
Abstract
Oestrogen administration can alter experimental stroke outcomes. Soy as a source of phytoestrogens may therefore modulate responses in ‘oestrogen-sensitive’ stroke models, thus potentially confounding results. We evaluated the effects of dietary soy on acute infarct volumes in a pilot study using a rat focal stroke model. We hypothesized that ovariectomized (OVX) rats fed a soy-rich diet would have smaller acute infarct volumes than rats fed a soy-free diet. OVX rats were randomly assigned to a soy-free (n = 6) or a soy-rich (n = 6) diet for four weeks and weighed weekly. Following the dietary trial, rats underwent 2 h of middle cerebral artery occlusion (MCAO). Mean arterial blood pressure, rectal and temporalis muscle temperatures, arterial blood gases and blood glucose were recorded peri-ischaemia. Rats were euthanized 22 h following 2 h of MCAO. Brains were stained with 2,3,5-triphenyl tetrazolium chloride for acute infarct volume analysis. Uterine weight and histology were also evaluated as additional internal oestrogen-sensitive controls. Rats on the soy-free diet had greater gains in body weight (259 ± 6% baseline body weight) than rats on the soy-rich diet (238 ± 4% baseline body weight). No differences were seen in uterine weight and histology, peri-ischaemic physiological parameters and infarct volumes between the treatment groups. The results of this pilot study suggest that the dietary soy level tested may not alter acute infarct volumes in ischaemic female rat brain. More studies addressing the potential confounding effects of dietary soy in ‘oestrogen-sensitive’ stroke models are needed if investigators are to make informed choices regarding diets used in experimental stroke research.
Exogenous oestrogen administration has been shown to alter experimental outcomes in rodent stroke models. 1 Structurally and functionally similar to oestrogen, phytoestrogens exhibit oestrogenic and antioestrogenic effects and are found in high abundance in most soy-containing foods.2,3 The main classes of phytoestrogens are isoflavonoids, which include isoflavones and coumestans, and lignans. Commercial laboratory rodent diets commonly use soymeal as their protein source, which contains predominantly the isoflavones genistein and daidzein in levels ranging from 100 to 600 μg/g.4-8 Because several widely used rodent stroke models are oestrogen-sensitive, 1 an important laboratory animal husbandry factor like dietary soy as a major phytoestrogen source could modulate physiological and behavioural responses similar to oestrogen in these models, thus potentially confounding experimental outcomes.
Several studies have demonstrated effects of phytoestrogens on uninjured rodent brain. For example, neurobehavioural studies have shown a direct link between dietary phytoestrogen consumption, high plasma isoflavone levels and alterations in learning and memory. 9 Morphological, biochemical and physiological brain changes have been demonstrated as well. Dietary phytoestrogen manipulations can alter the weight of the sexually dimorphic nucleus of the preoptic area of the brain, 10 cause a decrease in brain calcium-binding proteins 11 and produce anxiolytic effects. 12 These diet-induced changes in the brain could therefore have profound implications in rodent stroke models.
Very few studies have addressed the potential confounding effects of soy-derived dietary phytoestrogens in a rodent stroke model whose outcomes are influenced by oestrogen. Based on the paucity of information about the effects of dietary phytoestrogens in normal and injured rodent brain, researchers can only speculate about the possible consequences of dietary soy from laboratory rodent diets in oestrogen-sensitive rodent stroke models. Extrapolation from existing rodent studies outside of neuroscience is difficult due to a number of complexities, including differences concerning in vitro and in vivo oestrogenic potency, interactions with other binding proteins, dose–response variability, processing techniques and environmental influences.8,13,14 Studies evaluating the effects of dietary soy as a source of phytoestrogens in rodent stroke models are clearly needed if primary investigators are to make informed choices regarding rodent diets used in ischaemic brain research.
In this pilot study, we have chosen to evaluate the potential confounding effects of dietary soy in an established model of acute transient focal cerebral ischaemia in the ovariectomized (OVX) female rat. Infarct volume, a commonly used outcome in this acute model, has previously been shown to be reduced by exogenous oestrogen administration in male and OVX female rodents.1,15,16 We hypothesized that dietary soy will alter infarct volume acutely in OVX rats and thus could be a potential confounder when evaluating acute infarct volumes in our experimental stroke model. We therefore determined whether dietary soy reduces acute infarct volumes by evaluating the effects of soy-free versus soy-rich rodent diets in OVX rats. Body weight, uterine weight and histology are also influenced by oestrogen in female rodents and served as additional oestrogen-sensitive controls independent of acute infarct volume outcomes in the brain.
Materials and Methods
Animals and husbandry
Twenty-three age-matched (4–7 weeks old) female Wistar rats (Hsd:WI, Harlan Sprague Dawley, Madison, WI, USA) were used. According to vendor health monitoring reports, rats were negative for the following viral pathogens: Hantaan virus, Kilham's rat virus, lymphocytic choriomeningitis virus, mouse adenovirus types 1 and 2, pneumonia virus of mice, rat minute virus, rat parvovirus, rat Theiler virus, respiratory enteric virus III, Sendai virus, sialodacryoadenitis virus and Toolan's H-1 parvovirus. Animals were also negative for the following bacterial, mycoplasmal and fungal pathogens: CAR bacillus, Citrobacter rodentium, Clostridium filiforme, Corynebacterium kutscheri, dermatophytes, Helicobacter bilis, Helicobacter hepaticus, Helicobacter spp., Mycoplasma pulmonis, Pasteurella pneumotropica, Pneumocystis carinii, Pseudomonas aeruginosa, Salmonella spp., Staphylococcus aureus, Streptobacillus moniliformis, Streptococcus pneumonia and Streptococcus zooepidemicus. Additionally, animals were free of ectoparasites, endoparasites, enteric protozoan and Encephalitozoon cuniculi.
All aspects of the study were approved by the Oregon Health and Science University Institutional Animal Care and Use Committee and were performed in accordance with the institutional policy and the National Institutes of Health guidelines governing the humane treatment of vertebrate animals. Animals were maintained in accordance with the Guide for the Care and Use of Laboratory Animals in a facility accredited by the Association for the Assessment and Accreditation of Laboratory Animal Care, International. Rats were pair-housed in static polycarbonate isolator cages (area, 840 cm2; height, 20 cm) on ALPHA-dri™ bedding (Shepherd Specialty Papers, Chicago, IL, USA) in a temperature-controlled (20.0–22.2°C) and humidity-controlled (30–70%) room on a 12:12 h light:dark cycle. All animals were allowed to acclimatize to the vivarium for 11 ± 1 days before undergoing ovariectomy (Figure 1). Rats had ad libitum access to assigned experimental diets and ultraviolet filtered water. Animals were weighed upon arrival and weekly thereafter.
Experimental diets
Diets used in this study were TD.05087 and TD.96155 (Harlan TekLad, Madison, WI, USA). The TD.05087 diet includes 25.8% soybean meal, for a final total daidzein and genistein level of 500 ppm or mg/kg. This value, as well as value ranges cited from other references and laboratories in this paper, refers to the sum of genistein and daidzein expressed as aglycone or ‘free isoflavone.’ A review of the literature demonstrates that commercial soymeal diets have daidzein and genistein levels ranging from 100 to 600 ppm.4-8 Although post-production analysis was not performed, the soybean meal used in this lot was tested at 1940 ppm genistein and daidzein before production. The TD.96155 diet which is a minimal phytoestrogen diet (no soybean meal) was chosen as the soy-free diet. Although this diet was not analysed post-production for daidzein and genistein, the typical background level is less than 20 ppm. The same lot and batch number of each diet was used throughout the study. The formulation and nutrient analysis of the two experimental diets are summarized in Table 1.
Composition of the experimental diets
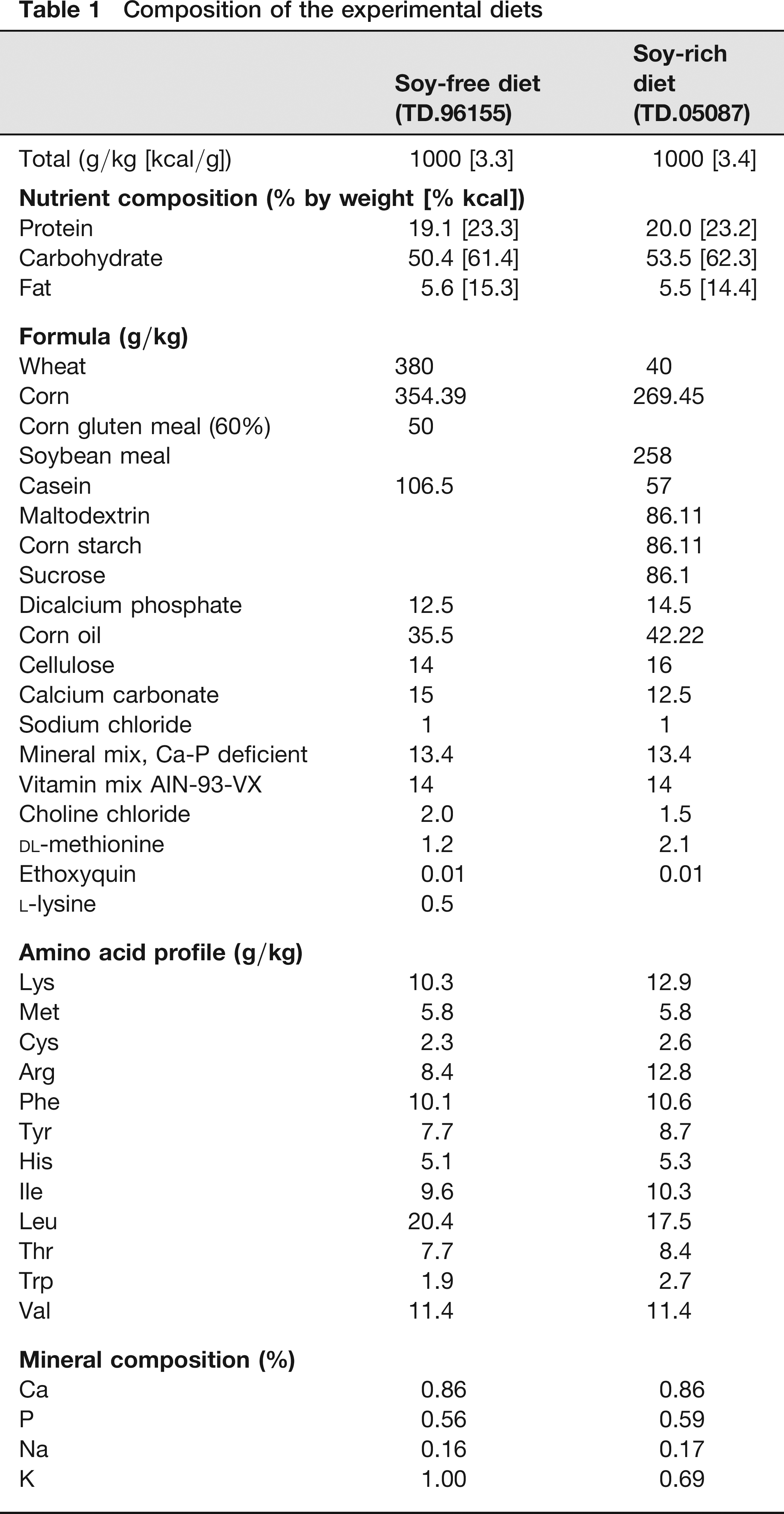
Rats were randomized to receive either soy-free (TD.96155, n = 11) or soy-rich (TD.05087, n = 12) rodent diets. Animals were placed on their assigned diet upon arrival at the study facility and remained on these diets throughout the study. Figure 1 shows the experimental time line relative to initiation of experimental diets, ovariectomy, experimental stroke (middle cerebral artery occlusion [MCAO]) and euthanasia.
Schematic overview of the experimental time line relative to initiation of experimental diets, ovariectomy, experimental stroke (middle cerebral artery occlusion [MCAO]) and euthanasia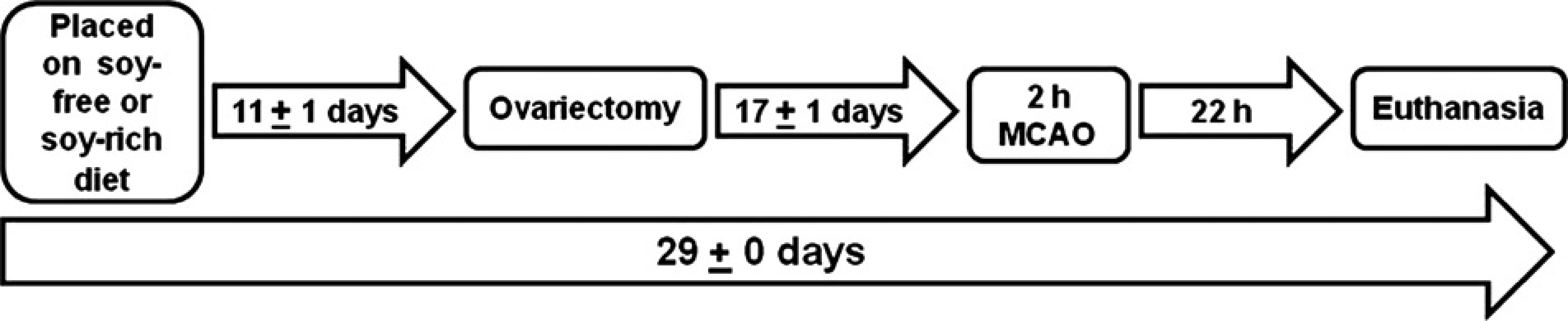
Ovariectomy
OVX rats were used to ensure relative uniformity of the animals’ endogenous hormonal background and to minimize interference from endogenous oestrogen during most of the dietary trial and experimental stroke studies. OVX rats were used instead of males as our laboratories have ongoing studies evaluating the effects of oestrogen and hormone replacement therapy in females on stroke outcomes. Ovariectomy was performed on young female rats (5–8 weeks) 17 ± 1 days before experimental stroke (Figure 1). Surgery was performed aseptically by a single surgeon blinded to dietary treatment group. Surgical anaesthesia was induced via chamber induction with 4–5% halothane and maintained with 1–2% halothane via face-mask in oxygen-enriched air. A 1 cm paracostal incision approximately 0.5 cm caudal to the last rib was made through the skin and then through the lateral body wall on one side of the animal after the intended surgical site was clipped free of hair and the skin prepared with beta-dine. The ovarian artery and vein were ligated and the ovary removed. The lateral body wall incision was sutured, and the skin closed with skin staples. The same surgical approach was then repeated on the opposite side. Rats were recovered and returned to animal quarters. Skin staples were removed 7–10 days following ovariectomy.
Analgesics were not given after ovariectomy as the use of non-steroidal anti-inflammatory drugs and opioids preceding acute experimental surgical stroke models can potentially confound outcomes. 17 However, all postoperative animals were scored based on previously described pain/distress indices18,19 two to three times daily postoperatively until skin staples were removed. Pain scores that fell within established ranges for moderate to severe pain mandated immediate veterinary consultation or euthanasia. 18
Reversible focal cerebral ischaemia
Surgery was performed aseptically by a single surgeon blinded to the dietary treatment group. Rats were anaesthetized with halothane (induction 4–5%, maintenance 1.25–1.5%) delivered via mask in oxygen-enriched air. Temporary femoral arterial cannulation was done for continuous monitoring of mean arterial blood pressure (MABP) and measurement of peri-ischaemic arterial blood gases and plasma glucose. Rectal and temporalis muscle temperatures were continuously monitored using temperature thermistors (Mon-a-therm, model 6510; Mallinckrodt Medical, Inc, St Louis, MO, USA) and maintained with warming blankets and heat lamps. In each rat, parietal cortical perfusion within the middle cerebral artery (MCA) territory was monitored by laser Doppler flowmetry (LDF) signal (model DRT4, Moor Instruments Ltd, Oxford, UK) obtained by probe placement 6 mm lateral and 2 mm posterior to the bregma.15,16 The effectiveness of vascular occlusion was determined by sustained reduction in LDF signal while restoration of blood flow following the release of vascular occlusion was assessed by increases in LDF signal towards baseline levels.
Reversible focal cerebral ischaemia was induced by MCAO via the intraluminal suture technique. 20 The right common and external carotid arteries were exposed and ligated. The right occipital artery was cauterized, and then the pterygopalatine artery was ligated. An incision was made in the common carotid artery distal to ligation and an occluding filament (4–0 monofilament nylon surgical suture with heat-rounded tip) advanced into the internal carotid artery to the MCA origin, a point at which LDF signal abruptly decreases, indicating low cortical flow.15,16 LDF signal was measured during ischaemia over 15–30 min intervals and for the first 15 min following termination of MCAO to confirm cortical reperfusion when the occluding filament was removed. To ensure relative uniformity of the ischaemic insult, animals were excluded if the mean ischaemic LDF signal was greater than 50% of the baseline LDF signal. Catheters and probes were removed after the final LDF measurement was made 15 min after termination of MCAO. Incisions were then closed, and the animals recovered from anaesthesia.
Pain-relieving drugs were not given after MCAO surgery as the use of non-steroidal anti-inflammatory drugs and opioids in acute experimental surgical stroke models can potentially confound outcomes. 17 However, all postoperative animals were evaluated for pain and distress according to a previously published behavioural scoring system based on subjective and objective measures for pain evaluation in the rat MCAO surgical model. 18 Pain scores that fell within established ranges for moderate to severe pain mandated immediate veterinary consultation or euthanasia. 18
After 22 h of recovery from MCAO, animals were deeply anaesthetized with halothane (4–5%), and blood samples for serum hormone (17β-oestradiol, progesterone) measurements were obtained by cardiac puncture before euthanasia via decapitation. Following euthanasia, brains and uteri were harvested for acute infarct volume and weight determinations subsequent to fixation, respectively.
Infarct volume determinations
Brains were sliced into 2 mm thick coronal sections (seven slices total) using a brain matrix (Kent Scientific Corporation, Torrington, CA, USA). Coronal sections were placed in a 1% 2,3,5-triphenyl tetrazolium chloride (TTC; Sigma, St Louis, MO, USA) and incubated at 37°C for 30 min. 21 Stained sections were then fixed in 10% neutral-buffered formalin. Viable brain areas were stained dark red due to reduction of TTC by mitochondrial enzymes, whereas infarcted areas were unstained and appeared white. The anterior and posterior sides of each stained coronal section were digitally photographed. Regional areas (cortex, striatum, hemisphere) within each cerebral hemisphere and infarct areas within each region were measured on the anterior and posterior sides of each coronal section using digital image analysis (SigmaScan Pro; SPSS Science, Chicago, IL, USA). Regional and infarct volumes for each individual coronal slice were then determined by averaging the anterior and posterior areas and then multiplying the average area by the thickness of the coronal section. Regional and infarct volumes were then integrated across all coronal sections and expressed as mm3, or indirectly as a percent of the contralateral structure (cortex, striatum and hemisphere) as a partial correction for oedema. 22
Uterine weight and histology
In each rat, the uterus was cut just above the junction of the uterine body with the cervix and at the junction of the uterine horns with the ovaries. Each uterus was then removed, trimmed free of fat and mesentery, blotted to remove excess fluid and weighed. Uterine-to-body weight ratios were calculated and presented as % terminal body weight (uterine weight/body weight x 100). Afterwards, 10% neutral-buffered formalin was injected into the uterine lumen until the uterus was visibly extended. Uteri were then placed in 10% neutral-buffered formalin for further fixation and eventual histological examination. Tissue was dehydrated, embedded in paraffin, sectioned at 4–5 μm, and stained with haematoxylin and eosin. Uterine histopathological analyses were performed by a single individual blinded to dietary treatment group.
Serum hormone measurements
Serum 17β-oestradiol and progesterone were measured in duplicate with commercial radioimmunoassay kits (Diagnostic Products Corp, Los Angeles, CA, USA). 17β-oestradiol was measured as it is the primary oestrogen observed in rodents and in women. Lower limit of detection for the 17β-oestradiol radioimmunoassay is 10 pg/mL. Samples with 17β-oestradiol levels <10 pg/mL were not used in calculating mean oestradiol levels per treatment group.
Statistical analysis
Values expressed as mean ± SEM. Using data from previous stroke studies with oestrogen, we performed a pretest power analysis (desired power = 0.8, number of groups = 2, α = 0.05, minimum detectable difference in means = 15, expected standard deviation of residuals = 8) to determine the minimum number of OVX rats needed to observe preliminary trends and differences in infarct volume. Based on these analyses, six OVX rats per experimental group were recommended for the proposed pilot study to examine the potential confounding effects of dietary soy on acute infarct volume outcomes. Baseline body weight was subjected to one-way analysis of variance (ANOVA) with post hoc Newman-Keuls test. Weekly body weight (g, % baseline body weight) was subjected to two-way ANOVA with post hoc Newman-Keuls test. Differences in mean ischaemic LDF (% baseline), infarction volume (mm3, % contralateral structure), serum hormone levels (17β-oestradiol, progesterone) and uterine weight (mg, % terminal body weight) between dietary groups were determined by one-way ANOVA with post hoc Newman-Keuls. Physiological variables (blood gases, blood glucose, MABP, rectal and temporalis muscle temperatures) and LDF signal were subjected to two-way ANOVA with post hoc Newman-Keuls test. Statistical significance was P < 0.05. Statistical analyses were performed using SigmaStat Statistical Software, Version 3.1 (SPSS, Inc, Chicago, IL, USA).
Results
Exclusions relative to experimental focal cerebral ischaemia
In the soy-free diet group (n = 11), four animals were excluded due to intraoperative death (n = 2, 18%), death during the post-MCAO recovery period (n = 1, 9%) or tearing of the right common carotid artery during surgical manipulations (n = 1, 9%). In the soy-rich diet group (n = 12), two animals were excluded due to death during the post-MCAO recovery period. Therefore, surgical mortality rates were 36% (n = 4) in the soy-free diet group, 17% (n = 2) in the soy-rich diet group and 26% (n = 6) overall. Our mortality rates fall within the range of values reported by our laboratories and collaborators using a comparable 2 h rat model of transient MCAO with 22 h of recovery from MCAO.23-27
One animal (9%) met the mean ischaemic LDF exclusion criteria of greater than 50% of baseline LDF in the soy-free diet group. In the soy-rich diet group, four animals (33%) were euthanized due to failure to achieve MCAO based on LDF monitoring (n = 2, 17%), maintain MCAO based on LDF monitoring (n = 1, 8%) or met the mean ischaemic LDF exclusion criteria of greater than 50% of baseline LDF (n = 1, 8%). Soy-free (n = 5, 45%) and soy-rich (n = 6, 50%) diet groups therefore had similar total numbers and percentages of rats that died or were excluded based on LDF monitoring and exclusion criteria. Consequently, only six rats per dietary group were included in the final data analysis.
Dietary effects on serum hormone levels
There were three samples from the soy-free diet group versus no samples from the soy-rich diet group that were below <10 pg/mL for serum 17β-oestradiol levels. Although not statistically significant (P = 0.093), serum 17β-oestradiol levels were generally lower in the soy-free diet group (17 ± 2 pg/mL, n = 3) compared with the soy-rich diet group (23 ± 2 pg/mL, n = 4) (Table 2). There were no significant differences (P = 0.719) in serum progesterone levels between the soy-free (12 ± 1 ng/mL, n = 6) and soy-rich (11 ± 3 ng/mL, n = 4) diet groups (Table 2).
Plasma 17β-oestradiol and progesterone levels at 22 h following 2 h of middle cerebral artery occlusion in ovariectomized Wistar rats fed soy-free (n = 6) or soy-rich (n = 6) diets for four weeks before experimental transient focal cerebral ischaemia
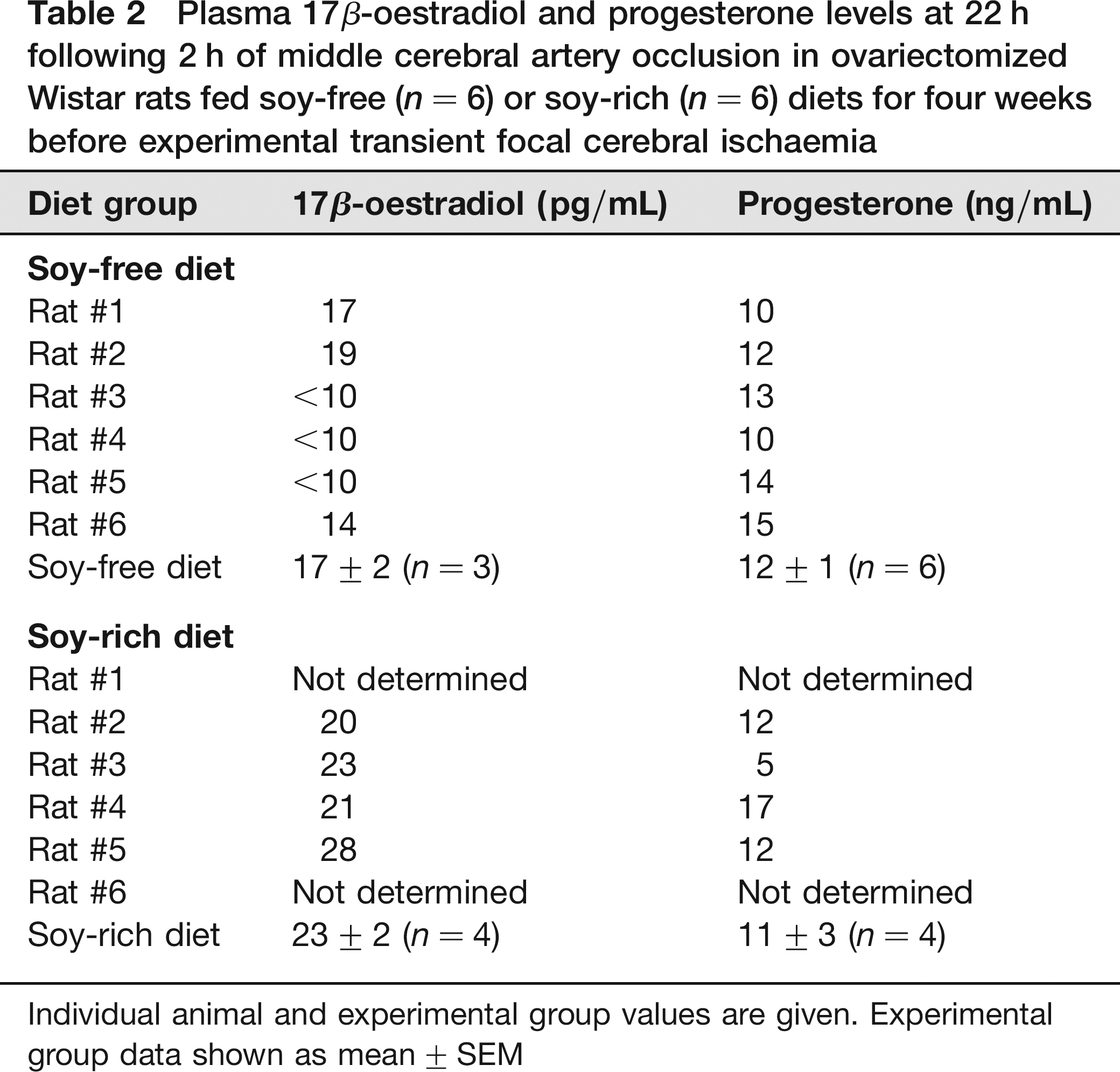
Individual animal and experimental group values are given. Experimental group data shown as mean ± SEM
Dietary effects on body weight and uterus
There were no significant differences (P = 0.731) in baseline body weights for soy-free (108 ± 2 g, n = 6) and soy-rich (109 ± 2 g, n = 6) diet groups (Figure 2a). All animals had significant gains in weekly body weight throughout the course of the study regardless of the assigned dietary group (Figure 2). The soy-free diet group (279 ± 7g) was observed to be significantly heavier than the soy-rich diet group (258 ± 7 g) only during the fourth and final week of the study (Figure 2a). However, significant differences in weekly body weight as a percentage of baseline values were observed during the third and fourth weeks of the study (Figure 2b), with the soy-free diet group (week 3, 201 ± 5%; week 4, 259 ± 6%) having greater gains in body weight than the soy-rich diet group (week 3, 191 ± 3%; week 4, 238 ± 4%).
Weekly body weights expressed as (a) absolute weight in g and as (b) % baseline body weight in ovariectomized female Wistar rats fed either a soy-free or a soy-rich diet before experimental transient focal cerebral ischaemia. Values are mean ± SEM. *P < 0.05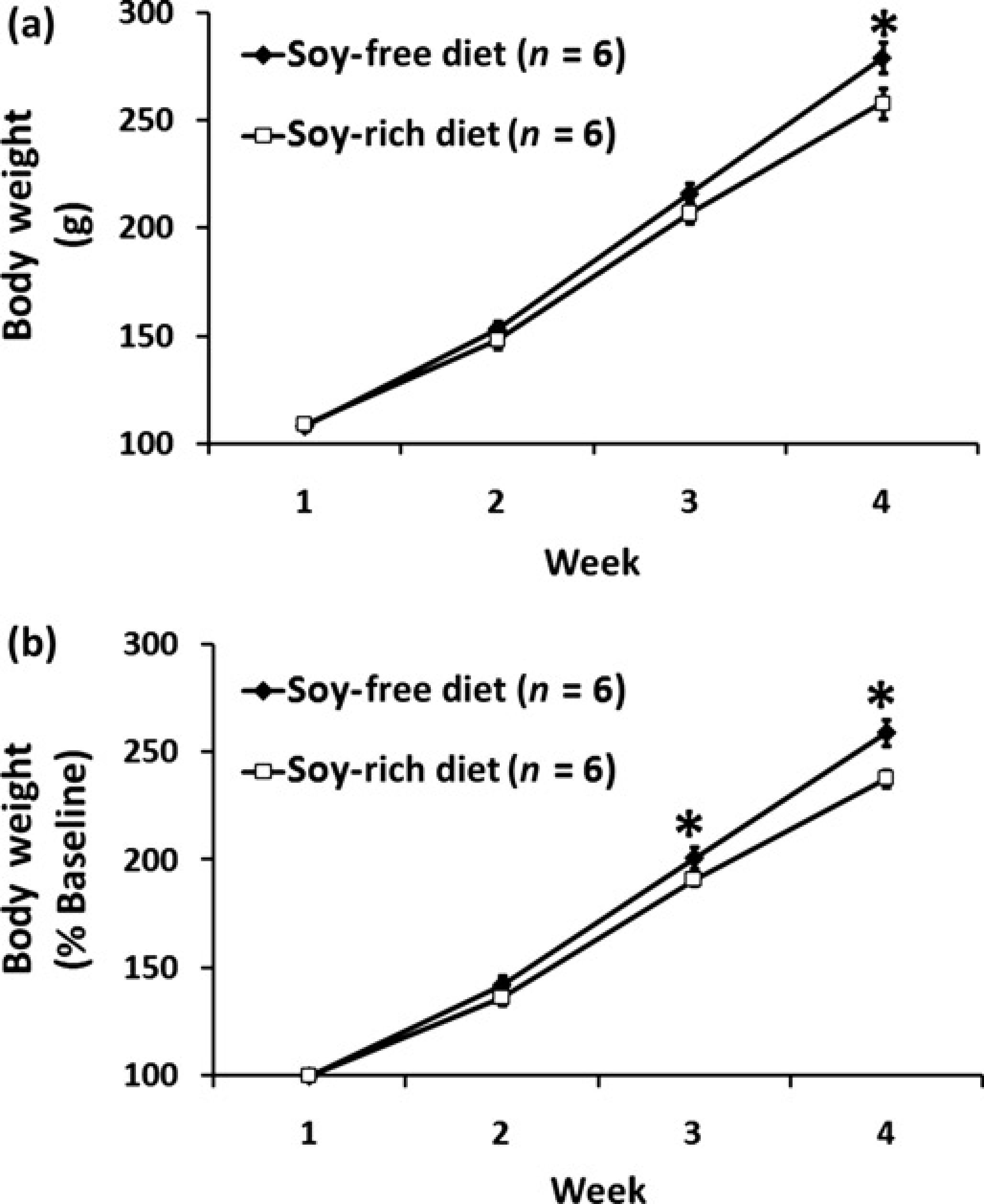
There were no significant differences (P = 0.440, P = 0.867) in uterine weight (mg, percentage of terminal body weight) for soy-free (168 ± 5 mg, 0.061 ± 0.003%; n = 6) and soy-rich (159 ± 10 mg, 0.062 ± 0.003%; n = 6) diet groups. Regardless of the dietary treatment group, histological evaluation of multiple sections from each animal demonstrated similar findings of a narrowed uterine lumen and limited endometrial gland development (data not shown).
Dietary effects in ischaemic brain
Baseline, intra-ischaemic and early post-MCAO MABP, blood gases and glucose were comparable between diet treatment groups (Table 3). Peri-ischaemic rectal and temporalis muscle temperatures were not significantly different between diet groups (Table 3). Mean ischaemic LDF (% baseline signal) in OVX rats fed the soy-free (36 ± 3%) or soy-rich (45 ± 2%) diets was significantly different (P = 0.032). However, there were no differences in reduction and maintenance of residual intra-ischaemic LDF signal as well as in increase of residual post-MCAO LDF signal (expressed as a percent of baseline signal) between diet groups (Figure 3). No significant differences in regional and total infarct volumes (lesion size - mm3, % contralateral structure) were observed between the soy-free (cortex, 65 ± 22 mm3, 23 ± 7%; striatum, 31 ± 6 mm3, 70 ± 11%; hemisphere, 96 ± 22 mm3, 13 ± 3%) and soy-rich diet groups (cortex, 45 ± 23 mm3, 15 ± 7%; striatum, 25 ± 13 mm3, 40 ± 18%; hemisphere, 70 ± 35 mm3, H ± 6%,) (Figure 4).
Cerebral cortical microvascular perfusion as determined by laser Doppler flow (LDF) signal during 2 h of middle cerebral artery occlusion (MCAO) and the first 15 min following recovery from MCAO. LDF was measured over the ipsilateral parietal cortex and expressed as a percentage of baseline signal in ovariectomized female Wistar rats fed either a soy-free or a soy-rich diet before MCAO. There were no differences in reduction and maintenance of residual intra-ischaemic LDF signal as well as in increase of residual post-ischaemic LDF signal between diet groups. Values are mean ± SEM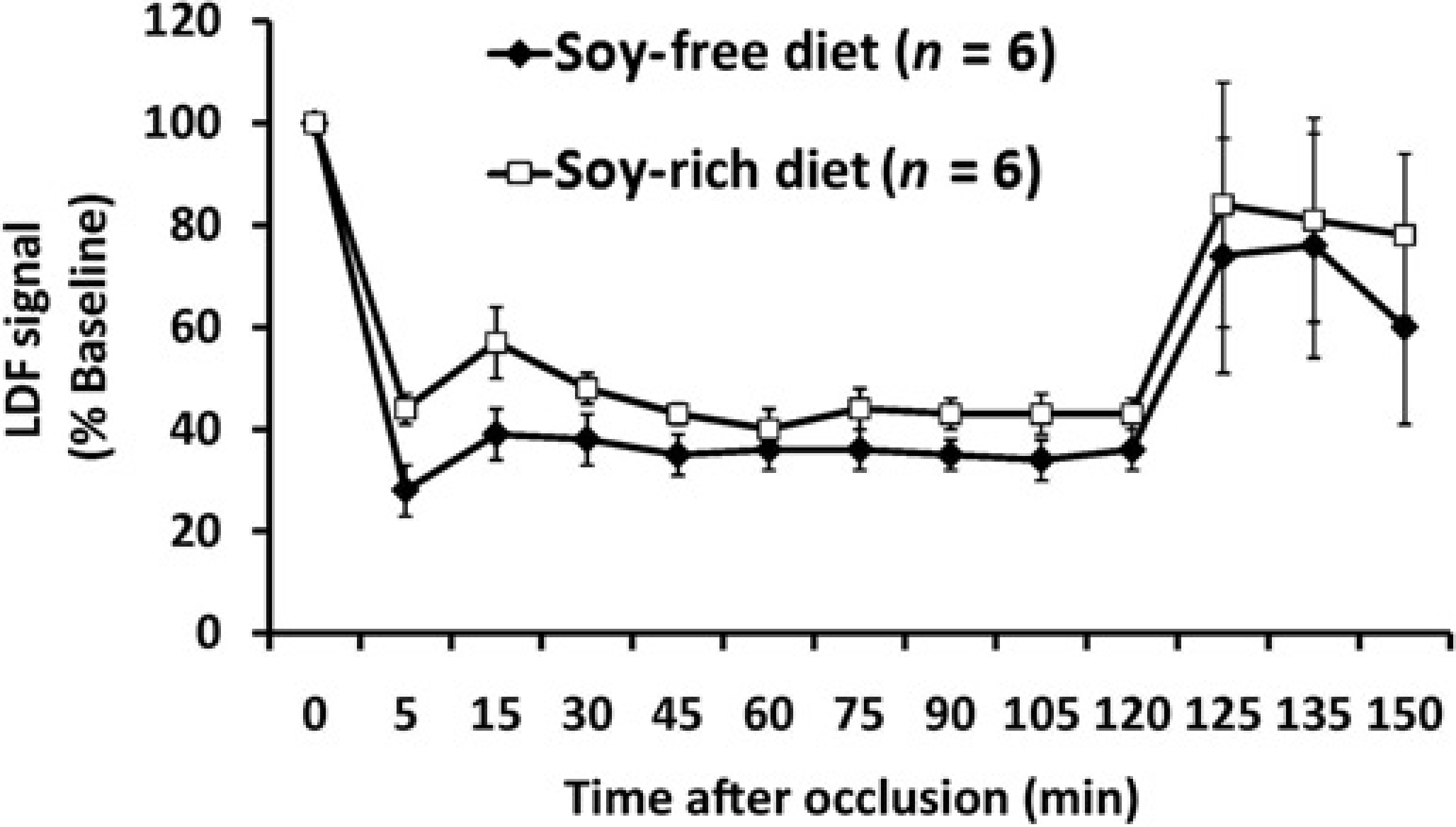
Cortical, striatal and total hemispheric infarction volumes as determined by 2,3,5-triphenyl tetrazolium chloride staining (expressed as (a) mm3 and indirectly as (b) % contralateral structure as a correction for oedema) in ovariectomized female Wistar rats fed either a soy-free or a soy-rich diet before 2 h of middle cerebral artery occlusion (MCAO) followed by 22 h of recovery from MCAO. There were no differences in regional and total infarction volumes between treatment groups. Values are mean ± SEM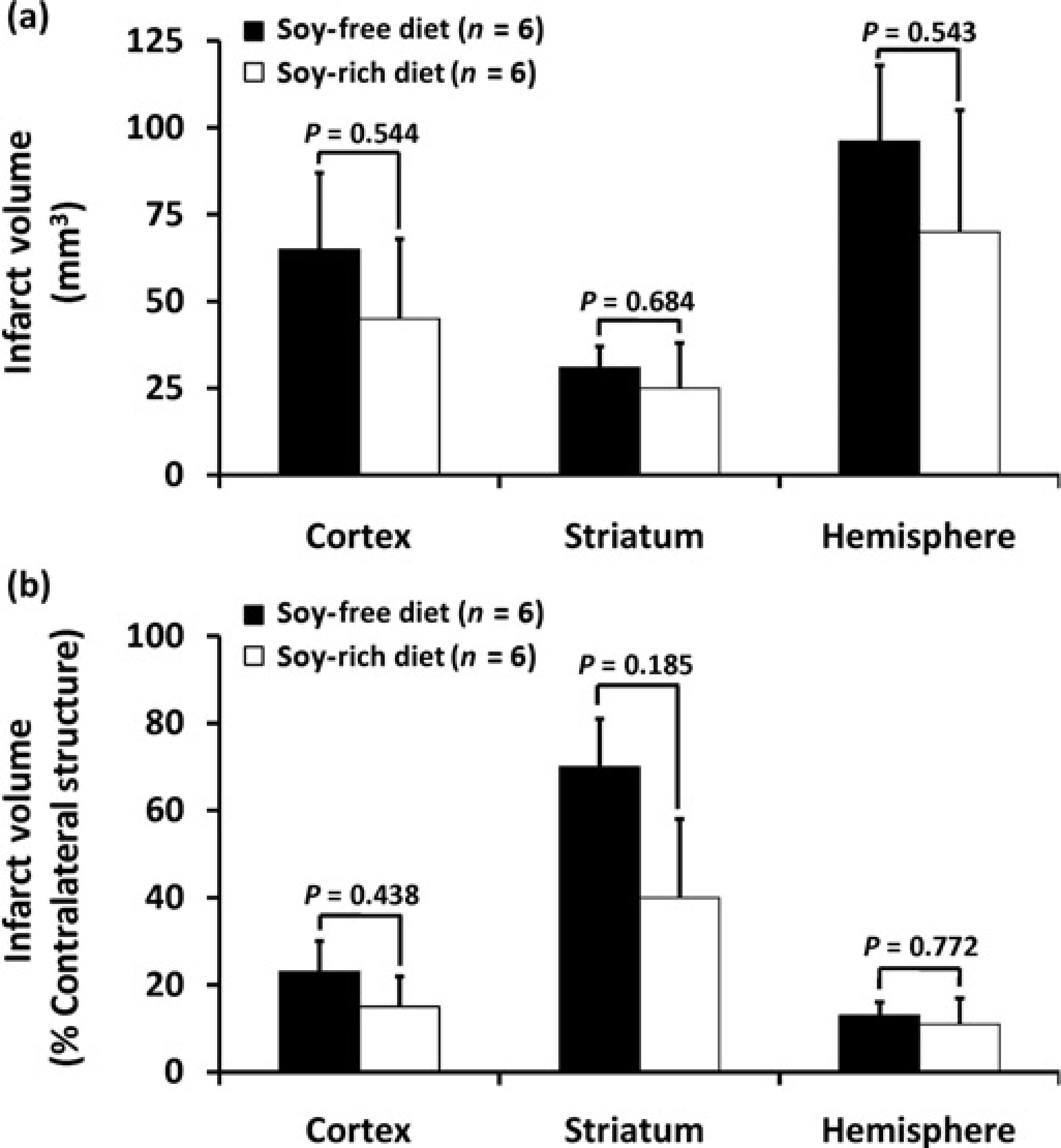
Physiological parameters before, during and after 2 h of middle cerebral artery occlusion (MCAO) in ovariectomized Wistar rats fed soy-free (n = 6) or soy-rich (n = 6) diets for four weeks before experimental transient focal cerebral ischaemia
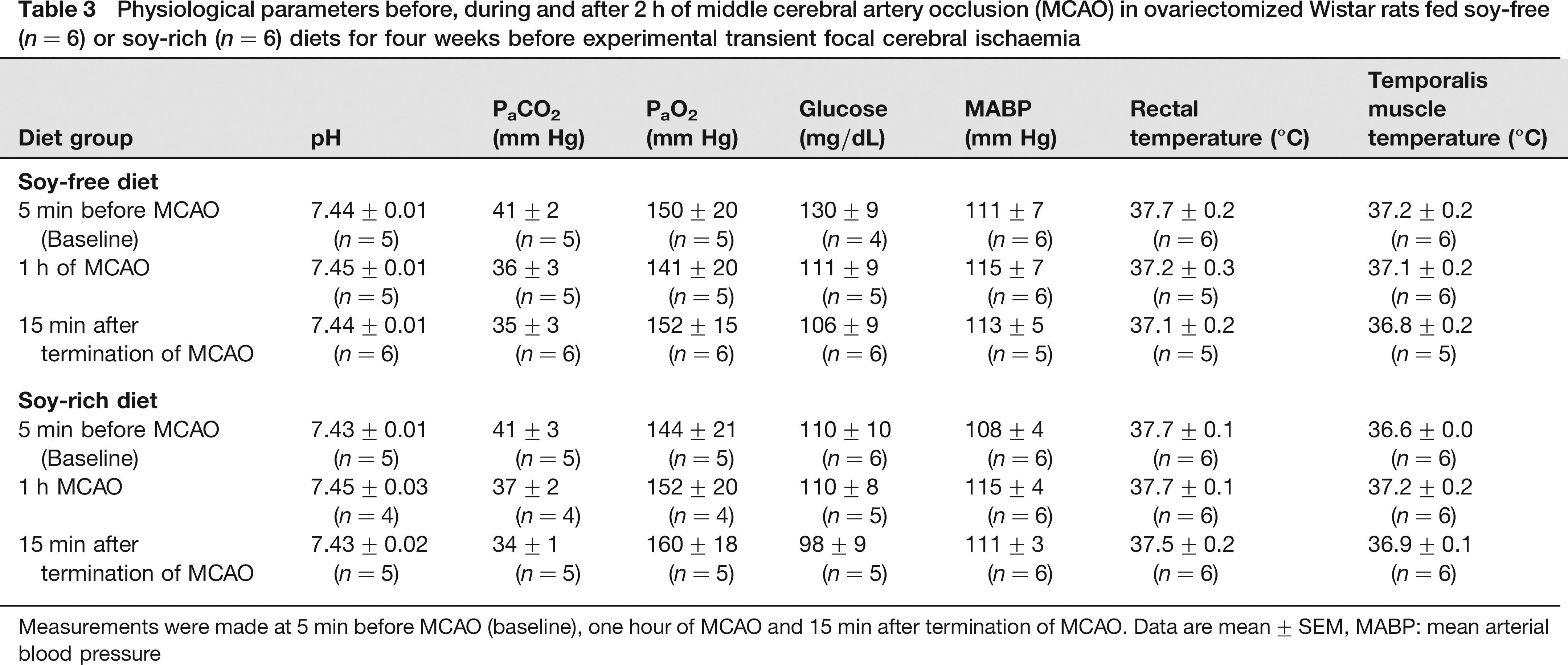
Measurements were made at 5 min before MCAO (baseline), one hour of MCAO and 15 min after termination of MCAO. Data are mean ± SEM, MABP: mean arterial blood pressure
Discussion
This pilot study demonstrates three findings. First, the presence of dietary soy at the level tested may have contributed to the attenuated weight gain in OVX rats. Second, there were no differences in uterine weights and histology in OVX rats between the soy-free and soy-rich diet groups. Third, acute infarct volumes 22 h following MCAO were similar regardless of the presence or absence of dietary soy. Our preliminary results suggest that dietary soy at the level tested may not alter acute infarct volumes in ischaemic female rat brain in our experimental rat focal stroke model.
Previous rodent stroke studies have focused on individual phytoestrogens28–33 rather than on dietary sources of phytoestrogens present in laboratory rodent chows. To our knowledge, this is the first study to evaluate the effects of dietary soy on acute outcomes in OVX rats following 2 h of transient focal cerebral ischaemia. We found that dietary soy at the levels tested did not appear to alter acute infarct volumes. Other rat studies with dietary phytoestrogens have evaluated acute outcomes in males and OVX females after 90 min of transient focal cerebral ischaemia34,35 or in OVX females following 24 h of permanent focal cerebral ischaemia. 36 In contrast to our findings, these studies using diets high in isoflavones demonstrated varying degrees of reduced infarct volumes in ischaemic brain.34–36 Age, gender, dietary trial length, total isoflavone content of the diets, other dietary components and duration of ischaemia could explain some of the differences in our findings relative to the effects of individual dietary phytoestrogens.
In this pilot study, serum 17β-oestradiol levels were generally lower in the soy-free diet group than in the soy-rich diet group while serum progesterone levels were similar. These results would suggest that ovariectomy reduced progesterone levels comparably but that the presence of dietary soy at the level tested may have increased oestradiol levels. In humans, dietary phytoestrogens have variable effects on oestradiol levels, with lower doses increasing and higher doses reducing oestradiol levels. 37 Based primarily on in vitro assays, phytoestrogens can potentially alter oestradiol biosynthesis and metabolism through modulation of steroidogenic enzyme activity and expression, thereby altering serum oestradiol levels. 38 However, phytoestrogen effects on oestrogen steroidogenesis are not as well characterized in vivo? 9 Another mechanism by which soy-based dietary phytoestrogens could promote increased oestradiol levels is through stimulation of gastrointestinal deconjugation of oestrone, thus leading to its peripheral reabsorption and conversion to oestradiol. 40 However, the small changes in oestradiol levels promoted by the soy-rich diet in our study were not sufficient to alter acute infarct volume outcomes.
Oestrogen is known to attenuate body weight gains in rats.36,41 In our study, OVX rats on the soy-rich diet had lower gains in body weight compared with animals on the soy-free diet. This is in agreement with other studies demonstrating that modest to high levels of dietary phytoestrogens can decrease body weight gains in rats.10,12,42,43 However, these studies did not assess circulating oestrogen levels as was done in this study. Therefore, the small differences in circulating oestrogen levels observed between dietary groups in this study may have been a contributing factor to the differences in body weight gain seen between the two dietary groups.
Differences in body weight gain could also be due to differences between the two diets in other nutritional factor levels of protein, carbohydrates fat, and metabolic energy. Although the protein, carbohydrate and fat content of both diets and the total kcal/g and g/kg were similar (Table 1), the formulation of the soy-free diet did require use of larger amounts of wheat, corn, corn gluten meal and casein to attain adequate protein levels which could have affected diet palatability and protein digestibility. Variable levels of trypsin inhibitor activity and biologically active dietary factors such as digestible amino acid levels, carbohydrate profile and non-starch polysaccharides between the diets might have contributed as well to diverging body weight gains between soy-free and soy-rich dietary groups.
Finally, although we did not measure food intake in our study, differences in food intake could potentially account for the differences in body weight gains observed between the two dietary groups. However, other studies have reported either no difference 34 or higher12,43 food intake in animals fed diets containing dietary soy or phytoestrogens compared with animals fed a soy- or phytoestrogen-free diet. Regardless of the possible reasons discussed above for the differences in body weight gains between the dietary groups, these differences in body weight changes between the two dietary groups did not appear to influence acute infarct volume outcomes.
Although oestrogen can increase uterine size and activity in the classical rat uterotrophic assay, 44 uterine responses to dietary phytoestrogens have been more variable depending on type, source, amount and food consumption rates.7,39,45 Previous studies suggest that modest to low levels of dietary phytoestrogens (350 μg total genistein equivalents/g diet or less) do not significantly increase baseline uterine weights or reduce uterine responsiveness. 7 Therefore, the current recommendation for conducting the rat uterotrophic bioassay is to use diets containing phytoestrogen levels less than 325–350 μg total genistein equivalents/g diet, as uterine responsiveness may be compromised at higher levels. 7 While the soy-rich diet tested in our study had higher phytoestrogen levels than the recommended limit for the rat uterotrophic bioassay, we did not observe any differences in uterine weight and activity as compared with the soy-free diet group. However, this recommendation is limited to immature females and does not include adult OVX females. 7 For control adult OVX animals, rat uterotrophic assay guidelines suggest that mean blotted uterine weights greater than 115 mg should be questioned. 7 In this study, mean blotted uterine weights in the soy-free diet group were greater than 115 mg. This suggests that control values may be anomalous for reasons unrelated to dietary phytoestrogens. Dietary factors other than phytoestrogen levels have been shown to affect uterine weights7,46-48 and may have been a contributing factor in the soy-free diet group.
Studies on cerebral vessels and cerebral blood flow (CBF) would suggest that phytoestrogens may act as relaxants and vasodilators.49-51 Although systemic haemodynamic parameters did not vary between the dietary groups in our study, we cannot rule out the possibility that there may have been some differences in cerebral haemodynamic status between the dietary groups. Although we did not observe any differences in the pattern of the peri-ischaemic LDF response in the parietal cortex or at the individual time points evaluated during and after ischaemia between the dietary groups, we did see a significantly higher mean ischaemic parietal cortical LDF signal for the soy-rich diet group compared with the soy-free diet group. This is in contrast to other studies that have reported no differences in cortical LDF between rats fed soy-based and isoflavone-free diets in transient (90 min) and permanent (24 h) focal stroke models.34-36 However, LDF signal measures only relative changes in cortical perfusion rather than absolute CBF in the cortex and other brain regions. Therefore, LDF may not be sensitive enough to determine whether soy-based dietary phytoestrogens have an effect on peri-ischaemic preservation of regional CBF in ischaemic brain and may partly explain the differences between our study and others regarding cortical LDF. Future studies using more quantitative measures of regional CBF will need to be carried out to determine whether soy-based dietary phytoestrogens have sufficient cerebrovascular effects in ischaemic brain to potentially confound acute experimental outcomes relative to infarct volumes.
Although our group sizes are small, we felt that the pattern of our initial findings on body weight gain, serum 17β-oestradiol levels, uterine response as well as acute infarct volume outcomes in OVX rats raised important issues relative to dietary choices in laboratory animals that would be of immediate interest to the laboratory animal and scientific communities. Using the data from this pilot study, we have performed a post-test power analysis. Based on these analyses, 17–21 rats and 7–36 rats would be needed in this model to detect significant differences in mm3 and % of volume of contralateral structure, respectively, in the cortex (n = 21 for mm3, n = 13 for %), striatum (n = 17 for mm3, n = 7 for %) and hemisphere (n = 20 for mm3, n = 36 for %). Future studies from our laboratory will expand upon this initial report by assessing the impact of the presence or absence of soy-based dietary phytoestrogens in OVX rats treated with either vehicle or a known protective oestrogen dose on acute infarct volume outcomes.
In summary, dietary soy at the levels tested did not appear to alter acute infarct volumes in OVX rat brain 22 h following transient MCAO in this initial pilot study but preliminary results suggest that more studies with larger groups are needed as well as using oestrogen administration as a positive control. In addition, the impact of soy-derived dietary phytoestrogens still remains to be examined on functional outcomes and at more chronic endpoints as well as in other oestrogen-sensitive brain injury models. Lastly, more detailed pathology will need to be done to further characterize damage at the cellular level and to evaluate effects of dietary soy on specific cell types. Future studies evaluating different levels, types and sources of dietary phytoestrogens in oestrogen-sensitive stroke and other brain injury models are needed if laboratory animal clinicians, animal facility managers and investigators are to make informed choices regarding diets in these models.
Footnotes
Acknowledgements
This study was supported by a grant from the Oregon Health and Science University's Center of Women's Health, Portland, OR, USA. Rats and experimental diets were provided by Harlan Teklad. The authors would like to thank Barbara Mickelson, Dave Johnson, Terry Burns-Heffner and Chuck Benton at Harlan Teklad in Madison, WI for assistance with diet selection and analysis.