
Abstract
Opportunistic pathogens have become increasingly relevant as the causative agents of clinical disease and pathological lesions in laboratory animals. This study was conducted to evaluate the role of Klebsiella oxytoca as an opportunistic pathogen in laboratory rodents. Therefore, K. oxytoca-induced lesions were studied from 2004 to early 2006 in naturally infected rodent colonies maintained at The Jackson Laboratory (TJL), Bar Harbor, USA, the Animal Research Centre (Tierforschungszentrum, TFZ) of the University of Ulm, Germany and the Central Animal Facility (ZTM) of the Hannover Medical School, Germany. K. oxytoca infections were observed in substrains of C3H/HeJ mice, which carry the Tlr4Lps-d allele; in LEW.1AR1-iddm rats, the latter being prone to diabetes mellitus; in immunodeficient NMRI-Foxn1nu mice; and in mole voles, Ellobius lutescens. The main lesions observed were severe suppurative otitis media, urogenital tract infections and pneumonia. Bacteriological examination revealed K. oxytoca as monocultures in all cases. Clonality analysis performed on strains isolated at the ZTM and TFZ (serotyping, pulse field gel electrophoresis [PFGE], enterobacterial repetitive intergenic consensus (ERIC) polymerase chain reaction, sequencing of 16S rRNA and rpoB genes) revealed that the majority of bacteria belonged to two clones, one in each facility, expressing the capsule type K55 (ZTM) or K72 (TFZ). Two strains, one isolated at the ZTM and one at the TFZ, showed different PFGE and ERIC pattern than all other isolates and both expressed capsule type K35. In conclusion, K. oxytoca is an opportunistic pathogen capable of inducing pathological lesions in different rodent species.
Klebsiella spp. are opportunistic Gram-negative pathogens that can cause community-acquired severe pyogenic pneumonia in humans, with a high mortality rate if left untreated (Carpenter 1990, Prince et al. 1997, Ishida et al. 1998). The vast majority of Klebsiella spp. infections are associated with hospitalization, the urinary tract being the most common affected site. Klebsiella spp. infections in humans are mainly caused by Klebsiella pneumoniae and to a lesser degree by K. oxytoca (Podschun & Ullmann 1998).
Mice and rats experimentally infected with K. pneumoniae serve as models for a variety of diseases including pneumonia, endotoxaemia, sepsis, cystitis and pyolenephritis (Baker 1998). However, reports of naturally occurring infectious diseases in rodents due to Klebsiella spp., especially K. oxytoca, are very infrequent (Schneemilch 1976, Jackson et al. 1980, Boot & Walvoort 1986, Rao et al. 1987).
Due to the elimination of many primary pathogens in experimental rodent colonies over the last decades, the hygienic status of animals used in research has considerably improved. However, by maintaining animals with a well-defined hygienic status (specified pathogen free), opportunistic pathogens have become increasingly relevant as the causative agents of clinical disease and pathological lesions in laboratory animals. The aim of this study was to determine the role of K. oxytoca as an opportunistic pathogen in laboratory rodents. Therefore, K. oxytoca-induced lesions were studied in naturally infected colonies of Mus musculus, Rattus norvegicus and Ellobius lutescens maintained in three different animal facilities, and K. oxytoca strains isolated from these rodents were characterized.
Materials and methods
Study design
From 2004 to early 2006, 92 cases of K. oxytoca infections from The Jackson Laboratory (TJL), Bar Harbor, USA, the Animal Research Centre (Tierforschungszentrum, TFZ) of the University of Ulm, Germany and the Central Animal Facility (ZTM) of the Hannover Medical School, Germany, were analysed for this study. Only animals displaying lesions, from which K. oxytoca was isolated as a monoculture, were included in this study. At the TFZ and ZTM, K. oxytoca isolated from cases as well as K. oxytoca isolated from non-affected animals during routinely performed hygienic monitoring programmes according to the Federation of European Laboratory Animal Science Associatons (FELASA) recommendations (Nicklas et al. 2002) were assembled for further analyses.
Histological and bacteriological examination
Clinically ill animals underwent necropsy in all three facilities. Animals were euthanized by CO2 asphyxiation. Swabs were taken from suppurative lesions and/or affected organs were removed aseptically and one part cultured as described below; the remaining samples were fixed in neutral buffered 4% formalin (TFZ, ZTM) or 10% acid–alcohol formalin (TJL), processed routinely, embedded in paraffin, sectioned at 5–6 µm and stained with haematoxylin and eosin.
Depending on the lesion, organs or swabs were cultured on blood agar (at the TFZ: Mueller-Hinton agar with 5% sheep blood [Merck, Darmstadt, Germany]; at the ZTM: blood agar base no. 2 with 5% defibrinated sheep blood [Oxoid, Wesel, Germany]; at the TJL: Columbia agar with 5% defibrinated sheep blood [Northeast Labs, Waterville, ME, USA]) and Enterobacteriaceae-specific agar (Gassner [ZTM; Oxoid] or MacConkey's agar [TFZ, TJL; Merck or Difco, Sparks, MD, USA, respectively]) for 24 h at 37°C directly or after enrichment in broth medium (thioglycollate [TFZ, ZTM; Oxoid] or tryptose phosphate [TJL, Difco]) at 37°C for up to one week.
Typing of Klebsiella isolates
Klebsiella isolates from clinical affected and unaffected animals were identified by the API20E system (bioMérieux, Marcy l'Etoile, France). Isolates from affected animals at the TFZ and ZTM as well as unaffected controls were also tested for assimilation of ethanolamine, histamine, D-melezitose and DL-3-hydroxybutyrate (Sigma-Aldrich, Munich, Germany) according to Monnet and Freney (1994) and subjected to serotyping and molecular analysis (pulse field gel electrophoresis [PFGE], enterobacterial repetitive intergenic consensus polymerase chain reaction [ERIC-PCR], sequencing of 16S rRNA and rpoB genes). Serotyping and PFGE were performed at the Institute for Infection Medicine, Kiel, ERIC-PCR at the TFZ and sequencing was performed at the ZTM.
Serotyping, PFGE, ERIC-PCR and analysis of clonality
To analyse clonality, strains were K-serotyped, subjected to PFGE and ERIC-PCR was performed. To determine the K-serotypes, the bacteria were grown on Worfel-Ferguson agar (0.025% w/v MgSO4, 0.1% w/v Ka2SO4, 0.2% w/v NaCl, 2% w/v saccharose [Merck], 0.2% w/v yeast extract and 1.5% Bacto agar [BD, Heidelberg, Germany]) for 24 h at 37°C and for an additional 24 h at room temperature to promote capsular production. The K-serotypes of the isolates were determined by the capsular swelling method using K-specific antisera as described elsewhere (Podschun et al. 1993).
PFGE patterns of the strains were determined after restriction of the bacterial DNA with the endonuclease XbaI as described previously (Sahly et al. 2000a). Strains that were indistinguishable, closely related or possibly related according to the Tenover's criteria for the analysis of PFGE pattern (Tenover et al. 1995) and expressing identical K-serotype were regarded as clonal.
K. oxytoca isolates were also typed by ERIC-PCR using the Ready-to-go RAPD Analysis Bead kit (Amersham Pharmacia, Little Chalfont, UK) and ERIC-1R primer (5′-ATG-TAA-GCT-CCT-GGG-GAT-TCA-C-3′) (Granier et al. 2003). PCR was carried out with an initial denaturation step of 5 min at 95°C, 45 amplification cycles of 1 min at 95°C, 1 min at 36°C, 2 min at 72°C followed by a final extension step of 10 min at 72°C.
Sequencing
Portions of 16S rRNA genes of Klebsiella isolates were amplified using primers A12 (5′-AAG-CCT-GAT-GCA-GCC-A-3′) and A13 (5′-TTT-CGC-ACC-TGA-GCG-T-3′) (Granier et al. 2003), the rpoB (RNA polymerase beta-subunit) encoding genes using primers CM81b (5′-TGA-TCA-ACG-CCA-AGC-C-3′) and CM32b (5′-CGG-AAC-GGC-CTG-ACG-TTG-CAT-3′) (Mollet et al. 1997). PCR was carried out using REDExtract-N-AMP-PCR ReadyMix (Sigma-Aldrich) and annealing temperatures were set to 45°C. Amplificates were loaded on a 2% SeaKem agarose gel (Biozym, Hessisch Oldendorf, Germany) containing SYBR Green (Gel Star, 4 µL/100 mL; Biozym), after 35 cycles of the PCR. PCR products were purified using the NucleoSpin Extract II kit (Macherey Nagel, Düren, Germany) and sequenced. Sequence alignments were performed with the Basic Local Alignment Search Tool (BLAST) (Altschul et al. 1990; www.ncbi.nlm.nih.gov/blast/Blast.cgi) and CLUSTAL W (Thompson et al. 1994; www.ebi.ac.uk/clustalw/). Sequences were submitted to GenBank (EF525558-EF525561).
Results
Klebsiella oxytoca infections in laboratory rodents (Mus musculus, Rattus norvegicus, Ellobius lutescens) maintained at the Central Animal Facility, Hannover Medical School (ZTM), the Tierforschungszentrum, University of Ulm (TFZ) and The Jackson Laboratory, Bar Harbor (TJL)
*Otitis media in mice; otitis media and osteolysis of the bulla in a vole
†Subcutanous, intra-abdominal, intrahepatic
Other: Keratoconjunctivitis and Harderian gland adenitis; meningitis; infection of the oral cavity, the maxilla, and salivary glands; lymphadenitis
UGI: urogenital tract infection
C3H/HeJZtm and C3H/HeJ mice carry the Tlr4Lps-d allele
Colony sizes of the affected breeding colonies were: an average of 20 voles at the TFZ, 60–70 C3H/HeJ mice and 70–80 LEW.1AR1-iddm rats at the ZTM.
Gross necropsy and histology
The main lesions observed were otitis media (that extended to cause osteolysis of the bulla in a vole), urogenital tract infections and pneumonia. Besides this, subcutaneous, intra-abdominal and liver abscesses, keratoconjunctivitis, Harderian gland adenitis, meningitis, infections of the oral cavity, maxilla and salivary glands were noted (Table 1, Figure 1). Lesions induced by K. oxytoca showed severe suppuration and extensive necrosis. Processes tended to spread to neighbouring tissue (Figure 1).
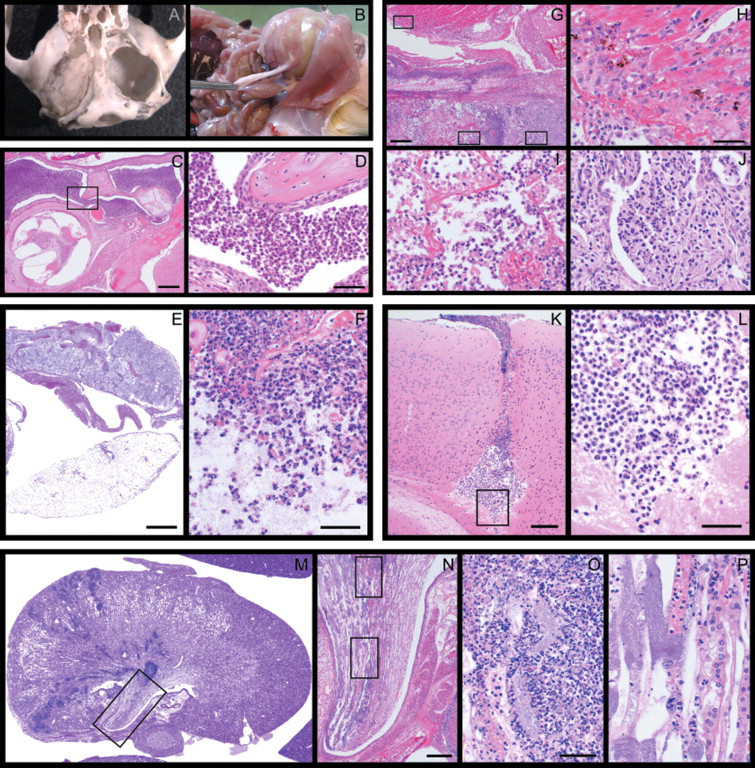
Bacteriological examinations
Klebsiella spp. were cultured from lesions described above, and biochemical identification (API20E and additional assimilation tests performed at the ZTM and TFZ) as well as sequencing of 16S rRNA and rpoB genes revealed K. oxytoca in all cases. Bacteria isolated from lesions, as well as K. oxytoca cultured from non-affected animals examined during routine health monitoring at the TFZ and the ZTM were further typed serologically and by molecular methods. Clonality analysis of the K. oxytoca strains revealed two clones, one in each facility, with two exceptions. At the end of this study, one strain was isolated from a keratoconjunctivitis in an NMRI-Foxn1nu mouse at the ZTM and one strain was isolated from a parotid gland abscess of a vole at the TFZ. Both strains showed different PFGE and ERIC patterns than all other isolates from each facility. Both expressed the capsular type K35.
The clonally indistinguishable strains isolated at the ZTM expressed the capsule type K55 and showed identical PFGE and ERIC-PCR patterns. In contrast, the clonally identical strains isolated at the TFZ showed different PFGE and ERIC-PCR pattern than the ZTM isolates, but were indistinguishable among themselves, and expressed the serotype K72. Sequencing revealed identical 16S rRNA and rpoB gene sequences in all indistinguishable ZTM strains investigated. However, ZTM and TFZ strains showed differences in both gene sequences.
Discussion
In human and veterinary medicine, Klebsiella spp. infections are primarily caused by K. pneumoniae. This applies also for laboratory rabbits and rodents. Here, we describe the identification of K. oxytoca in lesions of three different rodent species. In all lesions described, K. oxytoca was obtained as a monoculture, strongly suggesting its causative role.
Besides the two infections in Ellobius spp. that are not known to have any immune defect, K. oxytoca-induced lesions were restricted to mice and rats displaying characteristic strain-specific features. The two C3H/HeJ substrains are bacterial lipopolysaccharide (LPS) hyporesponsive due to a defect in their toll-like receptor 4 (TLR4) protein (Poltorak et al. 1998). Defective alleles of this receptor are associated with increased susceptibility to Gram-negative infections in humans (Agnese et al. 2002) and mice (Bernheiden et al. 2001, Branger et al. 2004). NMRI-Foxn1nu mice lack a thymus (T-lymphocytes) and are therefore immunodeficient. In addition, these athymic nude mice lack functional eyelashes due to severe follicular dystrophy and might be more susceptible to keratoconjunctivitis (which was seen in these mice) than other mouse strains (Bazille et al. 2001). LEW.1AR1-iddm rats develop diabetes mellitus (Lenzen et al. 2001), a risk factor for Klebsiella spp. induced urogenital tract infection (Chan et al. 1993) and liver abscesses (Lee et al. 2001) in humans.
In addition to predisposing conditions of the host, antibiotic treatment and virulence factors of the bacterium play a role in the pathogenesis of Klebsiella spp. induced lesions. Abnormal colonization with resistant K. pneumoniae has been described after antibiotic treatment of nude rats and mice (Hansen 1995), and antibiotic treatment is associated with increased colonization with Klebsiella spp. in humans as well. In the infected animal rooms at the ZTM, antibiotic treatment was performed with tetracycline (tetracycline hydrochloride 0.7 g/L drinking water for two weeks; bela-pharm, Vechta, Germany) at the end of 2003; however, K. oxytoca was also isolated from clinically ill animals before antibiotic treatment was initiated. However, prior to the use of antibiotics, lesions compatible with infection of K. oxytoca were never observed.
Klebsiella spp. are capable of producing a prominent capsule composed of complex acidic polysaccharides, which are likely major determinants of pathogenicity, at least for K. pneumoniae (Podschun & Ullmann 1998). Based on the structural variability of the capsular polysaccharides, Klebsiella spp. has been classified into 77 serotypes which differ in their pathogenicity and epidemiological relevance (Sahly et al. 2000b). The majority of K. oxytoca isolated from lesions at the ZTM were classified as the K55 serotype. K. oxytoca of the same capsule type was identified as the causative agent of septicaemia in human neonatal wards, suggesting a possible enhanced virulence of this serotype (Morgan et al. 1984, Tullus et al. 1992).
In conclusion, this study describes K. oxytoca as an opportunistic pathogen capable of inducing pathological lesions in three different rodent species. Interestingly, the appearance of cases in TLR4-deficient, diabetic prone or immunodeficient animals and the observation that antibiotic treatment is likely a predisposing factor for abnormal colonization and for induction of lesions show parallels to the situation in human medicine.
Footnotes
Acknowledgements
This work was supported in part by grants from the National Institutes of Health (RR00173 to JPS, CA34196 to The Jackson Laboratory for core facility support) and from the DFG (SFB621, HJH). The authors thank C Elvers and I Köhn for their technical assistance.