sensors
Mice were allowed to move freely between cages 1 and 2 and could be detected individually by the sensors. Each crossing and crossing direction were registered automatically. Crossing frequency and dwelling times of each animal in both cages could be analysed.
Procedure and experimental design
After arrival, mice were randomly allotted to stable groups of three, followed by five weeks of acclimatization. Cages were again randomly assigned into three experimental groups for different test combinations (A–C, Figure 1), six cages per strain for each combination. Preference tests were performed at the age of eight weeks for one week.
The position of cages in the rack was rotated. Food and water intake during the preference test were measured by weighing directly before and after the test.
Statistical analysis
From each preference test week, data of the first day (day 0) were omitted. Data of days 1–6 were analysed by using StatView computer program (Version 5.0, 1998, SAS Institute Inc, Cary, NC, USA). Differences between the dwelling times in cage 1 (time on shavings) and cage 2 (time on chips) were compared by the paired t-test. The differences between strains or between the test combinations were analysed using analysis of variance (ANOVA) test with a significance level of 0.05, followed by Scheffé test. To analyse the correlation between dwelling time and food/water consumption, the dwelling time of each cage was calculated. The data of all cages were pooled together for analysis (n = number of cage).
Results
Total dwelling times
In each test combination, shavings were clearly preferred with a high significance (Figure 3). On average, animals spent about 70% of their dwelling time on the shavings: 71.87 ± 11.51% in combination A (shavings versus fine chips), 71.67 ± 13.77% in B (shavings versus medium chips) and 65.27 ± 13.80% in C (shavings versus coarse-grained chips). Regarding the dwelling time on chips, there was a significant difference between the three test combinations (ANOVA F 2,95 = 4.223, P = 0.0175). Animals spent significantly more time on coarse-grained chips than on medium chips (Scheffé test P = 0.0349) and fine (Scheffé test P = 0.0445) chips. Therefore, the relative ranking of the four structures is as follows: shavings≫coarse-grained chips>medium chips = fine chips.
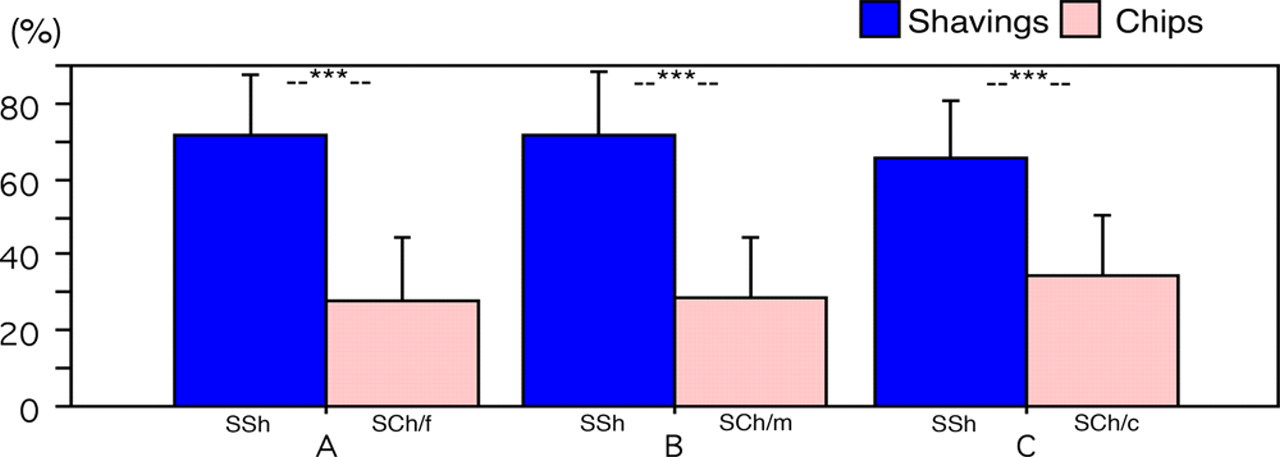
Dwelling times (% per day, mean ± SD) on shavings and chips, respectively, in test combinations A–C. S: softwood; Sh: shavings; Ch: chips; f: fine; m: medium; c: coarse-grained. ***P < 0.0001 (paired t-test, n = 36)
Light/dark period
In both light and dark periods, mice preferred the shavings in all test combinations (Figure 4). This preference was more pronounced in the light period than in the dark period (paired t-test P < 0.0001). Chips were occupied more during the dark period, but nevertheless shavings were always clearly preferred (P < 0.0001 for all combinations).
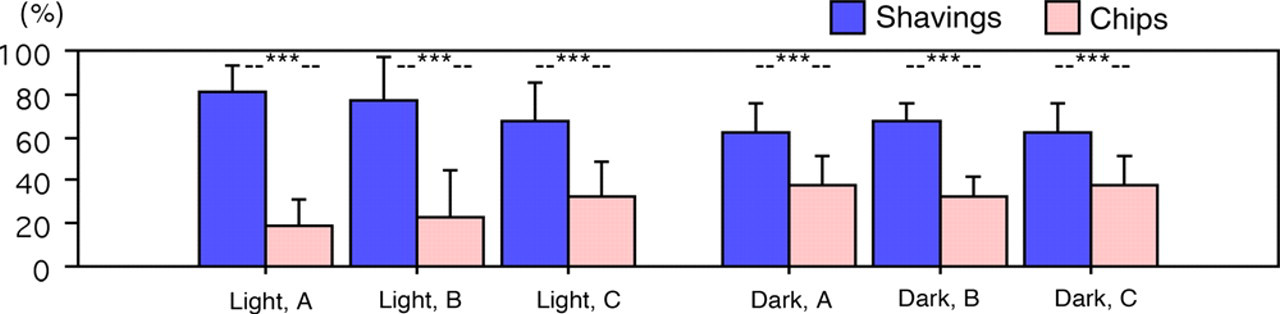
Light (left) and dark (right) periods: dwelling times (% per period, mean ± SD) on shavings and chips, respectively. ***P < 0.0001 (paired t-test, n = 36)
Strains
Both strains showed a significant preference for the shavings (Figure 5). In BALB/c mice, it was slightly lower but also explicit (P < 0.0001 for A, P = 0.0029 for B, P = 0.0079 for C), while in the case of C57BL/6J mice this preference was always highly significant (P < 0.0001 for all combinations). In contrast to C57BL/6J mice, BALB/c mice did not spend significantly more time on the coarse-grained chips than the finer chips.
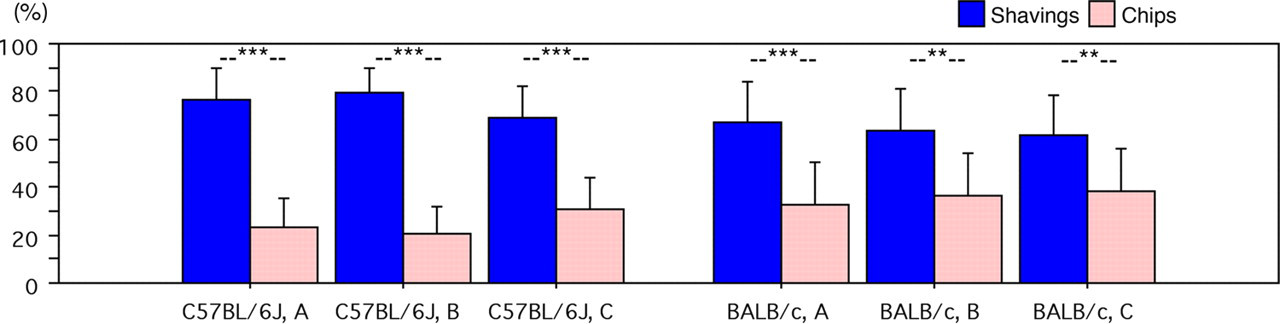
C57BL/6J (left) and BALB/c (right): dwelling times (% per day, mean ± SD) on shavings and chips, respectively. **P < 0.01, ***P < 0.0001 (paired t-test, n = 18)
Weekly course
In both combinations A and B, the shaving preference was present from the first day onwards and remained stable the entire test week (Figure 6), without any significant differences between the days. This applied to both strains. Combination C started with a shaving preference of 60% dwelling time and increased up to 75% on day 6.
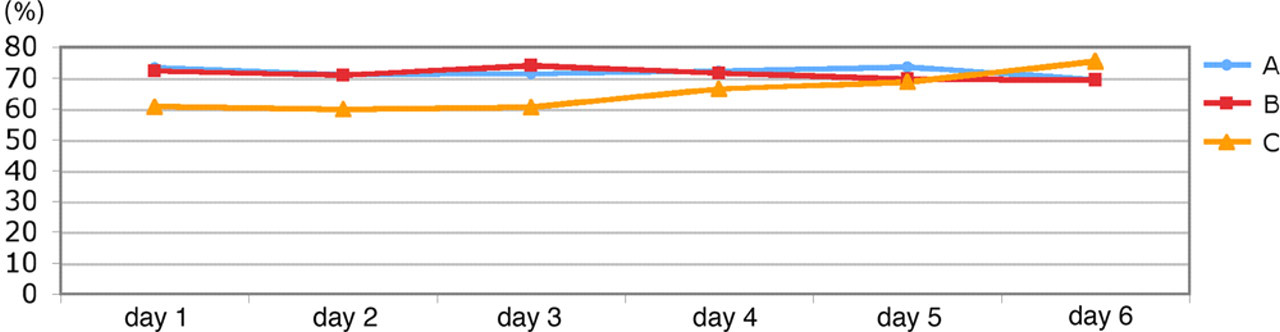
Weekly gradient: dwelling times (% per day, mean) on shavings (days 1–6)
Previous experiences
Mice having had chip experience during the acclimatization period occupied the chip bedding slightly more often than those who had experienced shaving bedding. This was more obvious in BALB/c mice. The influence of animals' previous bedding experience (acclimatization period) was significant, but did not influence the main results: all mice significantly preferred the shaving bedding in all test combinations.
Crossings between the cages
Crossing frequency was higher in combination C (669.7 ± 797.6 per animal and day) than in A (478.7 ± 332.1) and B (462.2 ± 213.4), especially for the C57BL/6J mice. C57BL/6J mice showed a continuously higher crossing frequency in combination C during the entire period, while the crossing frequency of BALB/c decreased after two days. BALB/c mice were less active than C57BL/6J mice for all combinations (A: P = 0.0151; B: P < 0.0001; C: P = 0.0080). Both strains had a higher crossing activity during the dark period (paired t-test P < 0.001).
Food and water consumption
C57BL/6J mice drank slightly more, but ate significantly less than BALB/c mice (ANOVA, P = 0.0021). Overall mice consumed more food on shavings than on chips (A: 60.32 ± 16.5%, B: 58.63 ± 21.9%, C: 62.57 ± 16.6%) (Figure 7), a significant difference (paired t-test P = 0.0312) was only found in combination C, mainly due to the C57BL/6J groups (paired t-test P = 0.0392). For water intake there was no significant difference in any of the combinations (Figure 7).
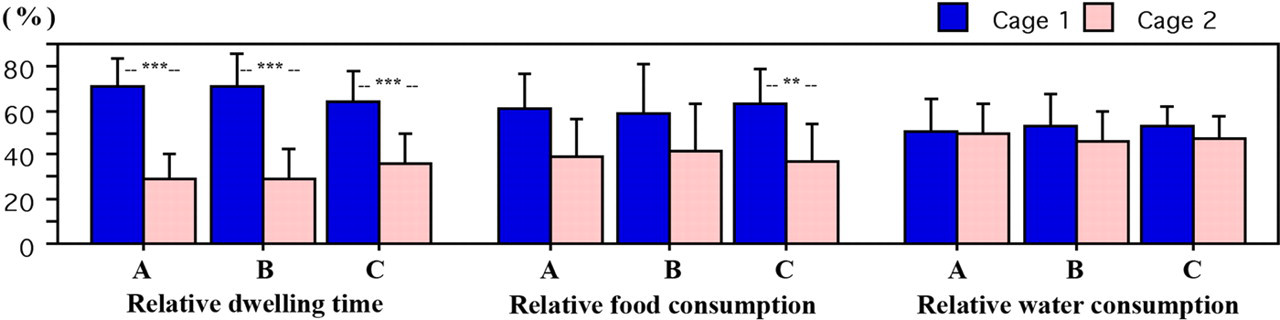
Relative dwelling time and food/water consumption during the preference test
While C57BL/6J mice consumed more water in the cages with shavings and nearly reached a statistical difference in combination A (paired t-test P = 0.0640), the BALB/c mice did not show a clear preference. Nevertheless, neither food nor water consumption significantly correlates with the relative dwelling time.
Discussion
In the present study, mice manifestly discriminated between different structures of softwood bedding, highlighting the importance of the structural aspect of bedding. Independent of the chips' particle size, female mice of both strains (C57BL6/JOlaHsd and BALB/cOlaHsd) showed a very clear preference for bedding with a shaving structure. This preference was evident both when data from individual mice were analysed and when the analysis was based on the average data of each cage. This tallies with previous studies, where individually tested mice and rats always preferred bedding consisting of long and coarse particles, which can be formed and manipulated easily. 32,35–38 The present results show that group-housed mice have the same structural preference. The relative ranking of the different structures (shavings≫coarse-grained chips>medium chips = fine chips) is congruent with these findings as well: Mice chose bedding structures suitable for manipulation and nest building.
All tested bedding products were commonly used wood beddings. Long paper stripes, cotton tissue or wood wool consisted of larger particles and were therefore preferred in other studies. 32,35,37 Nevertheless, these structures are also appropriate as nesting materials and mostly used for this purpose. Ideally, the preferred wood shavings should be combined with additional nesting material. As the purpose of the present study was to determine pure bedding preferences, no additional nesting material was provided. Mice had to construct their nests out of the bedding. This might explain why the coarse-grained chips were occupied more than the two finer ones. In this context, the detected strain differences are noticeable: although BALB/c mice are known to build better nests, 47 their bedding preferences are not as pronounced as those of the C57BL/6J mice. The nest building suitability of the tested bedding structures will be part of further studies.
Owing to the fact that the preferences were very constant during the test week, it can be presumed that they would also last over longer periods. Even in mice with chip experience, the shaving preference was stronger than a potential learned bias. 27,48 This indicates that previous experiences during the acclimatization period exert influence on the strength of preference but not on which option is preferred.
It has been reported that C57BL/6J mice showed more activity (1/3 more) during the light period than BALB/c mice 49 and performed around 20% of daily activity in the light period. 50 Expectedly, the crossing activity of C57BL/6J mice was higher than that of BALB/c mice. 51 Mice in combination A and B spent significantly more time on the shavings than mice in combination C, but showed less crossing activity in combinations A and B. This could be explained by the fact that the preference for the shavings was increased, when the fine- and middle-grained chips were offered to compete with the shavings. Although both strains in combination C moved more frequently at the beginning of the preference test, only C57BL/6J mice showed a continuously higher crossing frequency during the entire period. These results may indicate that chips with a larger size are more attractive for C57BL/6J mice than smaller chips, but not for BALB/c mice, therefore the crossing frequency of BALB/c mice decreased after a few days. In our primary experiment (validation, unpublished data), mice travelled on average 340 times (BALB/c) to 420 times (C57BL/6J) per day between two identical cages. As mice were highly motivated to use and monitor additional space with a high frequency, 52 the measured crossing activity could be assumed to be normal and may also account for mice moving between two cages during the bedding preference test. Food and water consumption were measured, due to which food and water intake may correlate with dwelling time. 41 As food and water were consumed mostly during the active phase (around 80% 53,54 ), it was predicted that the distribution of food and water consumptions would be more similar to the dwelling time in the dark phase. This presumption is not entirely correct. Similar to Ago et al., 35 both strains showed higher food consumptions in the preferred cage, but only C57BL/6J mice consumed more water on the shavings (no difference for BALB/c mice). Nevertheless, no statistical correlation between dwelling time and food/water consumption could be determined for any of the strains. This is in line with previous studies 36,32,38 suggesting that the distribution of food and water consumptions is not necessarily correlating to housing preferences and that the results can vary due to the strain used and choices provided in a study.
Blom et al. 36 showed that while most behaviours are equally distributed over cages with different bedding, resting, grooming and digging are primarily performed in association with the preferred bedding material. In our study, the importance of adequate contact bedding for the resting behaviour of mice is emphasized by the more pronounced bedding preferences during the light phase, i.e. resting period. This finding is also in accordance with various other studies. 30–32,36,38,48
It is undisputed that an ideal bedding does not exist. 23 Consequently, an animal facility has to choose between different bedding products and this choice always means a compromise between technical or hygienic aspects on the one hand and animal welfare aspects on the other hand. Compared with wood shavings, wood chips have diverse technical and hygienic advantages 19 and therefore are increasingly used. Nevertheless, our results show that a shaving structure is more preferred by C57BL/6J and BALB/c female mice in comparison with softwood chips, thus the shavings are more recommendable. If it is necessary to provide softwood chips, for example, due to technical reasons, coarse-grained with large particles would be proposed more than the low-formability fine chips. Information about material-related bedding preferences, long-term bedding preferences, preference strength (consumer demand studies), bedding-dependent health parameters and other aspects of welfare is still needed to further improve housing standards for laboratory mice.
Footnotes
ACKNOWLEDGEMENTS
We wish to express our thanks to the ECLAM–ESLAV Foundation and the Hans and Helga Maus Stiftung for supporting this research.