
Abstract
Background
In human serum, a portion of homocysteine (Hcy) exists as an N-linked form to the ε-amino group of protein lysine residues. N-homocysteinylated proteins differ structurally and functionally from native proteins. The present study strives to develop detection and potential semi-quantification methods for N-homocysteinylated apolipoprotein AI (N-Hcy-apoAI) in human serum.
Methods
Serum treated with or without cysteamine was supplied to isoelectric focusing (IEF) followed by an immunoblot using an anti-apoAI antibody. Cysteamine treatment increased the isoelectric point for N-Hcy-apoAI, but not for unmodified apoAI, due to the presence of -SH group(s) derived from Hcy and the absence of a cysteine residue in the apoAI molecule. N-Hcy-apoAI was semi-quantified from the scanned immunoblot pattern via a computer.
Results
After cysteamine treatment, N-Hcy-apoAI in the serum was identified by IEF at the position with a higher pI value compared with intact apoAI. The reproducibility (between assays) of the semi-quantification method was 19.1% CV (coefficient of variation) for an average ratio 5.9% of N-Hcy-apoAI to the whole apoAI in the serum. Approximately 1.0–7.4% of apoAI was N-homocysteinylated in the serum obtained from 27 healthy subjects. Neither the ratio of N-Hcy-apoAI nor its concentration, calculated by total apoAI concentration, indicated correlation with the so-called total (free and S-linked) Hcy concentration.
Conclusions
We directly found that a portion of apoAI in the serum undergoes homocysteinylation in an N-linkage manner, and used this to develop a potential semi-quantification method for N-Hcy-apoAI.
Introduction
Hyperhomocysteinaemia increases the risk of cardiovascular disease. 1,2 Although the definite mechanism of homocysteine (Hcy) toxicity remains obscure, several mechanisms, including endothelial cell dysfunction, 3,4 smooth muscle cell proliferation activation, 5,6 thrombogenesis 7 and a reduced concentration of plasma high-density lipoproteins (HDL), 8,9 have been reported to explain its role in the development of atherosclerosis. However, the effectiveness of lowering Hcy concentrations by administering folate and vitamins B6 and B12 to reduce the risk of vascular events is controversial. 10–12
N-homocysteinylated protein (N-Hcy-protein), which is observed in human blood and cultured cells, may increase the potential risk of developing cardiovascular disease. 13,14 To prevent Hcy from being used in protein biosynthesis, a portion of Hcy is converted to Hcy thiolactone (HcyT), a cyclic thioester, by methionyl-tRNA synthetase. 15 HcyT reacts with proteins to form an amide bond with the ε-amino group in lysine residues, which leads to denaturation and/or dysfunction of the protein. 13,15 These metabolic reactions occur in human endothelial cells at physiological conditions. 16 N-Hcy-protein has been detected in human myocardium and aortic valve tissues as well as aortic lesions from apolipoprotein(apo)E −/− mice by immunohistochemical staining using a specific anti-N-Hcy-protein antibody. 17 In comparison with the control, staining increased in mice fed a high-methionine diet. Undas et al. 18 have indicated the existence of autoantibodies against N-Hcy-protein with ∼32-fold variation in normal human plasma levels, which were positively correlated with the plasma total Hcy concentrations. Based on the contribution to the immune activation as a neo-self antigen, they hypothesize that autoantibodies against the N-Hcy-protein are important modulators of atherogenesis.
Recently, several methods have been developed to detect protein N-linked Hcy. 19–22 In one study, each of the purchased pure proteins was evaluated for N-linked Hcy content, which varied over an extremely wide range from 0.00006 mol Hcy/mol protein for chicken lysozyme to 0.470 mol Hcy/mol protein for human ferritin. 19 In normal human plasma, about 3–7% of the total Hcy combines with proteins as N-linked Hcy. 19–21 Although it was reported that HDL and low-density lipoproteins (LDL) are also involved in N-homocysteinylation, data for their apolipoproteins such as apolipoprotein (apo) AI, apoAII and apoB were not shown. 22
One of the mechanisms to protect against atherosclerosis by HDL and its major apolipoprotein, apoAI, is reverse cholesterol transport (RCT). In RCT excess cholesterol is extracted from atherosclerotic lesions, particularly macrophages accumulated by cholesteryl esters, and transported to the liver for excretion in the bile. 23 It is hypothesized that N-homocysteinylation of apoAI leads to RCT dysfunction and, consequently, advanced atherosclerosis-prone lesions.
Herein we develop a novel detection and semi-quantification method for N-homocysteinylated apoAI (N-Hcy-apoAI) to investigate the possibility of N-Hcy-apoAI as a potential independent biomarker for atherosclerotic diseases.
Materials and methods
Reagents
Isolation of HDL
HDL (1.063 < d < 1.21 g/mL) was isolated from pooled human serum by ultracentrifugation as described previously, 24 and dialysed against 20 mmol/L Tris-HCl (pH 7.4) containing 1 mmol/L EDTA-2Na. HDL was applied to the Sepharose CL-6B column (1.5 × 97 cm), equilibrated with phosphate-buffered saline to remove contaminated non-HDL lipoproteins, and the purified HDL fraction was concentrated by ultrafiltration.
N-homocysteinylation of apolipoprotein AI
N-homocysteinylation reactions were carried out as described previously. 25 Briefly, 100 μL of HDL suspension (approx. 10 mg protein/mL) was mixed with 25 μL of 6 mmol/L HcyT hydrochloride (dissolved in 20 mmol/L Tris-HCl [pH 10.4] to reach a neutral pH solution), 1.5 μL of 2% NaN3 and 23.5 μL of 20 mmol/L Tris-HCl (pH7.4), followed by incubation overnight at 4°C. The control HDL sample was prepared in a similar manner except the HcyT solution was replaced with saline.
Detection of sulphydryl groups (-SH)
HcyT-treated HDL (approx. 6.7 mg protein/mL) was mixed with the control HDL (approx. 6.7 mg protein/mL) at different rates (10:0, 8:2, 4:6, 3:7, 2:8 and 0:10), so that the mixtures were maintained at the same composition of apolipoproteins and lipids. The mixtures, diluted twice by 20 mmol/L Tris-HCl (pH7.4), were added to equal volumes of 14 mmol/L Tris(2-carboxyethyl)phosphine hydrochloride (Sigma), incubated for 1 h and then incubated with fluorescent-conjugated maleimide (Atto 495 maleimide) for 2 h to label the sulphydryl groups (-SH) of the apolipoproteins. The HDL fractions were separated from the free fluorescent-conjugated maleimide by PD MidiTrap G-25. The HDL fractions (100 μL) were placed into the wells of an ELISA plate and the fluorescent intensities were measured by Corona Multi Microplate Reader MTP-800 (Hitachi High-Technologies, Tokyo, Japan) at excitation and emission wavelengths of 492 and 530 nm, respectively.
Detection of N-Hcy-apoAI
Serum and HcyT-treated HDL were mixed with a cysteamine hydrochloride solution (dissolved in 20 mmol/L Tris-HCl [pH 10.4] to realize a neutral pH) to give a final protein concentration of approximately 21 and 4 μg/μL, respectively, which contained a 6.7-fold amount of cysteamine against protein. This mixture was incubated overnight at 37°C. The cysteamine-treated samples were delipidated with chloroform/methanol/ether (4:2:1, v/v) and washed with ether. The pellets were resuspended in 8 mol/L urea, and approximately 75 and 7.5 μg protein for serum and HcyT-treated HDL, respectively, were applied to isoelectric focusing (IEF) with a pH range from 4.0 to 6.5. Separated proteins were electrophoretically transferred onto a PVDF membrane (Immobilon-P, Millipore, Billerica, MA, USA), incubated with 20 mmol/L tris-buffered saline (TBS) (pH 7.6) containing 5% skim milk and 0.05% Tween-20 (blocking buffer) for 1 h at room temperature, and subsequently washed with TBS containing 0.05% Tween-20 (washing buffer). The membrane was then incubated with anti-apoAI antibody (goat) in TBS containing 0.5% skim milk for 1 h. After washing thrice with the washing buffer, the membrane was incubated with POD-conjugated anti-goat IgG for 1 h. Finally, the membrane was carefully washed, and the bands containing apoAI were visualized using 3,3′-diaminobenzidine tetrahydrochloride and hydrogen peroxide. As shown in Figure 1, apoAI usually does not react with cysteamine due to the absence of a cysteine residue in its molecules. In contrast, N-Hcy-apoAI has a free -SH group derived from Hcy in the apoAI molecule; hence, it does react with cysteamine. Thus, cysteamine treatment influences the isoelectric point (pI) of N-Hcy-apoAI, which changes toward the basic direction, but not unmodified apoAI.
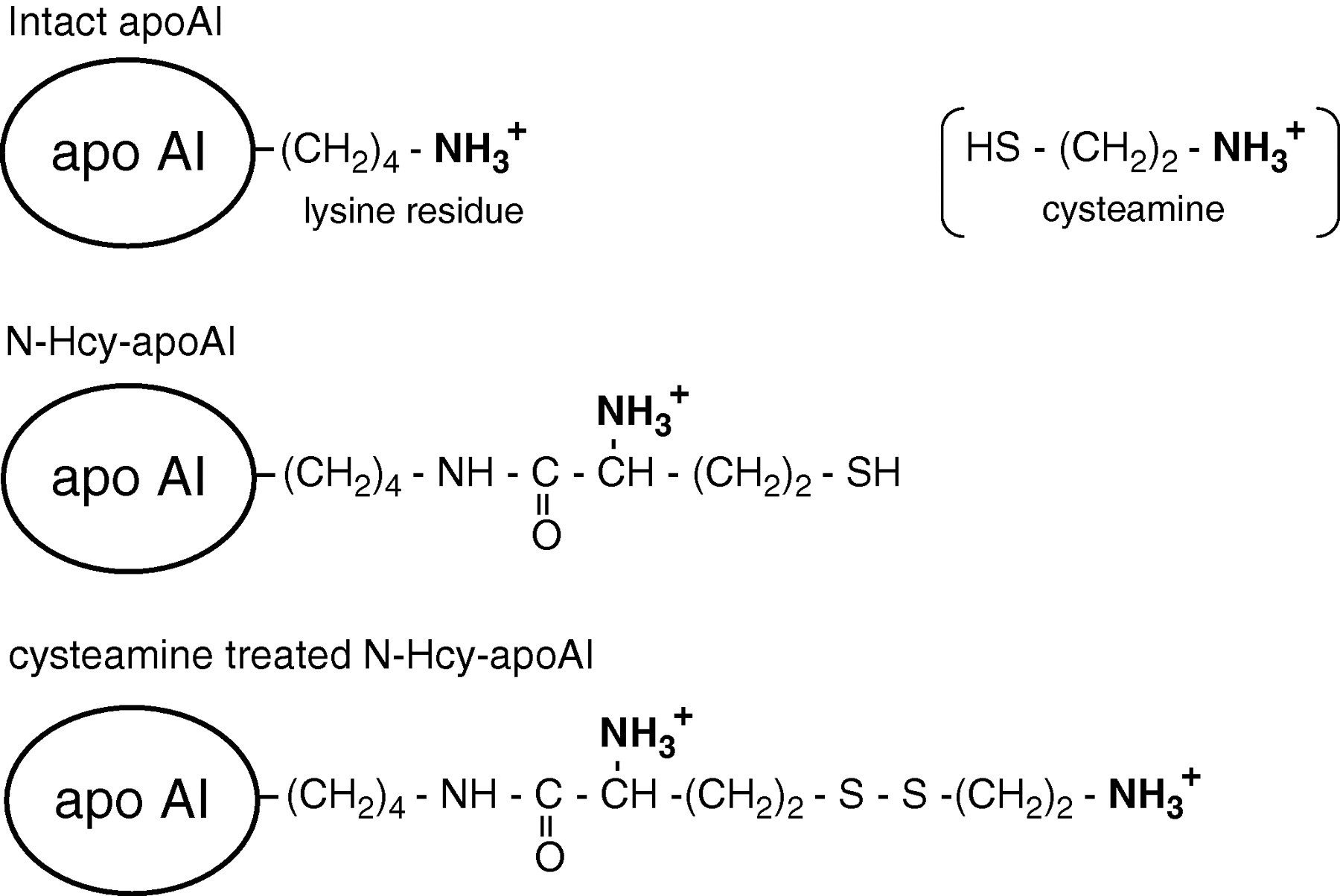
Schematic representation of intact apoAI (top), N-Hcy-apoAI (middle) and cysteamine-treated N-Hcy-apoAI (bottom). N-Hcy-apoAI, but not intact apoAI, reacts with cysteamine due to the presence of the -SH group derived from homocysteine. N-Hcy-apoAI, N-homocysteinylated apoAI
Analysis of HcyT-treated HDL particles
The profiles of HcyT-treated HDL were analysed by agarose-gel electrophoresis using a commercial kit (Rep-Lip; Helena Laboratories, Beaumont, TX, USA). Briefly, 1 μL of N-homocysteinylated HDL (approx. 10 μg protein) was added to the sample wells in a 1% agarose film. After electrophoresis, separated HDL was directly stained with Fat Red 7B. To assess HDL particle size, electrophoresis was carried out using 4–12% non-denaturing gradient gel (Invitrogen Japan) directly stained with CBB R-250.
Two-dimensional electrophoresis
Two-dimensional electrophoresis (2DE) was performed using ZOOM IPGRunner (Invitrogen). HcyT-treated HDL (1 μL) was mixed with 1 μL of ZOOM Carrier Ampholytes 4–7 (Invitrogen), 146 μL of the sample diluent (8 mol/L urea, 2% CHAPS, 0.002% bromophenol blue) and 6 μL of DW, and the mixture was applied to a ZOOM strip (pH 4–7; Invitrogen). After allowing the strip to swell (for at least 1 h), the proteins were focused using the protocol described in the instructions, and the strip was immediately applied to second-dimensional sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE). Finally, the gel was stained with CBB R-250.
Semi-quantification of N-Hcy-apoAI
Blood samples were drawn from apparently healthy volunteers (n = 27) who gave informed consent. The study was approved by our institutional research ethics committee. A pair of serum samples, with and without cysteamine treatment, was determined by IEF followed by immunoblot for apoAI. The bands of the apoAI isoforms were digitally scanned using a Cannon MP610 and analysed with a CS Analyzer (ATTO, Japan). To calculate the ratio of N-Hcy-apoAI to total apoAI, we defined N-Hcy-apoAI as the band observed at the position that is one charge basic compared with intact apoAI. Actually, the ratio of N-Hcy-apoAI was calculated by subtracting the ratio obtained without cysteamine treatment from that with cysteamine treatment, since a non-specific band was occasionally observed at that position without cysteamine treatment.
Results
Detection of N-Hcy-apoAI in normal human serum
The profile of apoAI isoforms in normal human serum was analysed by IEF and a subsequent immunoblot using the anti-apoAI antibody. Cysteamine treatment of the serum resulted in an increase or the appearance of more basic apoAI isoforms compared with the intact apoAI band (Figure 2). The change in the pI values of the apoAI isoforms observed in the serum treated with cysteamine was equivalent to one and two charges toward the basic direction, indicating that part of the apoAI may include -SH group(s) in the molecules.
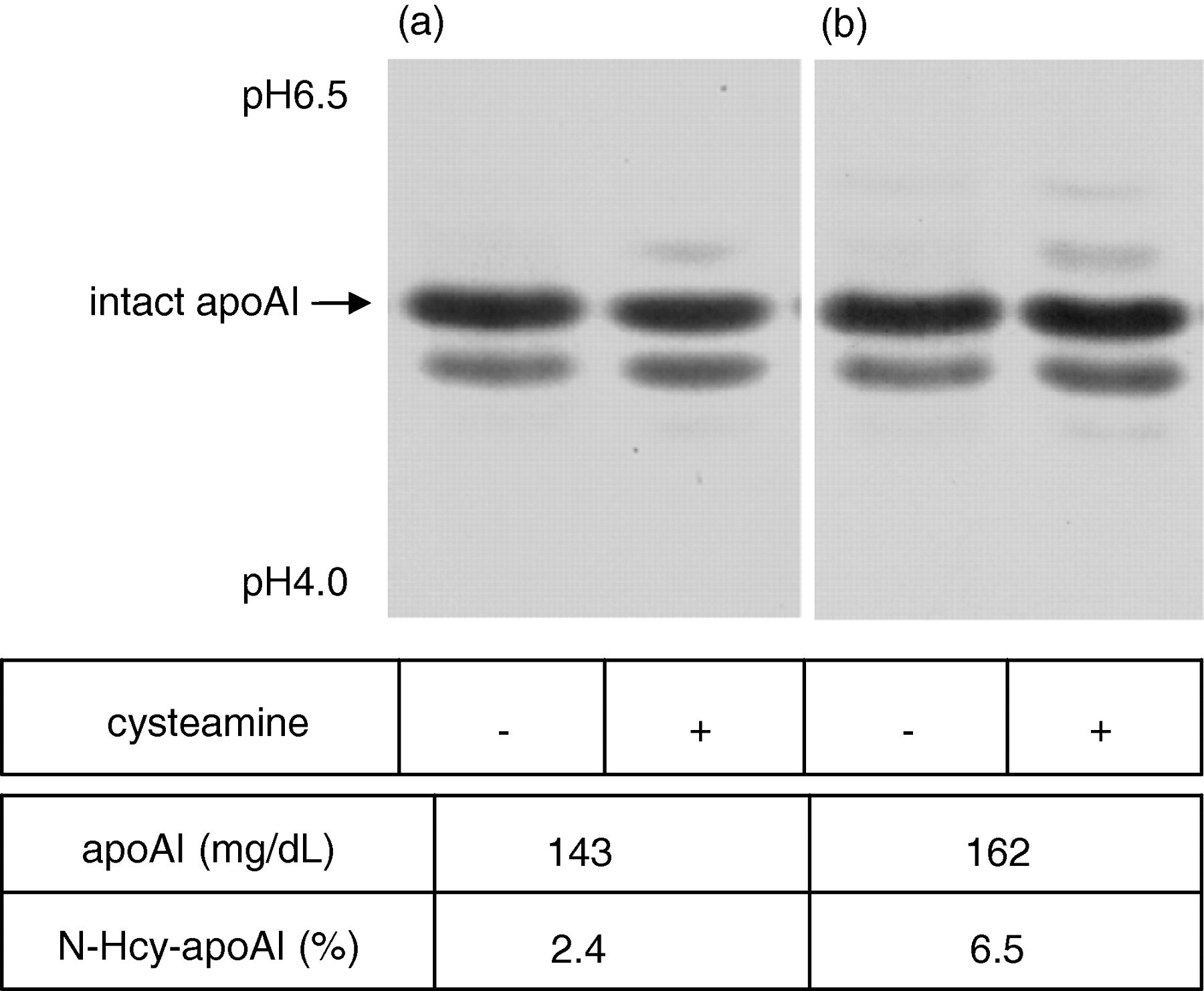
Detection of N-homocysteinylated apoAI (N-Hcy-apoAI) in normal human serum. Serum sample obtained from two individuals (a and b) with (+) and without (−) cysteamine treatment were analysed by isoelectric focusing followed by an immunoblot using an anti-apoAI antibody for the apoAI isoforms. The concentration of total apoAI and the ratio of N-Hcy-apoAI for the serum (a) and (b) were also indicated
Synthesis of N-Hcy-apoAI by HcyT treatment of HDL
Prior to cysteamine treatment, HDL was incubated with or without HcyT, and the profile of the apoAI isoforms was analysed by IEF and a subsequent immunoblot using the anti-apoAI antibody. Incubated HDLs without HcyT had similar patterns as those of normal serum samples both treated and untreated with cysteamine, but the preincubation with HcyT followed by cysteamine treatment induced at least five new apoAI isoforms toward the basic direction, reflecting plural N-linked Hcy in one apoAI molecule (Figure 3). The minimum concentration of HcyT for the statistically significant increase of N-Hcy-apoAI in vitro was estimated at 0.25 mmol/L using the present N-homocysteinylation and detected methods; however, the trend of the occurrence of N-homocysteinylation was observed at 0.05–0.10 mmol/L of HcyT on the immunoblot patterns (data not shown).
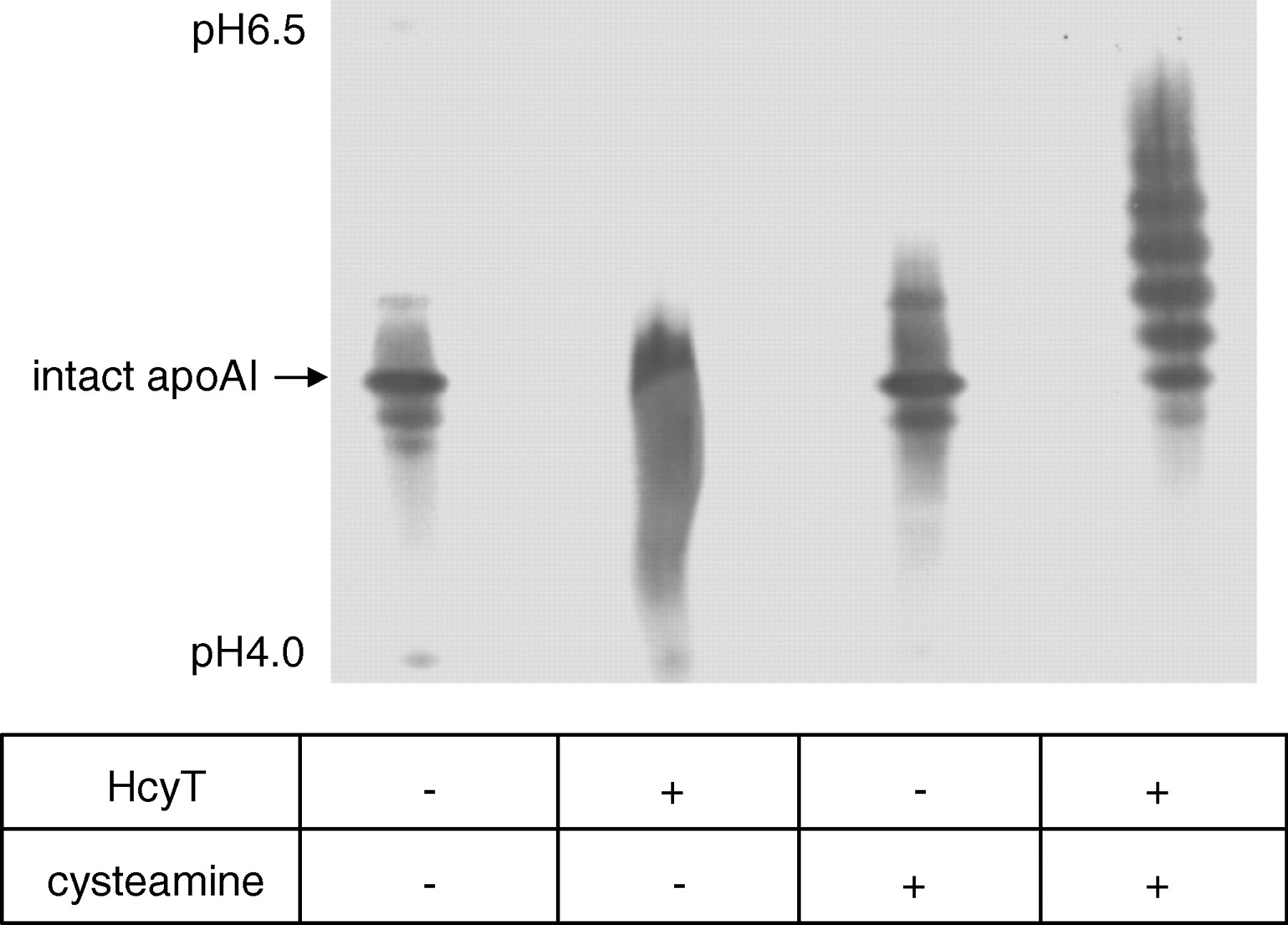
Synthesis of N-Hcy-apoAI by HcyT treatment of HDL. HDL suspension (approx. 1 mg protein) was treated with 1 mmol/L HcyT hydrochloride (HcyT + ). Control HDL sample was prepared in a similar manner except saline was used instead of HcyT solution (HcyT − ). Both HDLs were treated with (+) or without (−) cysteamine followed by isoelectric focusing. ApoAI isoforms were visualized by an immunoblot using anti-apoAI antibody. N-Hcy-apoAI, N-homocysteinylated apoAI; HcyT, homocysteine thiolactone
To confirm that the new apoAI isoforms were formed by N-linked homocysteinylation, the isoforms were subsequently visualized by cysteamine treatment; HDL treated with HcyT was mixed with untreated HDL at different ratios (10:0, 8:2, 4:6, 3:7, 2:8 and 0:10). Consequently, each mixture intrinsically included the same number of original cysteine residues. The -SH groups in the mixtures were then labelled by fluorescent-labelled maleimide, and detected at excitation and emission wavelengths of 492 and 530 nm, respectively. The fluorescent intensity increased in proportion with the concentration of HDL treated with HcyT, indicating the existence of newly generated -SH groups due to N-linked homocysteinylation by HcyT treatment, but N-linked homocysteinylation may have occurred in all of the apolipoproteins, including apoAI in HDL (Figure 4).
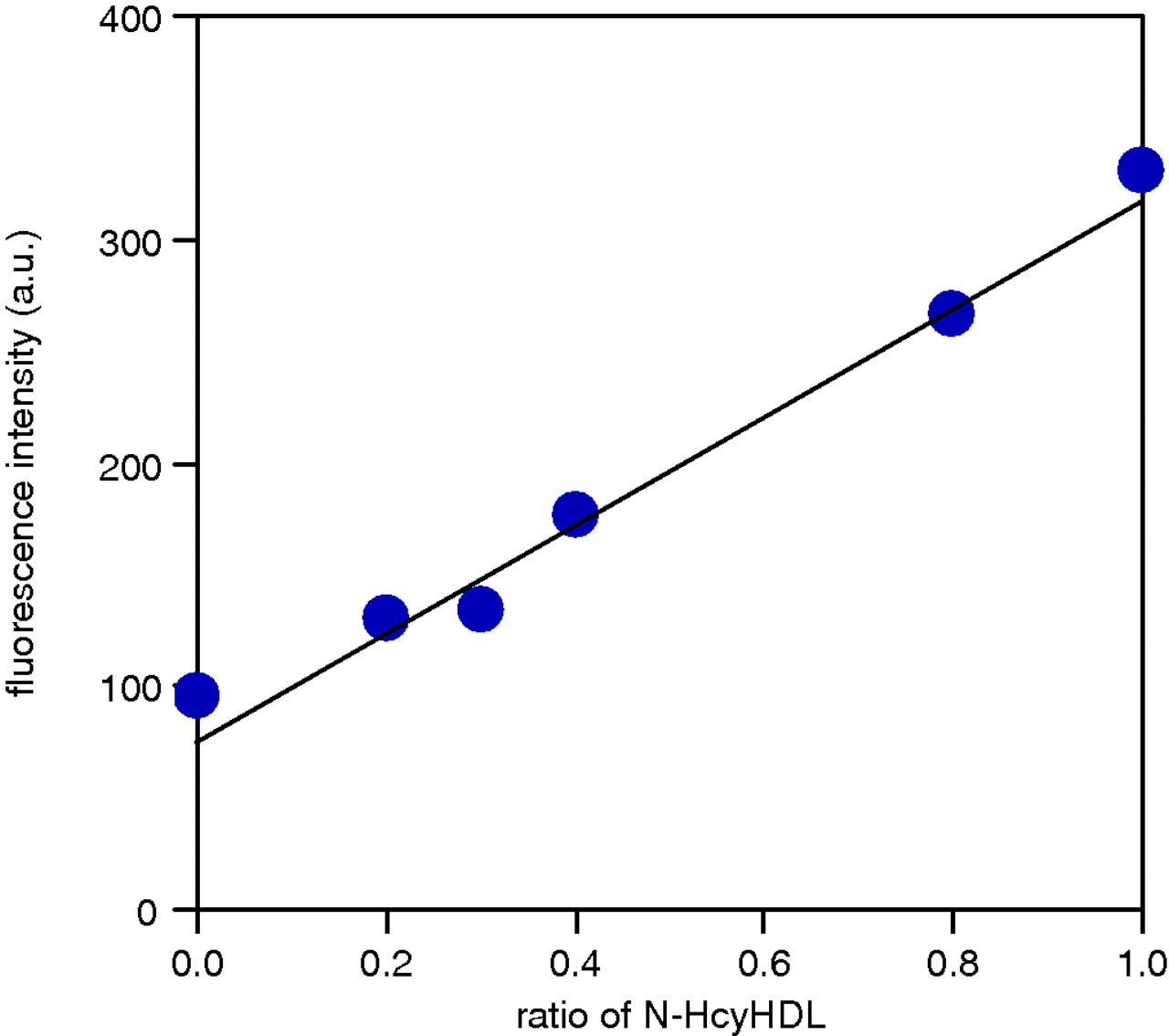
Detection of newly generated -SH groups due to N-linked homocysteinylation by HcyT treatment. HDL treated with HcyT was mixed with untreated HDL at different ratios, and the -SH groups in the mixtures were labelled by fluorescent-labelled maleimide. Fluorescent intensities of the HDL fractions, which were separated from the free fluorescent-conjugated maleimide by PD MidiTrap G-25, were measured. HcyT, homocysteine thiolactone; N-Hcy HDL, N-homocysteinylated HDL
Characterization of HcyT-treated HDL
Although the particle sizes of HcyT-treated HDL and intact HDL on a native-PAGE were similar (Figure 5a), HcyT-treated HDL moved slightly faster to the anode side on agarose gel electrophoresis (Figure 5b). To confirm that the raised pI values of apoAI in HDL sequentially treated with HcyT and cysteamine were not due to fragmentation caused by the treatments, 2DE was carried out (Figure 5c). ApoAI isoforms with a higher pI value than intact apoAI, which can be regarded as N-Hcy-apoAIs, indicated that N-Hcy-apoAIs and intact apoAI have almost the same molecular weight, suggesting that the raised pI values by both treatments are not due to fragmentation of apoAI.
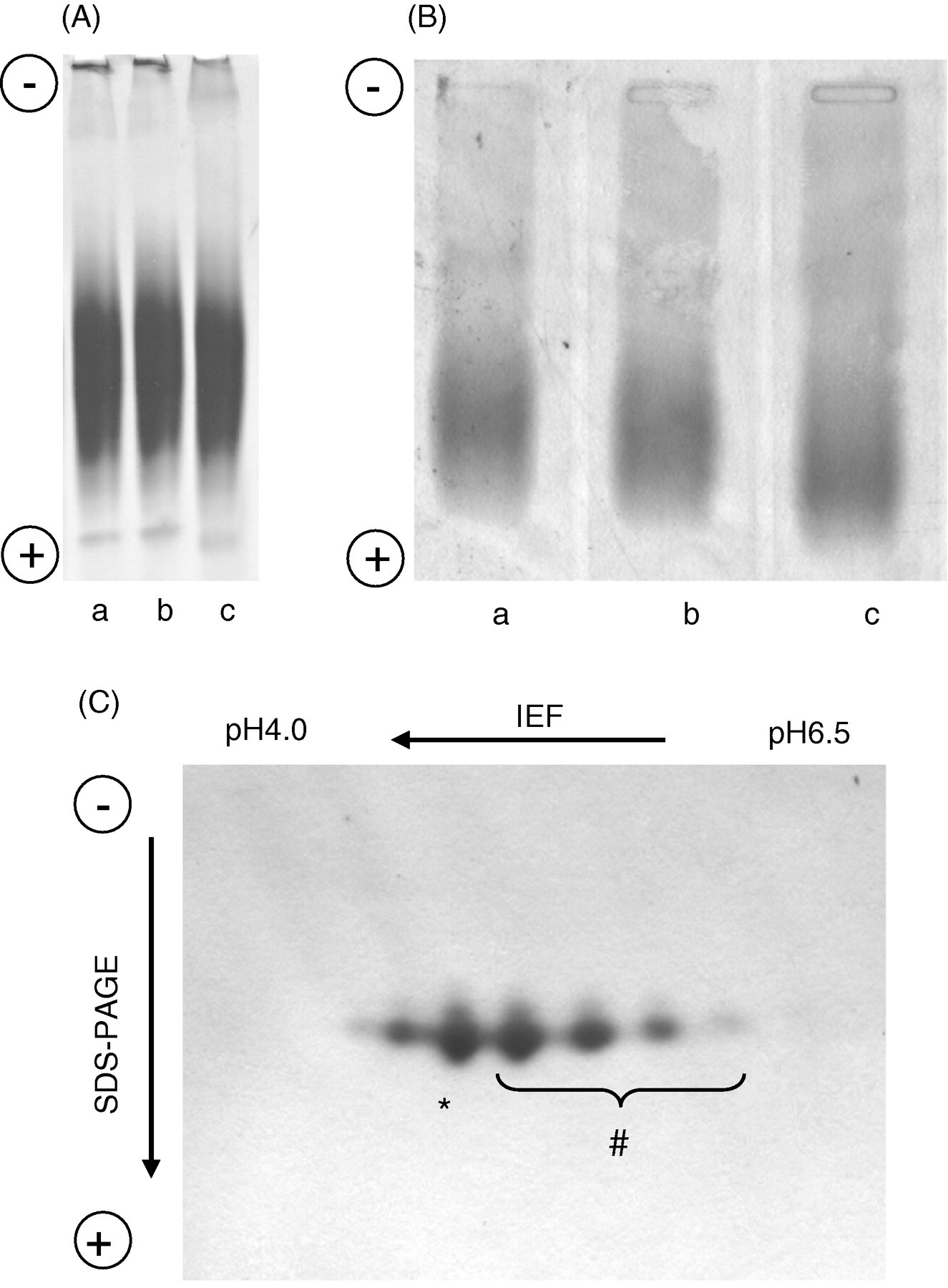
Characterization of HcyT-treated HDL particles. (A and B) Control HDL (a; untreated) and HDL incubated without (b) or with (c) HcyT were analysed by comparing size and charge using non-denaturing polyacrylamide gel electrophoresis followed by CBB R-250 staining (A) and agarose-gel electrophoresis followed by Fat Red 7B staining (B). (C) Effect of HcyT treatment on the molecular size of apoAI in HDL particles. two-dimensional electrophoreses was performed and the gel was stained with CBB R-250. * and # indicate the isoforms of intact apoAI and N-Hcy-apoAIs, respectively. HcyT, homocysteine thiolactone
Concentration of N-Hcy-apoAI in healthy subjects
The N-Hcy-apoAI concentration was calculated by multiplying its ratio (obtained as described in the ‘Materials and methods’) with the total apoAI concentration in the serum obtained by a turbidimetric immunoassay. The reproducibility of the semi-quantification method, obtained by three independent determinations performed in triplicate, was 19.1% (coefficient of variation) for an average ratio 5.9% of N-Hcy-apoAI to the whole apoAI in the normal serum. Although the ratios of N-Hcy-apoAI to the total apoAI varied from 1.0% to 7.4% (from concentrations of 1.2 to 14.5 mg/dL) in 27 healthy subjects, a relatively high correlation (r = 0.673) between N-Hcy-apoAI and the total serum apoAI concentration was observed (Figure 6a).
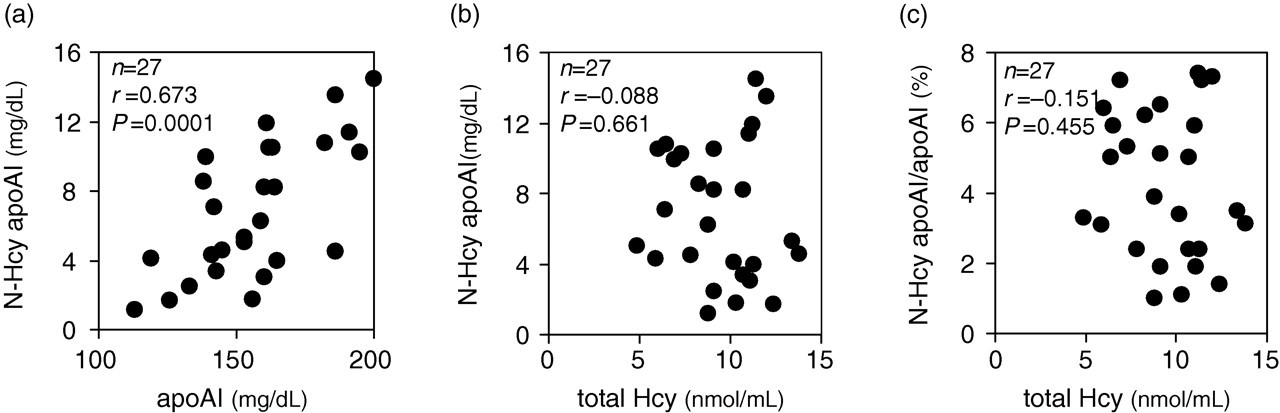
Correlation between N-Hcy-apoAI and total apoAI (a), and total Hcy (b), and between N-Hcy-apoAI/apoAI ratio in the serum and total Hcy (c) in the plasma. The Spearman's correlation coefficient (r) and statistical significance (P) are shown
Additionally, neither the quantity (Figure 6b) nor the ratio against total apoAI (Figure 6c) of N-Hcy-apoAI was correlated with total Hcy concentration, which included all of free Hcy, Hcy homodimer, Hcy-cysteine heterodimer and Hcy S-linked to plasma proteins, but not Hcy N-linked to plasma proteins (r = −0.088 and −0.151, respectively).
Discussion
Because the apoAI molecule lacks a cysteine residue, it does not react with cysteamine, which easily combines with protein through the -SH groups to increase the protein pI value. Therefore, if apoAI profiles on IEF are affected by cysteamine treatment, it is speculated that apoAI acquires -SH group(s) according to N-homocysteinylation. In fact, some of the apoAI isoforms in the IEF patterns were observed at positions of higher pI values after cysteamine treatment of normal serum and HDL.
N-homocysteinylation of HDL with HcyT followed by cysteamine treatment increased the amount of newly developed apoAI isoforms with more basic pI values as well as the large extent of change in the pI values on IEF. These data indicate that a portion of the apoAI molecules include -SH group(s) derived from Hcy, and that this method can determine N-Hcy-apoAI. To confirm the existence of -SH group(s), HDL treated with HcyT was labelled with fluorescent-conjugated maleimide, which specifically binds to -SH groups. The increased fluorescent intensity relative to the increased ratio of HcyT-treated HDL in the mixtures with untreated HDL indicates that -SH groups increase in apolipoproteins, including apoAI in HDL treated with HcyT, although the basic intensity for the sample consisting of 100% untreated HDL is almost derived from the -SH groups in the original cysteine residues.
Under physiological conditions, one or two lysine residue(s) of the 21 lysine residues in the apoAI molecule are involved in N-homocysteinylation (Figure 2). In vitro, protein N-homocysteinylation occurs even at HcyT concentrations as low as 10 nmol/L, 25 which is similar to physiological conditions (between 0 and 35 nmol/L). 26 In contrast, HDL incubated with an extremely high concentration of HcyT, which is approximately a 100,000-fold of human plasma (1 mmol/L), yields an extensive homocysteinylation in which a maximum of five or six lysine residues in apoAI molecle are likely modified by Hcy (Figure 3). To understand the effect of apoAI N-homocysteinylation on its physiological functions, further analysis to identify the N-homocysteinylated site(s) should be important.
Agarose gel electrophoresis (Figure 5b) demonstrated a slightly increased negative charge for HDL treated by HcyT, suggesting that N-homocysteinylation causes an apoAI conformation change in the HDL particle and/or that the ε-amino group(s) of the lysine residue(s) possesses a higher pI value than α-amino group(s) of Hcy residue(s). However, significant changes in the HDL particle size were not observed after HcyT treatment by a native PAGE (Figure 5a). To exclude the possibility that the change in the pI values of apoAI treated with a high concentration of HcyT is due to fragmentation of apoAI, 2DE, IEF for first dimension and subsequent by SDS-PAGE for second dimension were performed. Although the apparent molecular weight did not differ from that of intact apoAI, approximately 28 kDa, N-Hcy-apoAI was observed at positions where the pI values were higher than the intact apoAI as described above (Figure 5c). Treatments by HcyT and cysteamine did not cause fragmentation of apoAI.
The N-Hcy-apoAI concentration, which was calculated from the total apoAI concentration, shows a relatively high correlation (r = 0.673) with total apoAI concentration as a group (Figure 6a). However, the possibility of N-Hcy-apoAI as an independent biomarker for an individual progressive atherosclerosis is undeniable since the N-Hcy-apoAI values widely varied (from 1.8 to 11.9 mg/dL) among the individuals whose total apoAI concentrations in the serum were similar (around 150 mg/dL). In contrast, consistent with the previous report by Jakubowski and co-workers, 27 N-Hcy-apoAI and the total Hcy concentration were not correlated (Figure 6b and c).
The N-homocysteinylation concentrations vary among the types of proteins in the plasma. For example, no N-homocysteinylated transthyretin was found in the plasma obtained from patients with hyperhomocysteinaemia, 28 and 50% of ferritin have been undergone N-homocysteinylation in commercially available purified proteins. 19 Therefore, the maximum N-homocysteinylation ratio in the present study of 7.4% for apoAI is conceivable. Although some methods for total N-Hcy-proteins assay are available, few reports on individual plasma proteins have been published. Thus, to analyse the progression of atherosclerosis-prone legions, determination of N-Hcy-apoAI should provide more beneficial information than that of N-Hcy-proteins.
However, the method herein has limitations as a rigorous quantification assay. First, proapoAI and deamidated apoAI as well as intact apoAI exist in the plasma as the isoforms of apoAI. 29,30 In IEF after treatment with cysteamine, N-Hcy-apoAI at one lysine residue would show a similar isoelectric point with singly deamidated apoAI, which is simultaneously appended by two molecules of N-linked Hcys. Hence, the apparently intact apoAI band after treatment with cysteamine may include not only singly deamidated apoAI modified by one molecule of N-linked Hcy but also doubly deamidated apoAI modified by two molecules of N-linked Hcys. However, maintaining a neutral pH during the incubation with cysteamine would avoid deamidation and cause the aforementioned isoforms, including N-homocysteinylation and deamidation, to be negligible. In the present study, the ratio of N-Hcy-apoAI is defined as the ratio of the apoAI isoform with an increased isoelectric point equivalent to one charge compared with intact apoAI when the serum is treated with cysteamine. Second, it is unclear whether N-Hcy-apoAI has the same reactivity with the antibody as intact apoAI. Actually, modified apoAIs, such as nitration and chlorination of tyrosine residues, often induce higher reactivity than intact apoAI on immunoblot assays. 31 Despite these limitations, the current method could be useful to elucidate whether N-Hcy-apoAI leads to advanced atherosclerosis-prone lesions.
DECLARATIONS