
Abstract
Background
Bile acids can act as signalling molecules via various receptors including the nuclear farnesoid X receptor (FXR) and pregnane X receptor (PXR), and the cell surface G-protein-coupled receptor TGR5. The signalling has been implicated in the release of peptide YY (PYY) and glucagon-like peptide 1 (GLP-1), which improves glycaemic control and energy expenditure. We investigated whether morbidly obese subjects have altered postprandial bile acid responses in comparison to normal weight subjects.
Method
Blood samples were taken every 30 min from 0 to 180 min following a 400 kcal test meal. Samples were taken from 12 normal weight subjects with a body mass index (BMI) of 23.2 (2.8) kg/m2 (median [interquartile range (IQR)]) and seven obese patients with a BMI of 47.2 (7.2) kg/m2. Fractionated bile acids were measured on these samples using high-performance liquid chromatography tandem mass spectrometry.
Results
The obese subjects showed a lower postprandial response in total bile acids compared with the normal weight subjects. An increase of 6.4 (5.0) and 2.6 (3.3) μmol/L (median [IQR]) in normal weight and obese subjects was observed, respectively (P = 0.02). The difference was predominantly due to the glycine-conjugated fraction (P = 0.03). There was no difference in the increase of the unconjugated or taurine-conjugated fractions.
Conclusions
The decreased postprandial bile acid response in obese subjects compared with normal weight subjects may partly explain the suboptimal GLP-1 and PYY responses and could affect appetite, glycaemic control and energy expenditure.
Introduction
Bile acids not only form gallstones in obese subjects, but also are increasingly recognized to play an important signalling role in the control of energy balance. Bile acids can act as signalling molecules via different pathways including the nuclear farnesoid X receptor (FXR) and pregnane X receptor (PXR), and by the cell surface G-protein-coupled receptor TGR5. 1 Signal responses are dose dependent and often affected by the degree of hydrophobicity and/or hydroxylation of the particular bile acid. 2
Elevated concentrations of cyclic AMP resulting from bile acids such as deoxycholic acid (DCA) and lithocholic acid stimulating TGR5 results in the production of glucagon-like protein 1 (GLP-1) in enteroendocrine cells, helping to improve glucose homeostasis. 3 Primary bile acid concentrations have been positively correlated to gastric inhibitory polypeptide concentrations in morbid obesity. 4 In humans, DCA has also been shown to stimulate the release of peptide YY from endocrine cells of the distal intestinal mucosa. 5 Bile acids activate thyroid hormone in brown adipose tissue and muscle via the stimulation of type 2 iodothyronine deiodinase (D2), stimulating energy expenditure, and have been associated with insulin sensitization. 6
The majority of bile acids are conjugated either to glycine or taurine moieties in order to become bile salts. Around 95% of bile acids are reabsorbed from the gut in the terminal ileum and transported back to the liver via ‘enterohepatic recirculation’. First-pass hepatic extraction of bile acids is efficient (70–90%) and remains constant in the fasting and digesting state. Therefore, a post-prandial increase in plasma (about 5–15 μmol/L) is seen in the systemic circulation.
The aim of this study was to investigate whether morbidly obese subjects have altered postprandial bile acid responses in comparison with normal weight subjects.
Methods
Subjects and sample collection
EDTA blood samples were taken every 30 min from 0 to 180 min following a 400 kcal test meal; macronutrient contents were as follows: 48.8% carbohydrate, 10.2% protein and 41% fat. Samples were taken from 12 normal weight male subjects (age 21 [1] y, body mass index [BMI] 23.2 [2.8] kg/m2 [median (interquartile range [IQR])]) and seven obese female patients (age 48 [13] y, BMI 47.2 [7.2] kg/m2). All normal weight subjects had a BMI below 26 kg/m2 and all morbidly obese patients were above 43 kg/m2. Informed consent and local ethics committee approval was obtained for this study. After centrifugation, the plasma samples were stored at −80°C until analysed.
Bile acid measurement
These EDTA plasma samples were analysed for fractionated bile acids using high-performance liquid chromatography (HPLC) tandem mass spectrometry. The method was based on that of Tagliocozzi et al. 7 and developed using an Ascentis Express fused core C18 analytical column (150 × 4.6 mm2, 2.7 μm; Sigma-Aldrich Co., Poole, UK) on a Jasco™ LC 2000 HPLC system (Tokyo, Japan) coupled to a triple quadrupole mass spectrometer API 3200™ (Applied Biosystems, Cheshire, UK). Bile acids were quantitated using peak area analysis corrected by comparison to the respective internal standard, glycine, taurine or unconjugated deuterium-labelled DCA. Bile acids analysed were chenodeoxycholic acid, DCA, cholic acid and their respective glycine and taurine conjugates. The method was linear between 0.1 and 10 μmol/L for all bile acids and their conjugates with coefficients of variation ranging from 3.6% to 8.0% at the lower limit of quantitation (0.1 μmol/L). Multiple batches were used; interassay coefficient of variations ranging from 1.5% to 6.8% for these bile acids and their conjugates.
Statistical analysis
Results are reported as median (IQR). Statistical analysis was performed using Analyse-It (Analyse-It, Leeds, UK). Values for the area under the curve (AUC), after correction for baseline time zero values, were calculated with the use of the trapezoidal rule. Data were not normally distributed and a Mann-Whitney U test was used. Significance is shown by a P < 0.05.
Results
There was a significant difference between the BMIs of the normal weight and obese subjects (P = 0.0004), also in the age and sex of the two groups (P < 0.0001 for both). During fasting there was no difference in the total bile acid concentration between the normal weight and the obese subjects (concentration 1.4 [0.4] and 2.5 [3.7] μmol/L, respectively, P = 0.10), or in any of the bile acid fractions analysed. The postprandial response was defined as the maximum increase in concentration attained after the standard meal between 0 and 180 min. Total bile acids response was less for obese subjects than normal weight (increase 6.4 [5.0] and 2.6 [3.3] μmol/L in normal weight and obese, respectively, P = 0.02); this was also true for the AUC of total bile acids (2.8 [2.3] and 1.3 [1.5] μmol/L/min, respectively, P = 0.02). The changes were due to the glycine-conjugated fraction (increase 5.7 [3.5] and 3.1 [2.2] μmol/L for normal weight and obese, respectively, P = 0.028, AUC 2.8 [2.3] and 0.9 [0.8] μmol/L/min, respectively, P = 0.01). There was no difference in the increase of the unconjugated fraction in normal weight (0.08 [0.26] μmol/L) and obese subjects (0.24 [0.84] μmol/L), P = 0.17, nor was the AUC in normal weight (0.01 [0.27] μmol/L/min) and obese subjects (0.16 [0.45] μmol/L/min) different, P = 0.23. The increase in the taurine-conjugated fraction in normal weight (0.46 [0.49] μmol/L) and obese subjects (0.59 [0.58] μmol/L) was not different, P = 0.90, and neither was the increase in AUC for normal weight (0.23 [0.22] μmol/L/min) and obese subjects (0.20 [0.23] μmol/L/min), P = 0.54 (Figure 1).
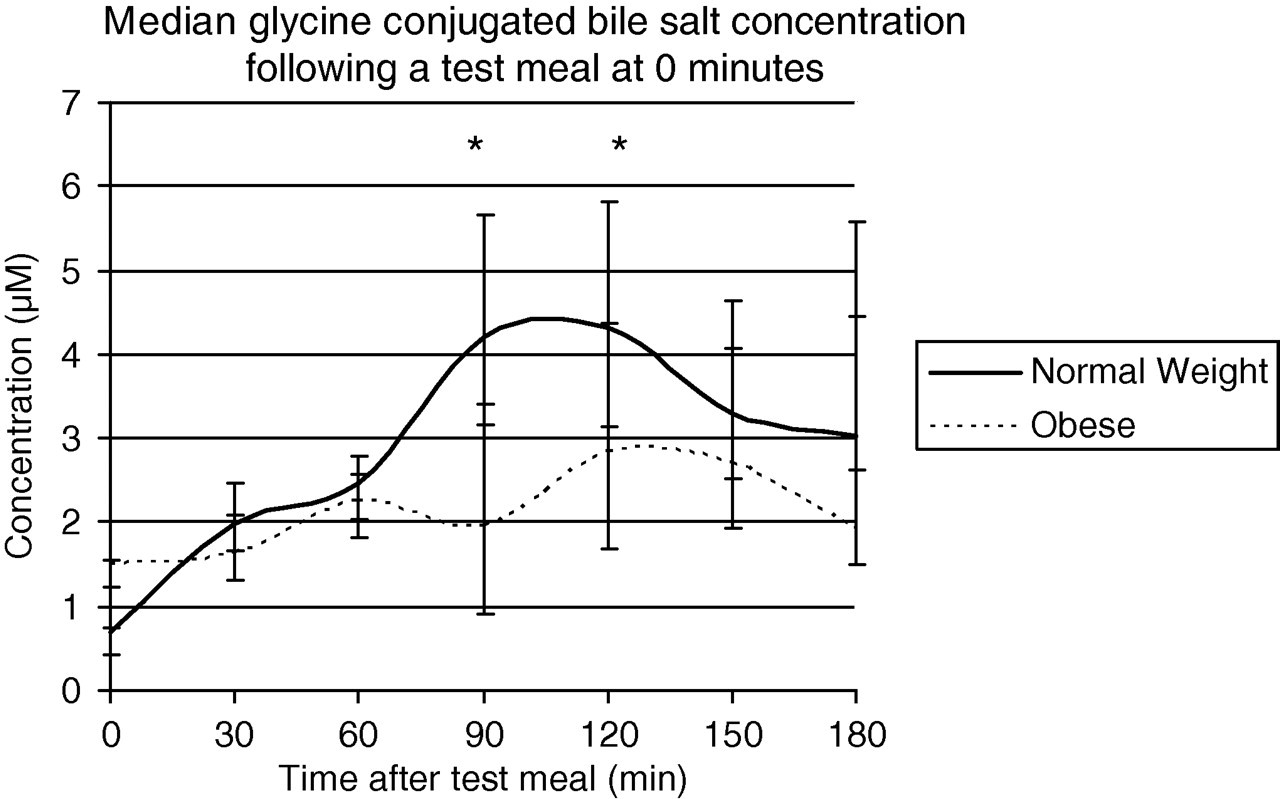
Median concentration of glycine-conjugated bile salts following a test meal at time 0 min in obese and normal weight subjects. Error bars show the interquartile range. The obese subjects have a significantly lower area under the curve for bile salt concentration over a 180-min period after a standard meal. Stars denote that the concentration increase is significantly different in obese subjects at both 90 min (P = 0.013) and 120 min (P = 0.028)
Discussion
We have shown that obese women compared with normal weight men have an attenuated postprandial conjugated bile acid response as a result of a lower response in the glycine-conjugated fraction of bile. Limitations of the study include that the controls were not age or sex matched, although there is no evidence that bile salt metabolism changes with age or is different between the genders.
Bile acids are known to be involved in enteroendocrine cell function and associated with energy expenditure in brown adipose tissue and muscle via stimulation of type 2 iodothyronine deiodinase. 3,5,6 Further work with prospective studies would be required to determine whether attenuated bile acid responses are causative or secondary to obesity.
DECLARATIONS