
Abstract
Background
Vitamin D binding protein (DBP) acts as a vitamin D carrier and an actin scavenger. We have previously reported a correlation between serum DBP and lipid parameters in a cystic fibrosis population. In the present study, the relationship between serum DBP, lipoprotein fractions and vitamin D is investigated.
Methods
The presence of DBP in lipoprotein fractions was examined using precipitation, gel permeation chromatography and ultracentrifugation. The association between DBP and lipids was investigated in a cohort study of 211 men. Total and actin-free DBP concentrations were assessed by immunonephelometry and enzyme-linked immunosorbent assay. Serum 25(OH)- and 1.25(OH)2-vitamin D3 were assayed by radioimmunoassay. Total cholesterol, high-density lipoprotein cholesterol (HDL), triglycerides and insulin were measured using routine methods. Low-density lipoprotein-cholesterol (LDL) was calculated according to Friedewald's formula.
Results
DBP was found to be present in very-low-density lipoprotein (VLDL). Gel permeation chromatography revealed a bimodal DBP distribution with a lipid-bound fraction besides the known free fraction. Ultracentrifugation confirmed the presence of DBP and 25(OH)-vitamin D3 in the VLDL particle. Total serum DBP concentration and the actin-bound DBP/DBP ratio correlated significantly with total cholesterol, LDL-cholesterol, triglycerides and albumin. The 25(OH)-vitamin D3/DBP ratio correlated negatively with serum triglyceride concentration and body mass index (BMI). The actin-bound DBP complex was identified with Western blot.
Conclusions
The lipid-bound DBP fraction may be of greater importance than initially thought. The present findings may have clinical consequences in view of the important physiological role of DBP.
Introduction
The functional properties of vitamin D binding protein (DBP) or group-specific component globulin (Gc-globulin) have been derived from both clinical and animal studies. First of all, this member of the albumin superfamily of binding proteins is a passive carrier and an active facilitator of the cellular uptake of vitamin D and its metabolites. 1 The large molar excess of DBP in relation to its major ligand 25(OH)-vitamin D3 opens the possibility to act as a circulating vitamin D reservoir.
Besides the well-described variety of DBP phenotypes (three common alleles, together with numerous other variants) and genotypes, little attention has been paid to the association of this plasma protein with the various lipoprotein fractions. However, this aspect of plasma DBP might be more important than initially thought. 2 As we have reported previously, a correlation between serum DBP concentration and lipid parameters (serum total cholesterol, low-density lipoprotein [LDL]-cholesterol, triglycerides and leptin concentration) was found in cystic fibrosis patients. 3 The distribution of the partially lipophilic DBP in plasma and its potential association with lipoprotein particles and vitamin D has only been investigated to a small degree. Using ultracentrifugation, Haddad et al. analysed upper and lower phases for 25(OH)-vitamin D3, 1.25(OH)2-vitamin D3, cholesterol and DBP. Forty-one percent of 25(OH)-vitamin D3 and >85% of cholesterol was detected at density <1.3 g/mL, whereas nearly all (>95%) 1.25(OH)2-vitamin D3 and DBP was found at density >1.3 g/mL. 4 However, 88% of 25(OH)-vitamin D3 is bound to DBP with high affinity (K a = 5 × 10−8 mmol/L). 2 Ongoing research is still focused on the precise role of DBP in the vitamin D biology.
Secondly, DBP also contains a domain near its C-terminus that facilitates high-affinity binding to globular (G)-actin. In cooperation with gelsolin, DBP takes part in the complex actin scavenging system in the vascular compartment. This mechanism is critical in cytoskeleton dynamics and in the modulation of intravascular thrombosis as DBP exerts an antithrombotic effect. 2,5 Nevertheless, the connection between DBP, lipid metabolism and actin has never been investigated.
In the present study, we explore the relationship between DBP, lipids and vitamin D. In addition, this paper discusses the actin scavenging function of DBP from a new point of view.
Methods
Study population
Male subjects between 71 and 85 y were recruited from the population register of a semi-rural community. Baseline data and detailed inclusion and exclusion criteria were described previously. 6 In brief, subjects with medical conditions or medication use, affecting bone metabolism and subjects with diabetes were excluded as well as subjects taking vitamin D supplements. The hepatic synthesis of DBP is oestrogen dependent and is significantly increased during pregnancy and oestrogen therapy. For that reason, we excluded women from our study population. In contrast to other plasma proteins, DBP maintains stable plasma concentrations throughout life. 2 After exclusion, 211 subjects were included in the present study. Written informed consent was obtained from all participants, together with the medical history.
In order to further investigate the functional integrity of DBP, in particular its interactions with actin and lipids, we collected some serum of patients, who underwent cardiac surgery in the Heart Centre at Ghent University Hospital (Gent, Belgium). Written informed consent was obtained from all participants.
Lipoprotein precipitation
In order to investigate the relationship between DBP and lipoproteins, a heparin-Mn2+ precipitation was performed. Fasting venous blood was centrifuged for eight minutes (3000
Gel permeation chromatography
High-pressure gel permeation chromatography (HPGPC) was performed using a Waters 650E Advanced protein purification system (Waters Corporation, Milford, MA, USA). A 150 μL volume of human serum was injected using the Wisp 712 automatic sampler (Waters Corporation, Milford, MA, USA). For the chromatographic separations, an 8 × 300 mm2 Protein PAK glass 300 SW column (Waters Corporation, Milford, MA, USA) was applied. Phosphate-buffered saline (PBS) 0.1 mol/L, pH 7.3 was used as the mobile phase. The flow rate was 1.0 mL/min. The presence of DBP in the collected fractions was confirmed nephelometrically. 9
Ultracentrifugation
Ultracentrifugation of the pooled sera was carried out with the adjusted Esterbauer method according to Terpstra et al. 10,11 VLDL was removed from serum and DBP was determined in the VLDL fraction and in the infranatant after removal of the VLDL.
Quantification of serum concentrations of total and actin-free DBP
Total serum DBP concentrations were determined by fixed-time immunonephelometry using anti-human DBP polyclonal antibodies (Dako, Glostrup, Denmark) on a BN 100 nephelometer (Dade Behring, Marburg, Germany). 12 DBP standard was purchased from Calbiochem (Merck, Darmstadt, Germany). In a representative subgroup of 88 subjects, actin-free DBP concentrations in serum were determined by an enzyme-linked immunosorbent assay (ELISA) (Kit 034, AntibodyShop, Gentofte, Denmark). Using this assay, microwells were coated with a monoclonal antibody capable of binding human DBP whether or not complexed with actin. Samples were diluted at 1:5000 and applied in duplicate to ELISA plates, each containing eight standard concentrations (calibrators) in duplicate. Coat-bound actin-free DBP was detected with a horseradish peroxidase (HRP)-conjugated monoclonal antibody whose binding to DBP was blocked by the presence of bound actin. This was followed by colour development via incubation with a chromogenic peroxidase substrate containing tetramethylbenzidine. The HRP linked to the bound detection antibody reacted with the substrate to generate a coloured product. The enzymatic reaction was stopped chemically (by addition of H2SO4) and the colour intensity was read at 450 nm in an ELISA reader. Reference values ranged from 1.7 to 6.0 μmol/L, as determined by the manufacturer. The median inter-assay coefficient of variance was 4.2%.
General biochemical determinations
Fasting venous blood was taken, centrifuged and stored at −80°C until analysis. Serum 25(OH)-vitamin D3 and serum 1.25(OH)2-vitamin D3 were measured by a radioimmunoassay (DiaSorin, Stillwater, MN, USA) after chromatographic extraction (C18-OH, Varian Europe, Middelburg, The Netherlands). 13 Triglycerides, 14 total serum cholesterol 15 and high-density lipoprotein-cholesterol (HDL-cholesterol) 16 were assayed in parallel on a Modular P analyser (Roche, Mannheim, Germany) using enzymatic methods. LDL-cholesterol was calculated according to Friedewald's formula. 17 Insulin was measured on a Modular E170 analyser (Roche, Mannheim, Germany). Lipemic index was determined by the difference between the absorbance of the sample, diluted in isotonic saline, at 660 and 700 nm. Body mass index (BMI) was calculated as weight (kg)/(length (m)) 2 .
DBP-phenotype and Western blotting
DBP-isoforms were separated and determined electrophoretically on a non-denaturating 8.5% polyacrylamide gel (30% T, pH 8.8). Five microlitres of serum were loaded on the gel. The gels were run for 90 min at constant voltage of 120 V using 0.75-mm-thick minigels in the BioRad Mini-Protean II electrophoretic system. Gels were stained with Coomassie blue and the DBP-phenotype was determined according to the migrating pattern. 18
Besides the phenotyping, a Western blot analysis was performed. After polyacrylamide gel electrophoresis (PAGE), the gels were pre-equilibrated in transfer buffer prior to electrophoretic transfer (15 min). This procedure facilitates the removal of contaminating electrophoresis buffer salts and neutralization salts. The separated proteins were transferred from the gel, at 340 mA (constant current) for 60 min, to a 0.45 µm nitrocellulose membrane using the BioRad Mini Trans-Blot Electrophoretic Transfer Cell. The blots were then incubated in a quenching buffer for 30 min at 4°C. After washing three times with a PBS solution, the primary antibody (anti-human DBP polyclonal antibody [Dako, Glostrup, Denmark], 1:1000 diluted in probing buffer) was added and the blot was incubated for 16 h at 4°C. After washing with different wash solutions, the blots were added to an affinity-purified secondary goat-anti-rabbit-HRP-conjugated antibody and incubated for four hours at 4°C. Finally, the blots were washed six times with wash solution and the DBP substrate solution was added until the bands were visible (usually 10 min). 19
Intron 8 (TAAA) n -Alu repeat genotypes
Prior to genotyping, genomic DNA was extracted from an ethylene-diamine tetraacetic acid-treated blood sample using a commercially available kit (Qiagen Mini Kit, Hilden, Germany). The (TAAA) n -Alu repeat polymorphism in intron 8 of the DBP-gene was determined by polymerase chain reaction, using primers derived from previous publications 20,21 and Genbank sequence. The forward primer with sequence 5′-CAGCGAGCCAAGATGGCA-3′ was labelled with the fluorophore carboxyfluorescein. The reverse primer with sequence 5′-CCGGAAGTATTCCTCAGTAA-3′ was unlabelled. Fragment length was determined on an ABI PRISM 310 Genescan Analyser (ABI Prism, PE Applied Biosystems, Foster City, CA, USA). The number of TAAA-repeats was calculated from fragment length, and confirmed by sequencing of the amplified fragment.
Statistics
Statistical analyses were performed using MedCalc (MedCalc, Mariakerke, Belgium). Values were expressed as median (interquartile range [IQR]). Differences between two groups were assessed by Mann-Whitney U test or between more than two groups by Kruskall–Wallis test. Spearman correlation coefficients were used to express associations between continuous parameters. Multiple linear regression analysis was performed after log transformation of variables to obtain normal distribution. Statistical significance was considered with a P value of <0.05.
Results
DBP-phenotypic and -genotypic distribution
DBP-phenotypic and -genotypic distributions were in agreement with the Hardy–Weinberg equilibrium, showing 55.0% of individuals with DBP 1-1 (n = 116), 35.5% with DBP 2-1 (n = 75) and 9.5% with DBP 2-2 (n = 20). The 8/8 (TAAA) n -genotype had the highest frequency (54.0%). No significant differences in DBP concentrations among (TAAA) n -Alu genotypes were observed. No significant differences in BMI, cholesterol, HDL-cholesterol, LDL-cholesterol and triglycerides were observed among the DBP-phenotypes or (TAAA) n -genotypes.
Serum 1,25(OH)2-vitamin D3 and DBP concentration in relation to DBP-phenotypes and intron 8 (TAAA) n -Alu repeat genotypes
The total serum DBP concentration in the study population ranged from 4.2 to 14.7 μmol/L. The median serum DBP concentration was 8.4 μmol/L (IQR, 7.3–9.5 μmol/L) in subjects with DBP 1–1, 7.8 µmol/L (IQR, 6.6–9.3 μmol/L) in subjects with DBP 2-1 and 7.2 μmol/L (IQR, 6.2–8.6 μmol/L) in subjects with DBP 2-2 (P < 0.05). No differences in total serum DBP concentrations among (TAAA) n -Alu genotypes were observed (P = 0.11). The 1,25(OH)2-vitamin D3 concentrations were dependent on the DBP-phenotype (P < 0.01), although the 1,25(OH)2-vitamin D3/DBP ratio was comparable between phenotypes. No difference in 1,25(OH)2-vitamin D3 concentrations among (TAAA) n -repeat genotypes was detected (P = 0.101).
Precipitation experiments and HPGPC analysis
Precipitation of the VLDL and LDL fraction resulted in a considerable loss of DBP and 25(OH)-vitamin D3 concentration, indicative of an association between these serum lipoproteins and DBP-vitamin D complexes. For the Lipid Research Clinic Programme method, relative decreases of 31 ± 3% (DBP) and 18 ± 2% (25(OH)-vitamin D3) (n = 5) were obtained. Similar results (relative decreases of 30 ± 2% [DBP]) were observed when the method of Burstein et al. 8 was applied.
Ultracentrifugation proved that DBP and 25(OH)-vitamin D3 were present in the VLDL as well as in the non-VLDL fraction. There was no statistically significant difference in 25(OH)-vitamin D3/DBP between the two obtained fractions (VLDL versus non-VLDL).
Following HPGPC, the concentrations of the various DBP containing fractions were determined by nephelometry. The chromatogram showed a bimodal DBP distribution with a lipid-bound fraction besides the known free fraction. The bound DBP fraction was characterized by an elution volume close to the void volume of the HPGPC column. Figure 1 illustrates the unique molecular mass distribution of serum DBP.
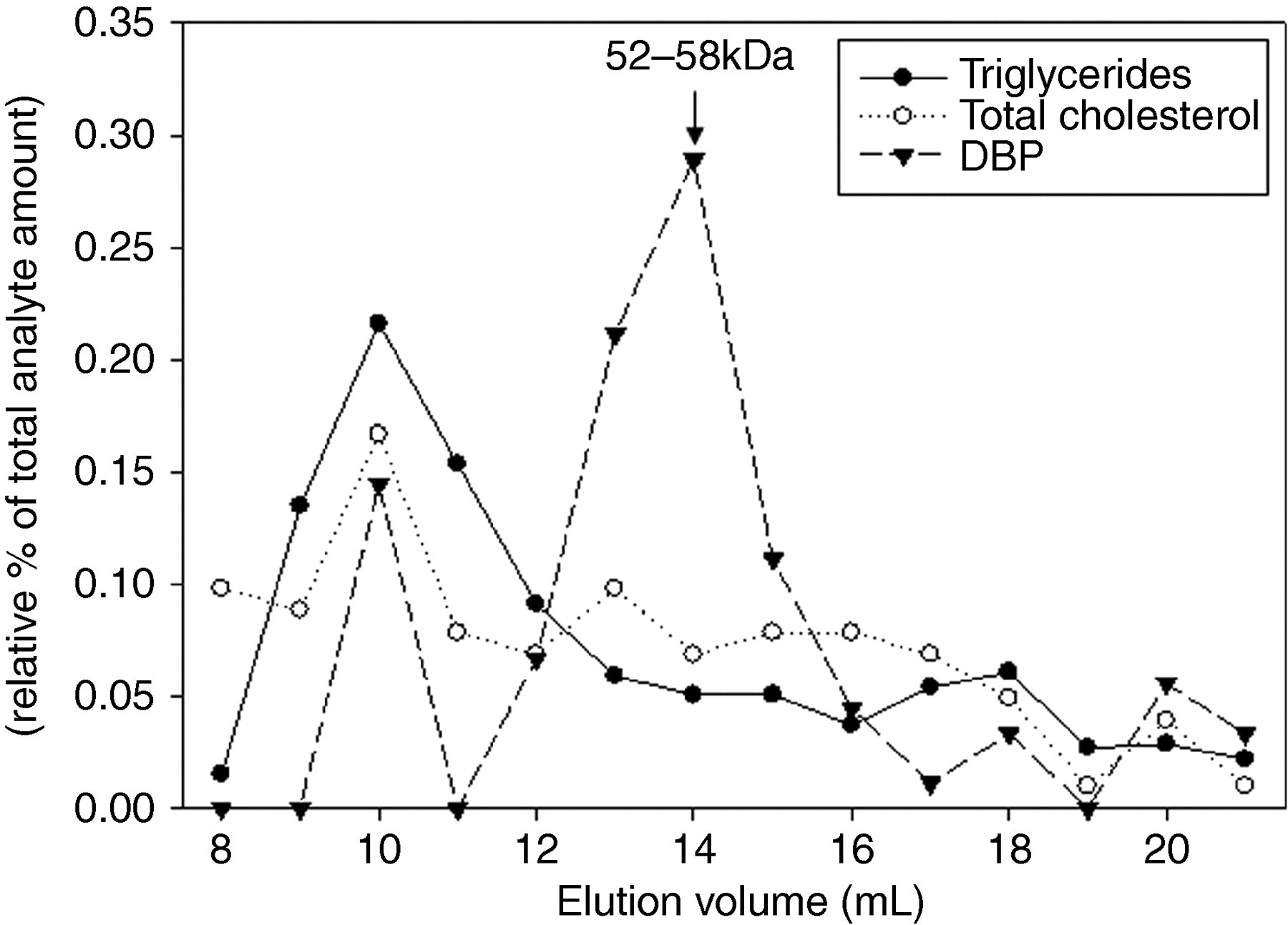
Typical high-pressure gel permeation chromatography of human serum indicating the distribution of vitamin D binding protein (DBP) in the various eluted fractions
Plasma lipid status in relation to DBP
By using multiple regression analysis, triglycerides (Figure 2), total cholesterol, LDL-cholesterol, albumin and 1,25(OH)2-vitamin D3 were found to be predictors of total serum DBP concentration. BMI did not reach significance in this model (Table 1). A negative correlation was observed between serum triglycerides and the 25(OH)-vitamin D3/DBP ratio (r 2 = 0.19; P < 0.0001; Figure 3).
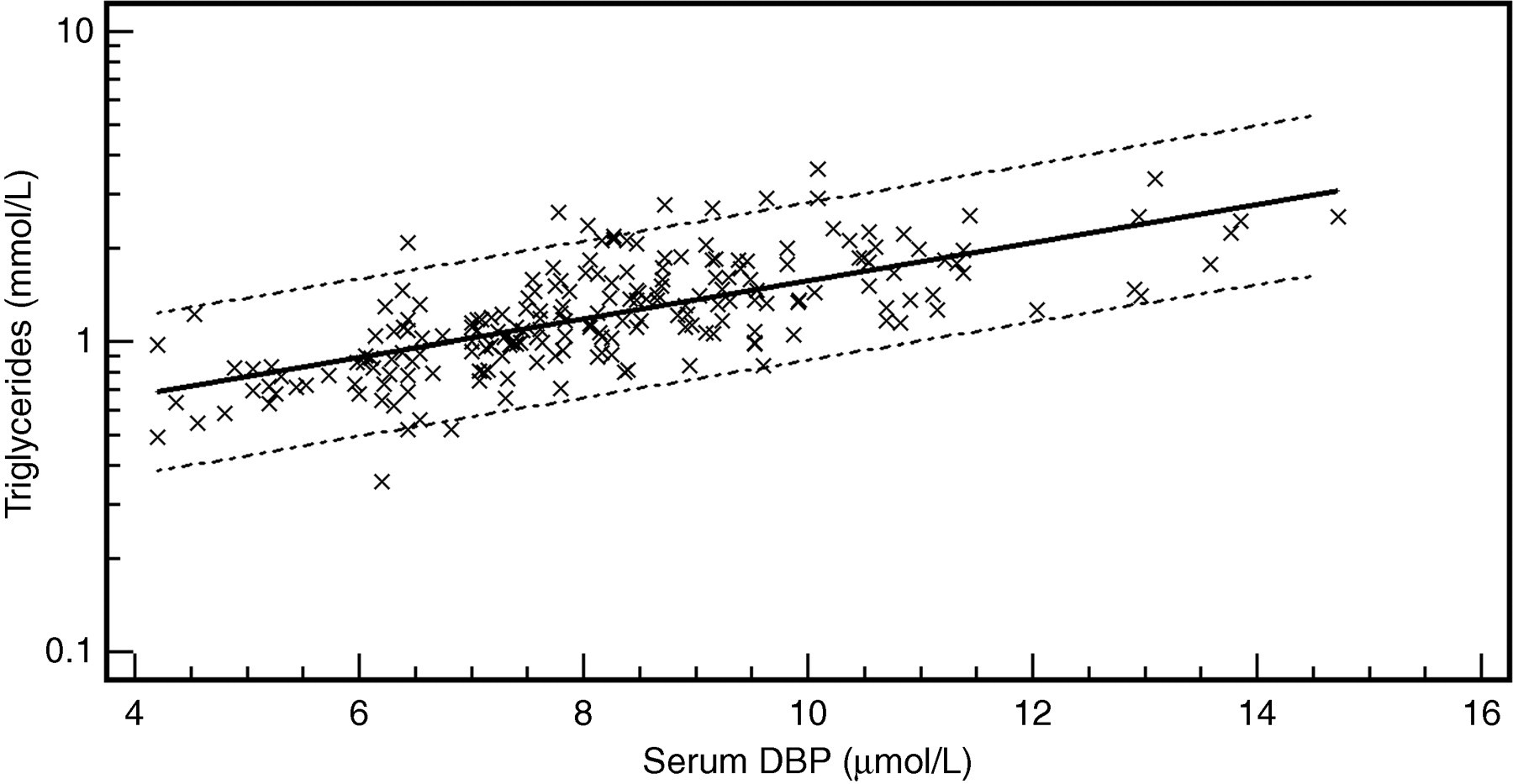
Log-transformed scatter diagram of serum D binding protein (DBP) versus serum triglyceride concentration (Log[y] = −0.4202 + 0.0615x; r 2 = 0.47; P < 0.0001). Dashed lines = 95% prediction interval
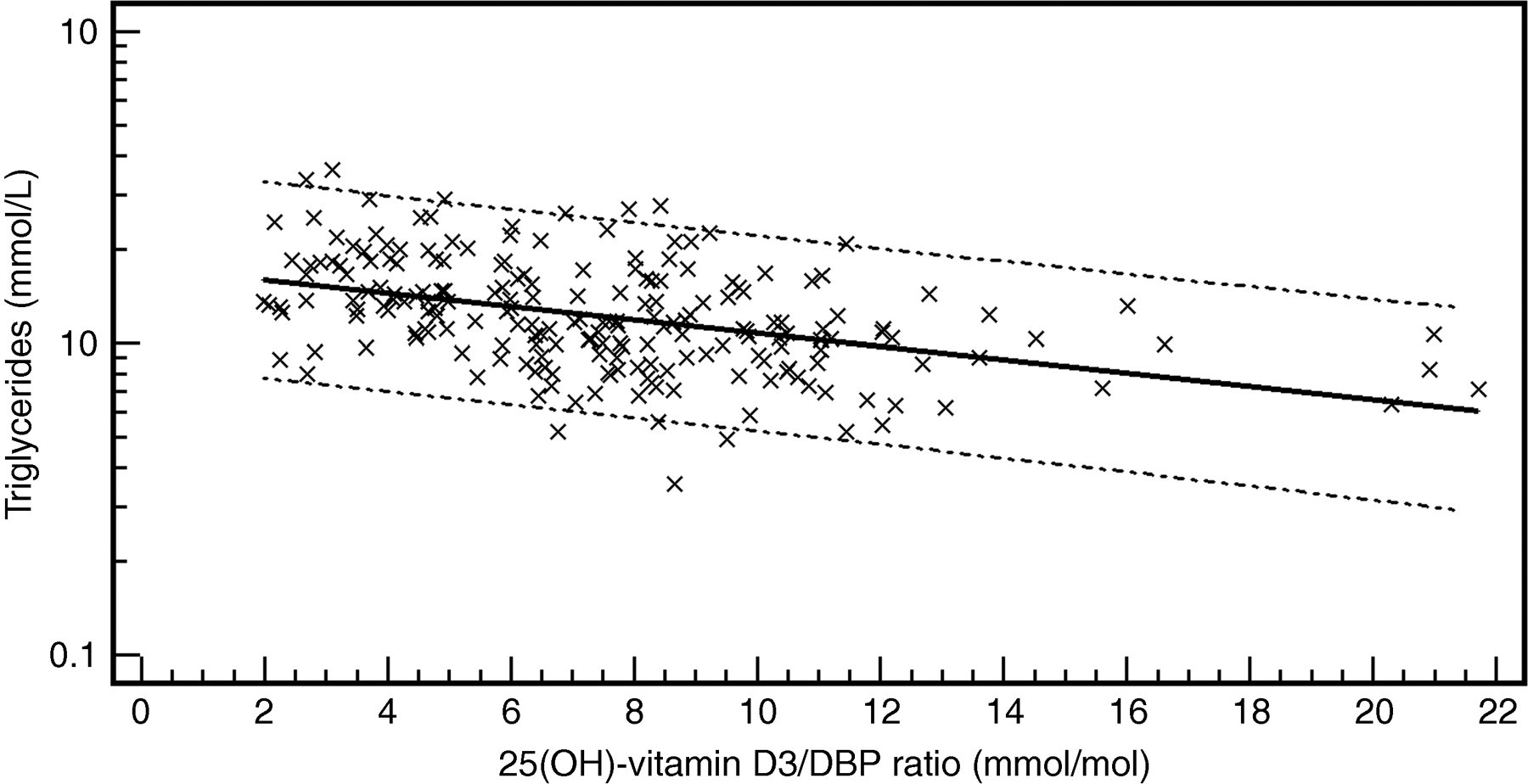
Log-transformed scatter diagram of the serum 25(OH)-vitamin D3/DBP ratio versus serum triglyceride concentration (Log[y] = 0.2444–0.0213x; r 2 = 0.19; P < 0.0001). Dashed lines = 95% prediction interval
Multiple linear regression model of determinants of total serum DBP concentration (μmol/L)*
BMI, body mass index
*r 2 = 0.44 with all variables included
Actin-free and actin-bound DBP
Actin-free DBP concentrations were dependent on the DBP-phenotype, being highest in DBP 1-1 phenotype, intermediate in DBP 2-1 and lowest in DBP 2-2 (Table 2). No differences in actin-free DBP concentrations among (TAAA) n -Alu genotypes were observed. As demonstrated in Figure 4, the actin-bound DBP/DBP ratio correlated significantly with triglycerides (r 2 = 0.34; P < 0.0001). By using multiple regression analysis, besides triglycerides, HDL-cholesterol, triglycerides and total serum DBP were found to be predictors of the actin-bound DBP/DBP ratio (r 2 = 0.51; P < 0.001). Similarly, actin-bound DBP correlated strongly with HDL-cholesterol, LDL-cholesterol, total serum DBP and DBP-phenotype (r 2 = 0.76; P < 0.001).
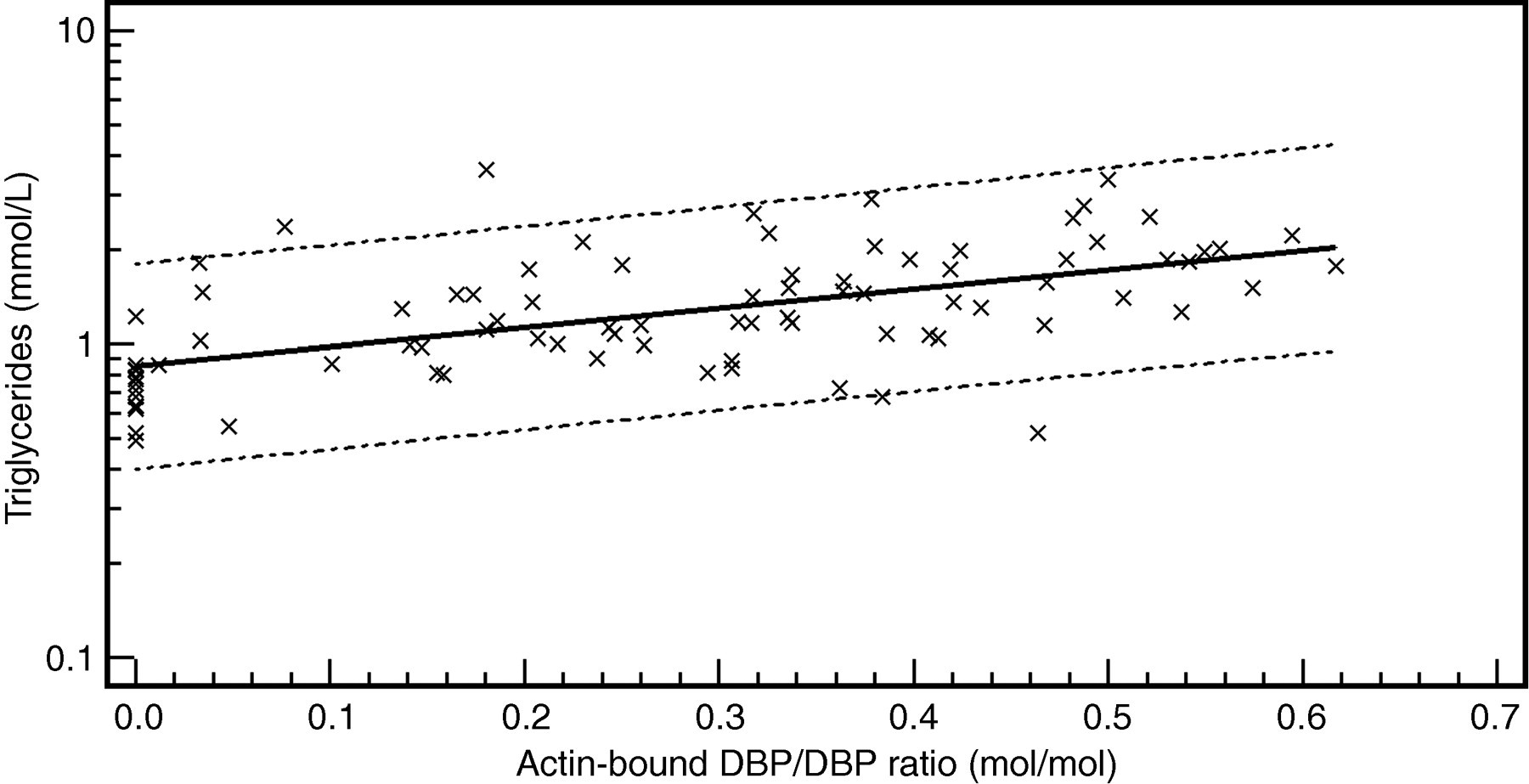
Log-transformed scatter diagram of the serum actin-bound D binding protein (DBP)/DBP ratio versus serum triglyceride concentration (Log[y] = −0.0707 + 0.6139x; r 2 = 0.34; P < 0.0001). Dashed lines = 95% prediction interval
DBP and vitamin D status according to DBP-phenotypes
DBP, vitamin D binding protein
Data expressed as median (interquartile range)
*P < 0.05, **P = 0.009; Kruskal–Wallis analysis according to DBP-phenotypes
Macromolecular-bound DBP fractions
In order to further investigate the functional integrity of DBP, in particular its interactions with actin and lipids, we collected some serum of trauma patients. After PAGE, we performed a Western blot with anti-human DBP polyclonal antibody. As illustrated in Figure 5, two fractions were observed: the free DBP fraction, characterized as the DBP-phenotype and a macromolecular-bound DBP fraction consisting of two electrophoretic bands. To confirm whether the appearance of the electrophoretic bands was related to isolated DBP or to a DBP complex, we incubated the sera with anti-C5a antibodies. An identical Western blot was obtained. Repeating this procedure with anti-actin antibodies led to the disappearance of the formerly lowest macromolecular band, thus suggesting that it actually was a DBP–actin complex.
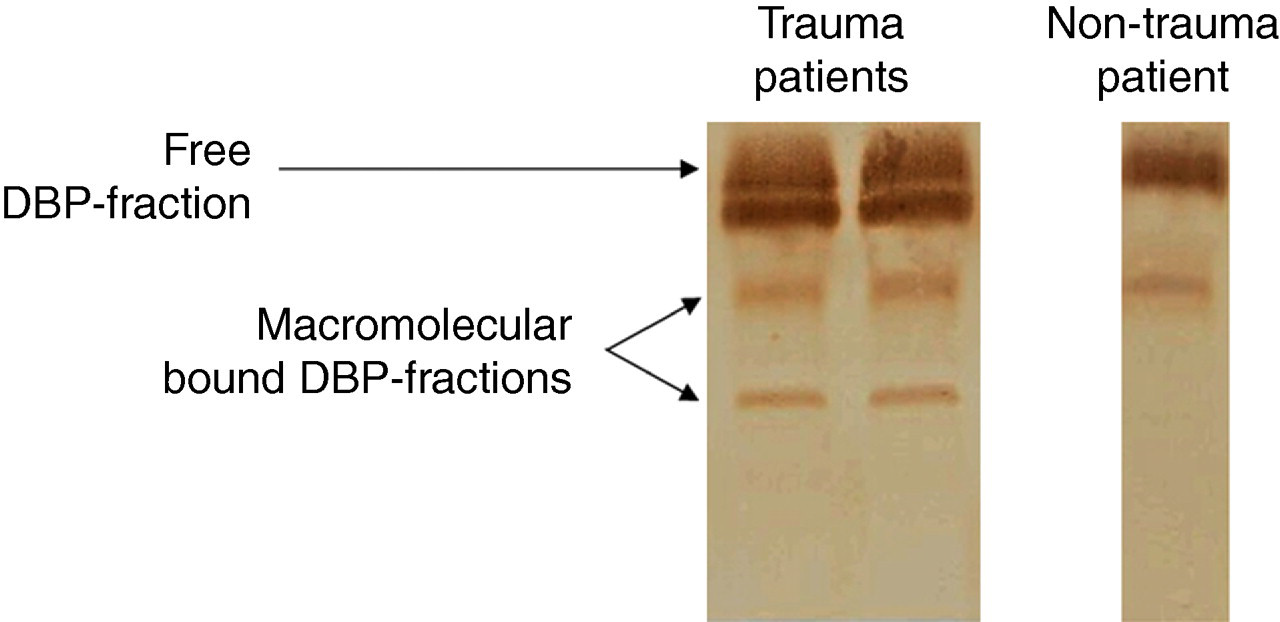
Western blot analysis of vitamin D binding protein (DBP) in trauma patients, identifying the lowest macromolecular band as the DBP–actin complex
Discussion
In the present study, we highlight the association of plasma DBP with lipoproteins. First of all, we confirmed the correlation between total serum DBP concentration and a number of lipid-related parameters (serum total cholesterol, LDL-cholesterol, triglycerides), described earlier in a cystic fibrosis population. 3 The observed relationship between DBP and triglyceride values is supported by animal models. Experimental research with male Sprague–Dawley rats, fed with a high-fat diet revealed syntropic changes in apo A–I, β-actin, DBP prepeptide and haptoglobin. Zhao et al. 22 hypothesized that some physiological changes increased serum DBP, including increased fatty acid caused by a high triglyceride intake and accelerated vitamin D metabolism in order to maintain energy balance under a high-energy diet. In agreement with this study, proteomic analysis showed decreased plasma DBP and reduced liver DBP mRNA concentrations during prolonged fasting. 23
Independent of the precipitation method we used, a partial co-precipitation of DBP, VLDL plus LDL particles was observed. This finding is in line with the results obtained by HPGPC and ultracentrifugation. HPGPC revealed a bimodal distribution of DBP. The macromolecular DBP fraction was characterized by an elution volume very close to the void volume of the HPGPC column and by the presence of lipids (cholesterol, triglycerides). Using ultracentrifugation, we isolated the VLDL fraction and detected DBP. To the best of our knowledge, the influence of the lipid compartment on the functional capacities of DBP has not yet been investigated. Swamy et al. 24 explored the vitamin D and actin binding properties of DBP, without considering the role of triglycerides. The partial association of DBP with VLDL may be supported by a similar complex formation between C-reactive protein and VLDL. 25 Additionally, DBP correlates with serum α-tocopherol (vitamin E), 3 which employs lipoproteins as transport vehicles. Following resorption in the gut, vitamin E is incorporated into chylomicrons and taken up into the liver by the LDL receptor and LDL receptor-related protein. Within the hepatocytes, vitamin E associates with α-tocopherol transfer protein (a-TTP), a cytosolic protein that is required for its intracellular transport and metabolism. 26 In a similar way, lipoproteins play an important role in vitamin D transport because after intestinal absorption, vitamin D is transported on chylomicrons. 27 It is reasonable that in the liver vitamin D associates with DBP and is partly secreted into newly synthesized VLDL particles.
In a third part of this study, we were surprised by the negative correlation between serum triglycerides and the vitamin D3 metabolites/DBP ratio. The pool of plasma triglycerides (±4 g) is a small compartment versus the adipose cells (>10 kg). The size of the adipose tissue compartment is metabolically linked to the plasma triglycerides (the ‘metabolic syndrome’). Apart from the in vitro experiments (e.g. ultracentrifugation) dealing with plasma (in which the adipose tissue does not play a role), there is an in vivo equilibrium with the vitamin D stores in the adipocytes (which are difficult to measure in a clinical setting since a biopsy is required). So, it is clear that the extraction of vitamin D by the adipocytes offers an explanation for the negative correlation coefficient.
The present findings may have clinical consequences in view of the important physiological role of DBP. The important function of DBP in sepsis, shock, trauma and fulminant hepatic necrosis is associated with its role as a strong and important actin scavenging protein in the extracellular and vascular system. During severe cell injury, intracellular actins are released into the systemic circulation. In the presence of Ca2+, gelsolin severs F-actin, it caps the end of F-actin and it nucleates actin-filament assemblies. The generated G-actin is locked in its monomeric state by DBP. Although DBP and gelsolin are both present in large concentrations in blood, their capacity to scavenge actin may still be overwhelmed during massive cell necrosis. 2 – 4 , 28 Low total and actin-free DBP concentrations have been demonstrated to be prognostic markers in situations of severe organ damage. 4 The influence of the different lipid-bound fractions on the actin scavenging function needs to be explored. A larger lipid-bound DBP fraction may account for a more extensive reserve capacity.
Due to its endotoxin binding properties, DBP could be an important scavenger in situations of endotoxinaemia, such as septic conditions. 4 In addition, an endotoxin-stimulated increase in circulating triglyceride-rich particles may have a protective function, as triglyceride-rich lipoproteins (VLDL and chylomicrons) have been shown to bind and inactivate endotoxin. 29 In view of our findings, the cooperative action of DBP and VLDL on endotoxin scavenging should be further investigated. Finally, DBP exerts immunomodulatory functions through its binding to C5a and the generation of C5a co-chemotactic activity. Several in vitro studies point to a positive pathophysiological interaction between the vitamin D carrier and actin scavenger DBP and the complement component C5a. Gressner et al. 30 indicated a minor importance of the predicted interaction between DBP and C5a in severe inflammatory clinical conditions in vivo. However, the role of the lipoprotein-bound DBP fraction has never been investigated in such situations.
The DBP-phenotype distribution in our study is comparable to previous reports. 31 However, some deviation of the observed frequency of (TAAA) n -Alu repeat genotype was observed, compared with published frequencies. The use of capillary electrophoresis with a higher resolution, in comparison with acrylamide gel electrophoresis in combination with silver stain, may explain this difference. 32 The observed dependency of DBP concentration on DBP-phenotype was in accordance to published results. 33 In contrast with 25(OH)-vitamin D3, a marked dependency of 1,25(OH)2-vitamin D3 on the DBP-phenotype was found in our study population. Lauridsen et al. 34 described a similar relationship between 1,25(OH)2-vitamin D concentrations and DBP-phenotypes in perimenopausal women, but also found differences in 25(OH)-vitamin D3 between phenotypes.
Using Western blots, we were able to identify two macromolecular-bound DBP fractions in trauma patients, identifying the lowest macromolecular band as the DBP–actin complex. Further studies are needed to analyse the relevance of the electrophoretic appearance of DBP in various clinical and therapeutic settings.
In summary, we demonstrated in detail the lipid-bound character of DBP. To the best of our knowledge, this study is the first one that discusses the interactions between the different key players (DBP, lipids, actin, vitamin D).
DECLARATIONS