
Abstract
Changes in clinical pathology parameters, particularly those related to blood coagulation, were examined throughout the gestation period in New Zealand White rabbits. As compared with the non-pregnant group, the following major changes were observed in the pregnant group. For blood coagulation-related parameters, platelets increased progressively and fibrinogen increased slightly from organogenesis, prothrombin time was significantly prolonged during organogenesis and shortened in the late fetal growth stage, activated partial thromboplastin time was significantly prolonged during the fetal growth stage, and antithrombin III increased during and after late organogenesis. Such changes in blood coagulation-related parameters during the later stages of gestation seem to be physiological responses in preparation for protecting against excessive haemorrhage or haemostasis at parturition. For the other haematological and blood chemical parameters as well as progesterone, red blood cell counts, haemoglobin and haematocrit began to decrease during organogenesis and continued to decrease thereafter. Reticulocyte counts significantly increased during organogenesis and decreased thereafter. White blood cell parameters, except for neutrophils, showed significant decreases during the fetal growth stage. Serum progesterone concentration reached its highest level early in organogenesis and decreased thereafter. Total protein, albumin, glucose, cholesterol, calcium, blood urea nitrogen and creatinine decreased significantly during the middle and/or late periods of gestation. In conclusion, the data obtained from the present study can be used as background data for effective evaluation of reproductive toxicology in rabbits, and pregnant rabbits may serve as models of pregnant women in research pertaining to clinical pathology and gestation.
It is generally acknowledged that gestation induces progressive alterations in several haematology and blood biochemistry parameters in laboratory animals. Therefore meaningful interpretation of data from toxicology studies using pregnant animals becomes difficult without the benefit of historical control data. In addition, to precisely assess the fetotoxicity of the compound administered, it is necessary to take the secondary effects of maternal toxicity on the fetus into consideration.
Pregnant rabbits of the New Zealand White strain (Kbl:NZW) are frequently and widely used in embryo-fetal development toxicity (teratology) studies. However, there is a paucity of data pertaining to haematology and blood biochemistry parameters in pregnant rabbits.1,2 Particularly, there are few detailed reference studies on the changes in blood coagulation-related parameters during gestation. This is important as it has been reported that disorders of the blood coagulation system tend to occur in women in the late stage of pregnancy.3–7
This study was performed to clarify how haematology and blood biochemistry parameters, especially those related to blood coagulation, change during the course of gestation in the rabbit and how well pregnant rabbits may serve as models of pregnant women in research pertaining to clinical pathology and gestation.
Materials and Methods
Animals
Thirty-five female rabbits of the New Zealand White strain (Kbl:NZW; Kitayama Labs, Co, Ltd, Nagano, Japan) at five or seven months of age were used in this study. The animals were maintained individually in aluminium cages in an animal room under controlled conditions (temperature: 22.5 ± 3.5°C; relative humidity: 50 ± 20%; air ventilation: 10-15 times per hour; artificial lighting: 12 h per day), and were allowed free access to diet (RC4; Oriental Yeast Co, Ltd, Tokyo, Japan) and to water via an automatic water supply system (Fuji Water Union, Shizuoka, Japan). After a one-week acclimatization period, 19 females that were judged to be in oestrus were housed together with male rabbits from the same colony in circles (650 mm in diameter × 500 mm in height) for mating on a 1:1 basis. The day when copulation was confirmed was regarded as gestational day 0 (GD 0). The 19 copulated animals were assigned to the pregnant (P) group and the remaining 16 animals that were not subjected to mating were assigned to the non-pregnant (NP) group. Furthermore, from the standpoint of animal welfare, the P and NP groups were subdivided into two subgroups each (Figure 1) in order to reduce stress by blood sampling. As a result of statistical analysis performed on all parameters between the two subgroups in each P group and NP group on the first blood sampling day (GD 0 [mating day] versus GD 6 [one day before implantation]), there were significant but slight differences detected sporadically in several parameters but there were no significant differences in blood coagulation-related parameters, which are the main concern of this study. Therefore, the two subgroups were merged and treated as one group.
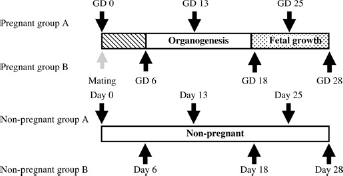
Experimental design, ↑ or ↓: blood sampling point; GD: gestational day; organogenesis: the period of organogenesis; fetal growth: the period of fetal growth
All animals were euthanized as scheduled on GD 28 by exsanguination from the abdominal aorta under pentobarbital anaesthesia.
The experimental procedures were conducted according to the Animal Welfare Guidelines of Bozo Research Center Inc, the Standards of the Institutional Laboratory Animal Resources Guide and the European Convention for the Protection of Vertebrate Animals used for Experimental and Other Scientific Purposes.
Body weight and food consumption
During the experimental period, all animals were checked for general condition every day, body weight on GDs 0, 3, 6, 8, 10, 12, 16, 18, 19, 22, 24, 26 and 28, and food consumption from GD 1 to GD 28.
Haematological and blood chemical examinations
Blood sampling was done twice each during the following gestation periods (Figure 1): (1) the period between fertilization of the ovum or ova and implantation (GDs 0 and 6); (2) the period of organogenesis (GDs 13 and 18); and (3) the period of fetal growth (GDs 25 and 28). Blood samples were collected from the auricle artery under sedation using the restrainer.
As shown in Figure 1, to reduce the stress on the animals, blood sampling was done in 10 animals of the P group and eight animals of the NP group on GDs 0, 13 and 25, and in the remaining nine animals of the P group and eight animals of the NP group on GDs 6, 18 and 28.
At each sampling timepoint, 1 mL blood was collected into collecting tubes containing ethylenediaminetetraacetic acid -2K (SB-41: Sysmex Corporation) and subjected to measurement or calculation of haematological parameters (red blood cells [RBC], haemoglobin [Hb], haematocrit [Hct], mean cell volume [MCV], mean cell haemoglobin [MCH], mean corpuscular haemoglobin concentration [MCHC], reticulocyte, platelets, white blood cells [WBC], differential leukocyte count and absolute value) using an ADVIA® 120 Haematology System (Siemens Healthcare Diagnosis, NY, USA). In addition, a blood sample (1.8 mL) collected into tubes containing 3.8 w/v% sodium citrate (Venoject® 2, VP-C052K, Terumo Corp Co, Ltd, Tokyo, Japan) was centrifuged at approximately 1600 ×
Statistical analysis
The means±standard deviations were calculated for the parameters examined in each group, and were first analysed for homogeneity of variance by the
Results
Changes in body weight and food consumption
There were no abnormal clinical signs observed in any animal throughout the gestation period. Body weight was significantly higher in the P group than in the NP group on GDs 6, 12, 18, 24 and 28 (Table 1). Although food consumption in the P group was higher than that in the NP group during early pregnancy, it was significantly lower on GDs 25 and 28 (Table 1). In comparison with that of the NP group, food consumption of the P group was similar until GD 18, but showed a 30% decrease on GD 28.
Changes in body weights and daily food consumptions in pregnant (P) and non-pregnant (NP) rabbits
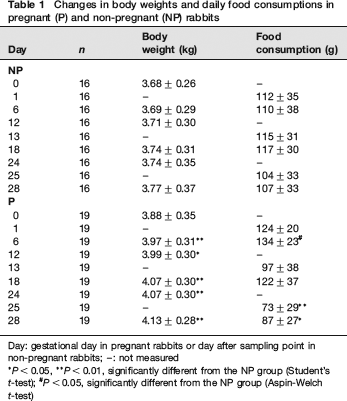
Day: gestational day in pregnant rabbits or day after sampling point in non-pregnant rabbits; -: not measured
Changes in blood coagulation-related parameters
Changes in blood coagulation-related parameters are shown in Table 2 and Figure 2. In the P group, prolongation of APTT was observed on and after GD 18 and it was remarkable on GDs 25 and 28. PT prolonged significantly on GDs 13 and 18, but it shortened on GD 28. ATIII showed significantly higher values on and after GD 18. Platelets showed significantly higher values on GDs 18 and 25. Fibrinogen showed slightly higher values in the P group than in the NP group on and after GD 13, although there were no significant differences between the two groups.
Changes in blood coagulation-related parameters in pregnant (P) and non-pregnant (NP) rabbits
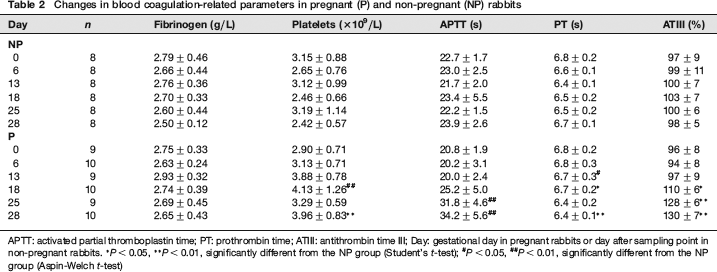
APTT: activated partial thromboplastin time; PT: prothrombin time; ATIII: antithrombin time III; Day: gestational day in pregnant rabbits or day after sampling point in non-pregnant rabbits.
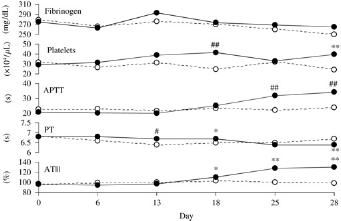
Changes in blood coagulation-related parameters in pregnant and non-pregnant rabbits. Day: gestational day in pregnant rabbits or day after sampling point in non-pregnant rabbits. Pregnant group (•); Non-pregnant group (○). *
Serum progesterone concentration
Serum progesterone concentration was markedly higher in the P group than in the NP group throughout the gestation period (Table 3). The concentration in the P group reached its highest level on GD 13, decreased on GD 18 and maintained a similar level until GD 28, although there were no significant differences observed among all pregnant groups.
Changes in serum progesterone concentrations in pregnant (P) and non-pregnant (NP) rabbits

Day: gestational day
Haematological findings
Sequential changes of RBC parameters are shown in Table 4a. In the P group, RBC, Hb and Hct decreased from GD 18, and they were significantly lower than those in the NP group. On the other hand, reticulocytes in the P group increased about 50% from GD 13 to GD 18, being significantly higher than those in the NP group. However, they became significantly lower than those in the NP group on GD 28. MCV, MCH and MCHC showed almost no significant changes throughout the gestation period.
Changes in red blood cell (RBC) parameters in pregnant (P) and non-pregnant (NP) rabbits
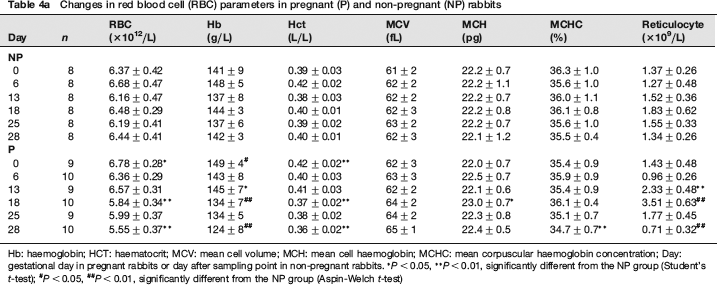
Hb: haemoglobin; HCT: haematocrit; MCV: mean cell volume; MCH: mean cell haemoglobin; MCHC: mean corpuscular haemoglobin concentration; Day: gestational day in pregnant rabbits or day after sampling point in non-pregnant rabbits.
Sequential changes of WBC parameters are shown in Table 4b. The parameters, except for neutrophils, showed significantly lower values in the P group than in the NP group on GDs 25 and/or 28. There were no significant differences in neutrophils between the P and NP groups throughout the gestation period. Lymphocytes were significantly higher in the P group than in the NP group on GD 13.
Changes in white blood cell (WBC) parameters in pregnant (P) and non-pregnant (NP) rabbits
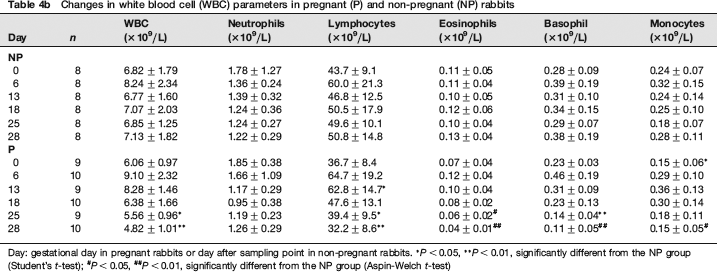
Day: gestational day in pregnant rabbits or day after sampling point in non-pregnant rabbits.
Blood biochemical findings
Sequential changes in blood chemistry are shown in Tables 5a and 5b. As compared with the NP group, total protein and albumin were significantly lower in the P group on and after GD 18. Similar sequential changes were observed in glucose and cholesterol. On the other hand, in the P group, triglyceride began to increase significantly on GD 13, reached the highest value on GD 18, and decreased markedly on GDs 25 and 28. As for electrolytes, the P group showed significantly lower calcium values on GDs 25 and 28 and significantly higher phosphorus values on GDs 13 and 25. In addition, as compared with the NP group, ALP and ALT activities in the P group were significantly lower on and after GD 18 and on GD 28, respectively. BUN and creatinine showed similar sequential changes to that of ALP activity. AST and LD activities in the P group were significantly higher on GD 28.
Changes in blood biochemical parameters in pregnant (P) and non-pregnant (NP) rabbits
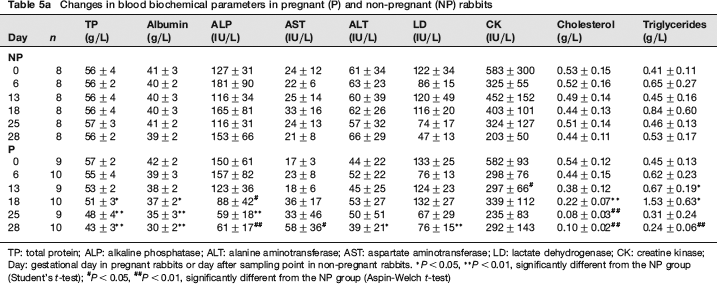
TP: total protein; ALP: alkaline phosphatase; ALT: alanine aminotransferase; AST: aspartate aminotransferase; LD: lactate dehydrogenase; CK: creatine kinase; Day: gestational day in pregnant rabbits or day after sampling point in non-pregnant rabbits.
Changes in blood biochemistry parameters in pregnant (P) and non-pregnant (NP) rabbits (cont'd
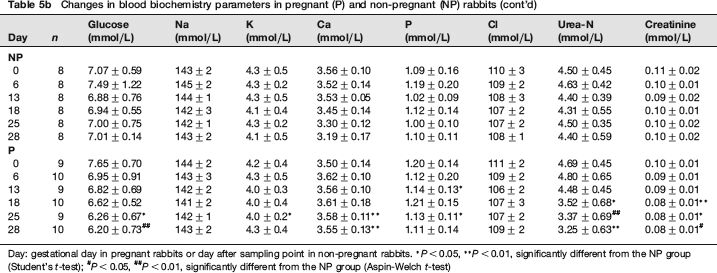
Day: gestational day in pregnant rabbits or day after sampling point in non-pregnant rabbits.
Discussion
In the present study, changes in haematology and blood biochemistry parameters, particularly those related to blood coagulation, were examined throughout the course of gestation in New Zealand White rabbits.
As mentioned in the introduction, it is reported that disorders in the blood coagulation system tend to occur during the late period of pregnancy in women. However, there are only a small number of reports of detailed changes in blood coagulation-related parameters in either pregnant women or rabbits during pregnancy. For example, an increase in fibrinogen,6,7 variable changes in platelets5,7,8 and no changes in ATIII
6
have been reported in women during pregnancy. In pregnant rabbits, Wells
The present data confirmed that there were significant changes in many blood coagulation-related parameters in the late period of pregnancy in rabbits. Namely, platelets increased progressively from organogenesis through the stage of fetal growth stage, as they do in pregnant rats. 9
As in the case of pregnant women, fibrinogen showed slightly higher values (but not significant) in pregnant than in non-pregnant rabbits from early organogenesis through the fetal growth stage, while fibrinogen showed significantly higher values in pregnant than in non-pregnant rats. 9 This suggests that there is a species difference in the effect of pregnancy on fibrinogen production.
In addition, PT prolonged during organogenesis and shortened in the last stage of fetal growth in pregnant rabbits while it prolonged throughout the gestation period and recovered to normal value on GD 20 in pregnant rats. 9 PT was slightly shortened during the late period of pregnancy. A similar change was observed in rats and was considered to be one of the physiological responses to protect prolonged bleeding at delivery. 10 Furthermore, the prolongation of APTT was observed during the fetal growth stage in pregnant rabbits, the same as in pregnant rats. 9 ATIII apparently increased from late organogenesis through the fetal growth stage in pregnant rabbits. Prolongation of APTT and a high value of ATIII during fetal organogenesis were observed in rats. 10 Therefore, these changes in blood coagulation-related parameters during the late period of pregnancy seem to be physiological responses in preparation for protecting against excessive haemorrhage or blood coagulation at parturition, or a mechanism to prevent the development of deep tissue thrombosis in dams.
In women under normal pregnancy, increases in fibrinogen and platelets through the gestation period, an increase in prothrombin volume in the first trimester, and a decrease or steady status in prothrombin volume in the third trimester have been reported.4–7 In addition, increases in such factors as VII, VIII, IX, X and XII and a decrease in XI factor have been observed during gestation.
In the present study on pregnant rabbits, as in the case of pregnant women, a slight increase in fibrinogen was observed from organogenesis to fetal growth.
In addition, prolongation of PT in organogenesis as well as shortening of PT and an increase in platelets in late gestation were observed. However, this study did not examine coagulation factors, for which changes were reported in pregnant women as mentioned above, and therefore total similarity in pregnancy-related changes in the blood coagulation system of women and rabbits could not be confirmed. Further studies are needed to clarify this point.
RBC, Hct and Hb began to decrease during organogenesis and decreased progressively thereafter as reported by Wells
In the present study, reticulocytes drastically increased during organogenesis (GD 13 to GD 18) probably because of the increased demand for RBC during this stage, 12 and they decreased thereafter as previously reported.2,15 In addition, the present data demonstrated that serum progesterone concentration reached its highest level on GD 13, suggesting that the increase of serum progesterone concentration might initiate the enhancement of erythropoiesis. 12,16 There are, however, conflicting reports of reticulocyte changes in pregnant women.17,18
WBC parameters, except for neutrophils, showed significant decreases during the late stage of pregnancy in the present study, and this is thought to be brought about by haemodilution. 2 However, it was reported that WBC parameters increased during the whole period of gestation in pregnant women19–21 and rats, 12 and the reason for such a discrepancy between species remains obscure.
Total protein and albumin decreased at the same level as previously reported in pregnant rabbits,
2
rats
22
and women,
23
and this is thought to reflect the increased blood volume. As mentioned above, these changes were thought to be mainly brought about by haemodilution. In addition, glucose and total cholesterol decreased markedly from late organogenesis through the fetal growth stage.
2
Wells
Serum calcium, BUN and creatinine decreased significantly during the fetal growth stage in the present study. It has been reported that women exhibited steady decreases in electrolytes because of electrolyte demands by the fetus during the fetal growth stage. 27
In the present study, ALP activity, one of the indicators of liver function, decreased from the end stage of organogenesis, and a striking decline in ALP activity was observed during the fetal growth stage. Wells
In conclusion, the data obtained from present study can be used as background data for effective evaluation of reproductive toxicology in rabbits. In addition, pregnant rabbits may serve as models of pregnant women in research pertaining to clinical pathology and gestation, although further studies are needed.
Footnotes
Acknowledgement
The authors would like to thank Dr Kunio Doi, Emeritus Professor of the University of Tokyo, for reviewing of this manuscript.