
Abstract
The limited capacity and duration of working memory can impede learning, particularly in online educational settings. As working memory capacity varies among individuals, the impact of cognitive load in online learning may differ between neurotypical and neurodivergent students. This pilot study explored the connection between neurodiversity and cognitive load experienced during online learning in higher education, using electroencephalography (EEG) and eye-tracking to assess neurophysiological responses. The study involved 72 participants, equally divided between neurotypical and neurodivergent students with attention deficit hyperactivity disorder (ADHD), autism spectrum disorder and/or dyslexia. Participants viewed an online video lecture while their eye movements and brain activity were recorded. ADHD traits were assessed with the Adult ADHD Self-Report Scale, autistic traits with the Autism Spectrum Quotient, and dyslexic traits with the Adult Reading History Questionnaire. Participants completed pre-test and post-test questionnaires on the educational content and a computerised cognitive task to evaluate working memory capacity. Results showed that neurodivergent students exhibited higher theta/alpha ratios (EEG), greater extraneous cognitive load (eye fixations on irrelevant features) and reduced intrinsic cognitive load (eye fixations on task-relevant content) compared to neurotypical students. However, no differences in learning performance or working memory capacity were found. These findings suggest that while neurodivergent students may experience distinct patterns of cognitive load during online learning, they can achieve similar learning outcomes, potentially reflecting compensatory strategies. The study serves as proof of concept for employing multi-modal neurophysiological measures of cognitive load to examine its effects on online learning in neurodiverse classrooms and suggests the need for larger samples to clarify the relationship between neurodiversity and cognitive load, with the ultimate aim of informing inclusive online education design.
This pilot study explored how cognitive load (or mental effort) during online learning differs between neurotypical students and those with attention deficit hyperactivity disorder (ADHD), autism spectrum disorder (ASD) and/or dyslexia. Participants watched an online lecture while their eye movements and brain activity were recorded. Although marginal neurocognitive differences were observed, learning performance was similar across groups. The findings contribute to understanding the impact of neurodiversity on learning and can inform inclusive education practices.
Introduction
In recent years, online education has become more prominent, with an increasing number of students completing part or all their classes in an online learning environment (Alevizou, 2015). Coinciding with this trend is a rise in the number of neurodivergent (ND) students engaging in online learning, such as those with attention deficit hyperactivity disorder (ADHD), autism spectrum disorder (ASD) and dyslexia (Dixon et al., 2023). The term ‘neurodivergent’ refers to individuals whose brain functions fall outside prevalent societal norms, while ‘neurotypical’ describes individuals with typical brain functioning and development (Goldberg, 2023; Shah et al., 2022). Many factors likely impact the experience of ND students in online learning, but they have not yet been extensively studied (Adam et al., 2017; Cheawjindakarn et al., 2013; Tinmaz et al., 2022). One such factor is cognitive load, also known as working memory load. Cognitive load refers to the amount of working-memory resources required to complete a task, and it is thought that learning is impaired when task demands exceed capacity (Paas et al., 2016; Sweller, 1994). Cognitive Load Theory, which aims to optimise the amount of used working memory resources to best facilitate learning, is well-established for neurotypical (NT) students (Mutlu-Bayraktar et al., 2019; Skulmowski & Xu, 2022). For instance, it has been found that cognitive load can impact a student's willingness to engage with learning tasks and be a significant predictor of acceptance of online learning (Feldon et al., 2019; Tzafilkou et al., 2021).
Historically, cognitive load has often been assessed through indirect subjective measures such as questionnaires and rating scales (Ayres, 2017; Sweller, 1994; Sweller et al., 2011). Some scales distinguish between intrinsic cognitive load (ICL) pertaining to educationally relevant stimuli, extrinsic cognitive load (ECL) pertaining to irrelevant stimuli that distract from learning, and sometimes germane cognitive load which pertains to supportive stimuli that enhance learning (Klepsch et al., 2017; Leppink et al., 2013), although the nature of the latter has been disputed (Kalyuga, 2011). However, these self-report measures have been brought into question regarding their potential intrusiveness during task performance and susceptibility to response biases (Brünken et al., 2003; Naismith et al., 2015; Orru & Longo, 2018). As such, some researchers have turned to neurophysiological measures in an attempt to objectively assess cognitive load, including electroencephalography (EEG), functional magnetic resonance imaging, and eye-tracking (Antonenko et al., 2010; Whelan, 2007). For instance, eye-tracking and in particular area of interest (AOI) analysis allows researchers to distinguish between attention directed towards educationally relevant content (contributing to ICL) versus task-irrelevant interface elements (contributing to ECL), operationalising Cognitive Load Theory's core distinction between productive and counterproductive mental effort (de Jong, 2010; Liu & Cui, 2025; Sáiz-Manzanares et al., 2024; Zagermann et al., 2016).
Prior research suggests that ND students may experience higher cognitive load during learning than their NT peers. In ADHD, difficulties in sustaining attention and reduced working-memory capacity have been consistently documented (Kofler et al., 2020; Ortega et al., 2020). Dyslexia has been linked to phonological processing difficulties that increase the effort required to integrate spoken and written information in multi-media learning contexts (Peng & Fuchs, 2016; Wang et al., 2022). In autism, atypical modulation of theta and alpha oscillations during working-memory tasks indicates differences in how cognitive resources are allocated under load (Larrain-Valenzuela et al., 2017). Recent work further shows that ND university students report higher levels of ECL in online environments compared with NT peers (Le Cunff et al., 2024b). These findings suggest that ND students may experience increased cognitive load during online learning.
Although ADHD, ASD and dyslexia have been found to be associated with differences in working memory (Kofler et al., 2020; Rabiee et al., 2020; Wang et al., 2022), few studies have examined cognitive load among ND online students and most exclude co-occurring conditions (Le Cunff et al., 2024a). This pilot study aimed to assess the feasibility of combining EEG, eye-tracking and validated self-report trait scales to investigate cognitive load in a neurodiverse university sample and to examine potential differences in cognitive load in online learning between NT and ND students in higher education. Co-occurrence rates as high as 25–40% have been reported between ADHD and dyslexia (Sexton et al., 2012; Willcutt & Pennington, 2000) and it is estimated that 28–44% of adults diagnosed with ASD also meet the criteria for ADHD, with some estimates as high as 50–70% (Hours et al., 2022; Polderman et al., 2014). Such high co-occurrence rates make disentangling the unique neural correlates of each condition challenging (Astle et al., 2022). However, these conditions are often investigated separately, with co-occurring conditions excluded from the samples. In this study, participants reporting a diagnosis of ADHD, ASD or dyslexia were instead grouped together, and their results compared to a NT group. In addition, neurodiversity was further operationalised dimensionally using self-reported trait scores on validated scales for the three conditions (Baron-Cohen et al., 2001; Kessler et al., 2005; Lefly & Pennington, 2000), thus ensuring the inclusion of co-occurring neurodevelopmental conditions. Based on the emerging evidence, it was hypothesised that cognitive load in online learning would be higher in the ND group and that higher trait scores on ADHD, ASD and dyslexia scales would be associated with higher cognitive load.
Methods
This study was approved by the Research Ethics Committee at King's College London (Ref: LRS/DP-22/23-35603). The protocol was co-designed with a Research Advisory Board of ND students who reviewed the experimental procedure and documents to ensure the comfort of participants and minimise the impact of factors that might affect responses in neuroscience experiments (Le Cunff, Glover et al., 2024).
Participants
All participants were required to be currently enrolled full-time in a UK campus-based university degree programme that requires the use of an online learning platform. They had to be over 18 years old and a native English speaker.
In addition, ND participants had to self-report a diagnosis of at least one of the following conditions: ADHD, ASD or dyslexia, and to specify whether this diagnosis had been made by a healthcare professional, an educational psychologist or was self-identified (see Sample Characterisation). In line with our co-design process with the Research Advisory Board, we included participants across all three of these categories. This decision was made to reflect the reality of university cohorts in which many ND students remain undiagnosed or are mid-assessment. In the United Kingdom, adult diagnostic services face multiyear backlogs, and late diagnosis is especially common among women, who are frequently missed or misdiagnosed due to camouflaging behaviours (Lang, 2024; Quinn & Madhoo, 2014; Smith et al., 2024). Including self-identified students therefore helped ensure that the sample was representative of the lived experiences of ND university populations. Lastly, although ND participants in this study represent a range of diagnoses, we follow recent work suggesting that including overlapping traits reflects the lived reality of neurodivergence in educational contexts (Luo et al., 2019).
As the cross-sectional design prioritised ecological validity and minimal burden for ND participants by limiting data collection to variables directly relevant to the research aims, medication status was not assessed in this study to avoid unnecessary disclosure of sensitive clinical information. In addition, given the pilot sample size and expected heterogeneity in medication type, dosage, timing and adherence among participants, meaningful subgroup analyses would not have been statistically powered.
Before participating, all individuals were provided with an information sheet and written informed consent was obtained from each participant.
Procedure
The study used EEG, eye-tracking technology, computerised cognitive tasks and online learning tasks. EEG theta/alpha ratio (TAR) and eye blink rate (EBR) were used to measure cognitive load. These metrics are frequently employed in cognitive load research as they provide direct estimates of load (Antonenko et al., 2010; Siegle et al., 2008). A higher TAR indicates increased cognitive load (Gevins & Smith, 2003). Furthermore, higher EBR is an oculomotor measure of higher cognitive load that is not affected by luminosity and gaze position (Zagermann et al., 2016). Considering both brain-based and eye-related measures provides converging evidence from central and peripheral nervous systems (İşbilir et al., 2019; Scharinger et al., 2020). We also measured midfrontal theta and occipital alpha as the spectral power of those two bands in these channels is frequently extracted to study brain activity related to working memory, with higher frontal theta and lower occipital alpha both indicating increased cognitive processing (Popov et al., 2018).
The experiment was conducted in a quiet, dimly lit room. Participants were seated in front of a computer screen with dimensions of 1920 × 1080 pixels. They first completed a short multiple-choice knowledge pre-test on the study topic. They were then presented with a 10-minute instructional video embedded within an interface mimicking a real online learning environment (Supplemental material A). The video covered meteorology basics adapted from prior cognitive load research (Liu et al., 2011). Based on a systematic review of cognitive load in online learning (Le Cunff et al., 2024a), the interface incorporated redundant modalities, meaning that core instructional content was delivered via both narration and synchronised onscreen text and images, consistent with multimedia learning principles (Mayer, 2014). This redundancy was designed to support learning while controlling for modality-specific processing differences (Mayer, 2014; Paivio, 1986). Linguistic complexity was standardised at university level with a Flesch Reading Ease Score (Flesch, 1948) of 49.9 to ensure appropriate difficulty across participants. The design deliberately limited interactivity in order to isolate perceptual and attentional components of cognitive load associated with online learning rather than executive demands such as navigation, planning or multi-tasking (Costley et al., 2021; Sharma et al., 2021). While this visually realistic but non-clickable online interface might improve internal validity by reducing variability introduced by task-switching behaviours (Liefooghe et al., 2008), it likely reduces the ecological validity of the online learning environment (see Discussion section).
For eye-tracking analysis, AOIs corresponding to ICL and ECL were pre-defined through consensus by three researchers. The video player was designated as the sole ICL region as it contained the instructional content directly relevant to the learning task. In contrast, peripheral elements such as the header and side banners were classified as ECL regions as they contained no instructional content and could act as distractors. This classification was based on the premise that visual fixations indicate where cognitive processing resources are allocated and, therefore, a higher proportion of fixations on the video reflects engagement with task-relevant material whereas fixations on peripheral elements reflect processing of task-irrelevant stimuli (Liu, 2014; Liu & Cui, 2025; Mitra et al., 2017; Sáiz-Manzanares et al., 2024; Zagermann et al., 2016; Zu et al., 2020). Eye movements were recorded using an Eyelink 1000 Plus tracker (SR Research Ltd., Canada) with a camera-to-screen distance of 535 mm. The eye-tracker was mounted below the screen and a chin rest limited head movement. The eye-tracker was calibrated at the start of the experiment using a 9-point grid, and validation confirmed calibration accuracy within 0.5° average and 1.0° maximum error. Gaze data was acquired from both eyes at 500 Hz. Stimulus display and responses were synchronised via the Experiment Builder software (SR Research).
EEG signals were collected from a 32-channel acquisition system with silver/silver chloride (Ag/AgCl) electrodes (Emotiv, USA) fitted on a 56 cm cap (EasyCap, Herrsching, Germany) at locations AFz (DRL), FCz (CMS), Fp1, Fp2, F7, F3, Fz, F4, F8, FT9, FC5, FC1, FC2, FC6, FT10, T7, C3, Cz, C4, T8, CP5, CP1, CP2, CP6, TP9, TP10, P7, P3, Pz, P4, P8, O1, Oz, O2 as defined by the international 10–20 system (Klem, 1999). A 0.5% saline solution (2–3 mL) was applied to each electrode and the programme TestBench SDK (Emotiv, USA) was used to verify the impedance of each electrode. EEG data was amplified and digitised continuously (Emotiv Extender, SCAN 4.3) with a digitisation at 1,024 Hz and down-sampling to 128 Hz. We did not record a resting-state baseline because our focus for this pilot study was on group differences during online learning. Standard rest baselines alter the sensory context (eyes-closed rest markedly increases alpha power and eyes-open rest removes the lecture's dynamic audiovisual stimulation), and such mismatches make baseline-task contrasts hard to attribute specifically to cognitive load (Keil et al., 2014; Klimesch, 1999; Laufs et al., 2003). To reduce the impact of between-person baseline variability without a rest block, we used relative power to reduce sensitivity to differences in overall signal amplitude (Gevins & Smith, 2003; Klimesch, 1999). Adding rest blocks would have also lengthened the multimodal session, increasing risk of calibration-drift and participant fatigue (Gevins & Smith, 2003; Holmqvist et al., 2011; Keil et al., 2014). Consistent with our co-designed protocol to minimise burden for ND participants, we therefore prioritised continuous task recordings (Le Cunff, Glover et al., 2024).
After watching the video, the well-established and validated computerised OSPAN task (Kane et al., 2001) was used to measure participants’ working memory capacity (WMC). The task involves participants solving a series of basic mathematical problems while simultaneously attempting to remember a series of unrelated words. It was hosted on the experimental platform Gorilla (Anwyl-Irvine et al., 2020) using the implementation of Janssen et al. (2022). Participants then filled out three validated scales to assess ADHD, ASD and dyslexia traits, respectively: the Adult ADHD Self-Report Scale (ASRS; Kessler et al., 2005), Autism Spectrum Quotient (AQ; Baron-Cohen et al., 2001), and Adult Reading History Questionnaire (ARHQ; Lefly & Pennington, 2000). Lastly, they completed a multiple-choice knowledge post-test on the study material. All scales, pre-test and post-test questionnaires were hosted on Qualtrics (Qualtrics, 2024). The full session lasted approximately 1 hour.
Data Processing and Analysis
Eye-tracking data were processed and exported with the EyeLink Data Viewer software package (SR Research, Version 3.2.1., 2018) and segmented into 10 1-minute trials spanning the entire duration of the video. Blinks were identified as zero pupil diameter for over 100 ms (Hollander & Huette, 2022). Ocular cognitive load was operationalised as the mean EBR in blinks/minute. Fixations were based on the strict average across both eyes and adjacent fixations were merged if they were less than 100 ms apart and less than 0.5 degrees apart (Mahanama et al., 2022). Gaze-based ICL and ECL were calculated as ratios of fixations in relevant versus irrelevant AOIs to the total number of fixations (Lai et al., 2013; Lavie & Dalton, 2014; Liu et al., 2022).
The MATLAB toolbox EEGLAB was used to process and export the EEG data (Delorme & Makeig, 2004). Bandpass filtering was applied with a high-pass cutoff of 0.1 Hz and a low-pass cutoff of 30 Hz, and the data was re-referenced to the average reference. Artefact-contaminated signals in the continuous data were detected and rejected using an automated EEGLAB plugin (clean_rawdata) to identify artefact EEG channels if they were flat for more than 5 s, poorly correlated with surrounding channels (r < 0.8), and/or too noisy (±5 SD). The EEG data was segmented into 60s epochs to facilitate the detection and rejection of temporal artefacts and to ensure temporal alignment across EEG and eye-tracking modalities (Delorme & Makeig, 2004; Holmqvist et al., 2011). This segmentation approach also allows for more reliable artefact rejection and preprocessing prior to averaging, a standard step in EEG cleaning pipelines (Coelli et al., 2024; Keil et al., 2014). EEG signals were decomposed into independent source components using independent component analysis (Lee et al., 1999) to isolate stereotypic artefacts from non-brain source activations present in the epoched EEG data. The ICLabel plugin (Pion-Tonachini et al., 2019) was used to distinguish between brain-related independent components (ICs) and artefact ICs. The identified artefact ICs were removed from the EEG signals. ICs classified with more than 90% probability as artefactual (e.g., eye movement and muscle activity) were flagged and removed from the EEG signal. This method allows for more precise separation of brain activity from noise, improving the accuracy of the final EEG analysis (Delorme & Makeig, 2004; Luca Pion-Tonachini et al., 2019). The relative spectral power of each band, defined as the ratio of the power in a specific frequency band to the total power across all measured frequency bands, was calculated using a fast Fourier transform and the relative power spectral density (PSD) in the theta (4–7 Hz) and alpha (8–12 Hz) bands was computed using the Welch's method (implemented in the pwelch MATLAB function). This approach helps to normalise the data by accounting for individual differences in baseline brain activity (Klimesch, 1999). The spectral power of those two bands was also averaged for channels relevant to working memory related brain activity to obtain measures of midfrontal theta (average of F3, Fz, F4) and occipital alpha (average of O1, Oz, O2) power.
All files from eye-tracking, EEG recordings, cognitive task and questionnaires were compiled using Python 3.8 (Van Rossum & Drake, 2009), and the analyses were performed with the statistical software Jamovi (The Jamovi Project, 2023). WMC was computed using the OSPAN partial-scoring method (Conway et al., 2005), learning performance (LP) was operationalised as post-test minus pre-test, ocular-motor-based cognitive load as the EBR, and EEG-based cognitive load as the relative PSD in the theta and alpha bands to obtain the TAR. Descriptive statistics were generated for all variables, including age, gender, study level, neurodiversity scores for each scale (ASRS, Autism Spectrum Quotient [ASQ] and ARHQ), WMC, LP, ICL, ECL, CL, midfrontal theta and occipital alpha power across both NT and ND groups. Skewness, kurtosis and visual examination of the histogram and the QQ plots for both NT and ND students did not show evidence of non-normality, allowing for the use of parametric tests with ASRS, ASQ, ARHQ, WMC, LP, EBR, ICL, ECL, TAR, midfrontal theta and occipital alpha as variables of interest. A Pearson correlation matrix was created to examine the relationship between these variables.
No correction for multiple comparisons was applied in the initial analysis because the comparisons were specified a priori as part of the original study design. In pilot and exploratory research, strict correction procedures can sometimes suppress meaningful patterns by overemphasising type I error at the expense of detecting genuine but subtle effects (Benjamini, 2010; Rothman, 1990). Our approach prioritises the identification of potentially important associations that may guide future hypothesis-driven studies. Nonetheless, we recognise the importance of gauging the robustness of findings when multiple statistical tests are conducted. To this end, we applied the Benjamini-Hochberg False Discovery Rate (FDR) correction (Benjamini & Hochberg, 1995) as a secondary, conservative step. This approach allows us to balance the risk of false positives with the exploratory intent of the study, particularly in a field where underpowered designs and heterogeneity in ND populations are common. Reporting both unadjusted and adjusted results enables a more nuanced interpretation of the data while maintaining transparency and scientific rigour.
Results
Sample Characterisation
The sample comprised 72 participants, including 36 ND (24 women, 8 men, and 4 non-binary) and 36 NT (26 women, 9 men, and 1 non-binary) students. Forty participants were studying at the bachelor's level (55.6%), 26 participants at the master's level (36.1%) and 6 participants at the doctoral level (8.3%). Fifteen ND participants declared a diagnosis of ADHD (41.7% of ND), three declared a diagnosis of ASD (13.9%), six declared a diagnosis of dyslexia (16.7%), three declared a combined diagnosis of ADHD and ASD (8.3%), and six declared a combined diagnosis of ADHD and dyslexia (16.7%) (Supplemental material B). Twenty-two ND participants were diagnosed by a healthcare professional (61.1%), 10 were diagnosed by an educational psychologist (27.8%), and 4 were self-diagnosed (11.1%) (Supplemental material C). There was no significant difference in age, gender or study level between the two groups (Table 1).
Demographics.
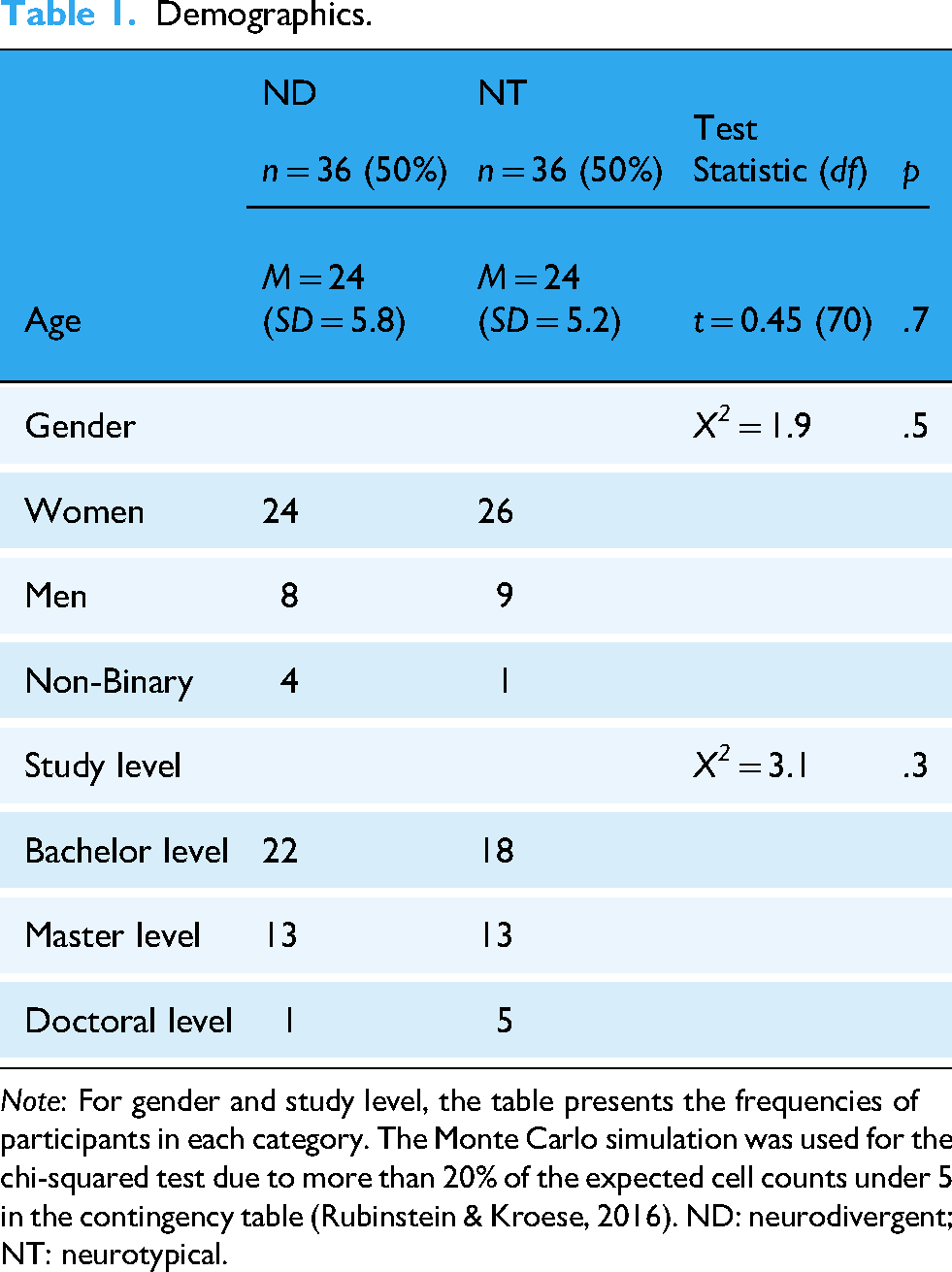
Note: For gender and study level, the table presents the frequencies of participants in each category. The Monte Carlo simulation was used for the chi-squared test due to more than 20% of the expected cell counts under 5 in the contingency table (Rubinstein & Kroese, 2016). ND: neurodivergent; NT: neurotypical.
Descriptive Statistics
ND students exhibited significantly higher scores for ADHD traits as measured by the ASRS, t(70) = 8.06, p < .001, d = 1.90, autistic traits as measured by the ASQ, t(70) = 4.78, p < .001, d = 1.13, and dyslexic traits as measured by the ARHQ, t(70) = 5.66, p < .001, d = 1.33. TAR was significantly greater for ND students (M = 1.99, SD = 0.30) compared to NT students (M = 1.81, SD = 0.37), t(70) = 2.22, p < .05, d = 0.52. Additionally, ND students demonstrated a lower ICL, t(70) = −2.20, p < .05, d = −0.52, and a higher ECL, t(70) = 2.13, p < .05, d = 0.50. In ND students midfrontal theta was found to be higher, t(70) = 2.12, p < .05, d = −0.50 and occipital alpha to be lower, t(70) = −2.06, p < .05, d = −0.49. There were no differences in WMC, EBR, and LP between ND and NT students (Table 2).
T-Tests Examining Neurodiversity and Cognitive Load in Online Learning.

Note: * p < .05, ** p < .01, *** p < .001. ARHQ: Adult Reading History Questionnaire; ASQ: Autism Spectrum Quotient; ASRS: Adult ADHD Self-Report Scale; EBR: eye blink rate; ECL: extraneous cognitive load; ICL: intrinsic cognitive load; LP: learning performance; MidTheta: midfrontal theta power; ND: neurodivergent; NT: neurotypical; OccAlpha: occipital alpha power; TAR: theta/alpha ratio; WMC: working memory capacity.
As explained in the Methods section, our initial analysis did not adjust for multiple comparisons. Nonetheless, as a robustness check, we also applied the Benjamini-Hochberg FDR correction (Supplemental material D). After this adjustment, three group-level differences – ADHD, autism and dyslexia trait scores – remained statistically significant. Meanwhile, differences in cognitive load measures such as ICL, ECL, TAR, midfrontal theta and occipital alpha did not retain significance but remained near threshold (adjusted p-values just above .05) with moderate-to-large effect sizes (Cohen's d ≈ 0.5).
Relationship Between Neurodiversity and Cognitive Load
There was a medium correlation between TAR and ADHD traits as measured by the ASRS (r = 0.52, p < .05) and a small correlation between TAR and dyslexic traits as measured by the ARHQ (r = 0.25, p < .05). ADHD traits were positively correlated with midfrontal theta power (r = 0.41, p < .05) and negatively correlated with occipital alpha power (r = −0.30, p < .05). ASD traits did not correlate with any other measures. Dyslexic traits were positively correlated with midfrontal theta power (r = 0.22, p < .05). ICL, ECL, WMC, EBR and LP did not significantly correlate with any of the individual scales of neurodevelopmental conditions. Furthermore, ICL and ECL showed a significant negative correlation (r = −0.90, p < .001), in line with their operationalisation in this study, where ICL pertains to relevant on-screen information and ECL to irrelevant on-screen information. The resulting correlation matrix (Table 3) suggests a complex interplay between cognitive load and neurodiversity.
Pearson Correlation Coefficients for Study Variables.
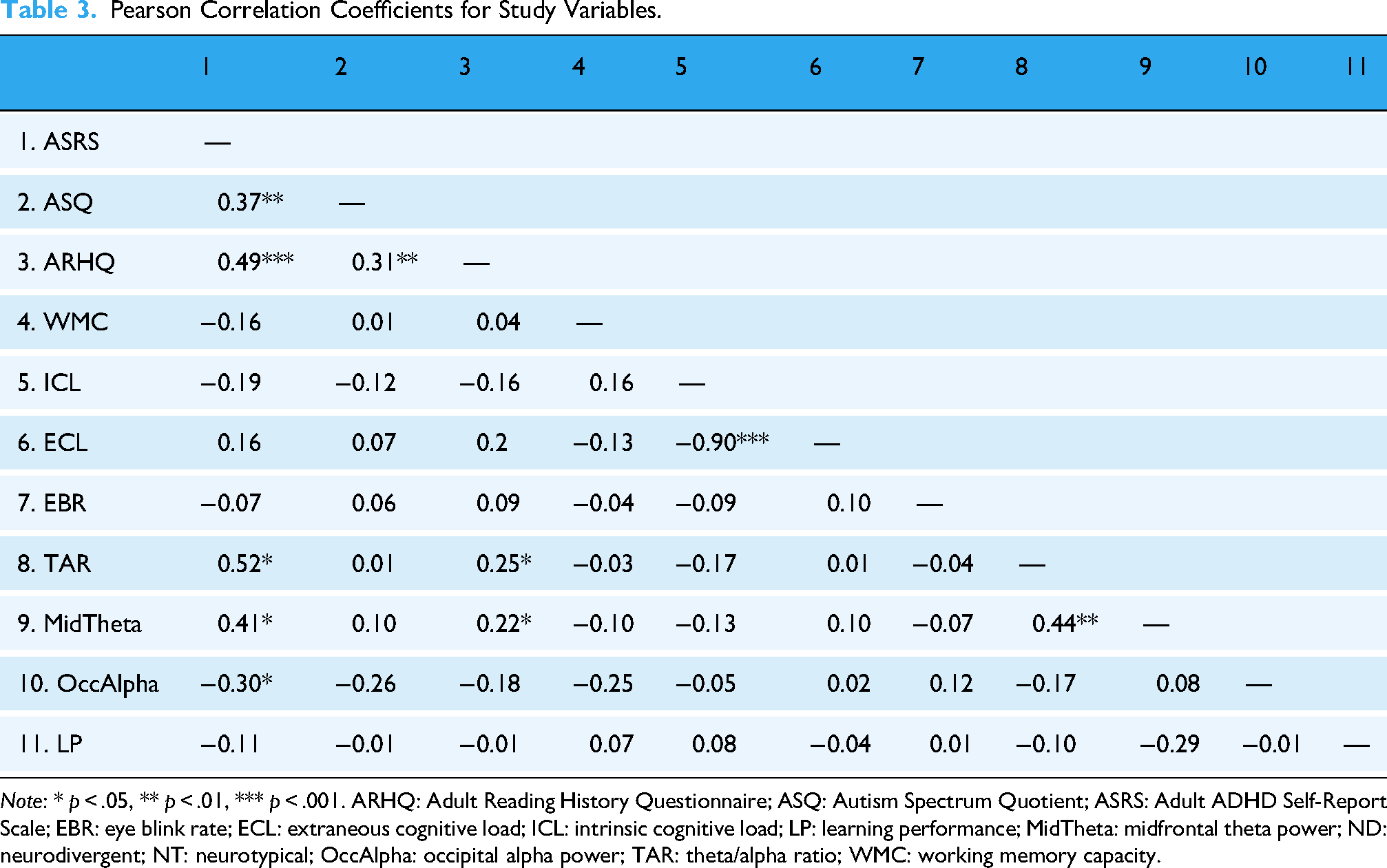
Note: * p < .05, ** p < .01, *** p < .001. ARHQ: Adult Reading History Questionnaire; ASQ: Autism Spectrum Quotient; ASRS: Adult ADHD Self-Report Scale; EBR: eye blink rate; ECL: extraneous cognitive load; ICL: intrinsic cognitive load; LP: learning performance; MidTheta: midfrontal theta power; ND: neurodivergent; NT: neurotypical; OccAlpha: occipital alpha power; TAR: theta/alpha ratio; WMC: working memory capacity.
While multiple tests were conducted – raising the risk of false positives – we approached this analytically by reporting both uncorrected and FDR-adjusted p-values. After FDR correction, three group-level differences (ASRS, ASQ and ARHQ scores) remained statistically significant, suggesting that these trait differences are robust. Other cognitive load measures – including ICL, ECL, TAR, midfrontal theta and occipital alpha – did not retain statistical significance after correction.
In the context of an exploratory pilot study with moderate-to-large effect sizes, these trends might represent meaningful directions for future investigation, especially given the consistency of the results across independent neurophysiological indicators, which will be discussed in the following section alongside directions for future research.
Discussion
This pilot study explored cognitive load in online learning using a novel combination of EEG, eye-tracking and validated trait scales. As a proof-of-concept, it aimed to assess the feasibility of integrating neurophysiological and behavioural measures in educational neuroscience, particularly for populations often excluded due to heterogeneity or diagnostic overlap.
A key finding was that the TAR, an EEG measure of cognitive load, was significantly higher in ND compared to NT students, although after FDR correction this was only a trend towards significance (p = .058). The marginally elevated TAR observed in ND students is broadly consistent with previous studies that have reported altered neural oscillatory activity associated with cognitive load and theories of atypical cognitive processing in ND populations (Le Cunff et al., 2023). However, the exact nature of these neural oscillations and relationship to cognitive load in ND students warrant further investigation.
ND students also exhibited marginally reduced ICL and higher ECL, as evidenced by fewer gaze fixations on the task-relevant video and elevated fixations on irrelevant design elements compared to NT students. A substantial body of research has established a link between working memory and sustained attention, both functionally (Morey et al., 2011; Unsworth & Spillers, 2010) and anatomically (Silver & Feldman, 2005). Our findings, although non-significant after adjustment for multiple comparisons, are in line with prior research suggesting attentional difficulties in ND populations (Lewandowska et al., 2014; Schachar et al., 2023). The marginal differences in ICL and ECL may be indicative of the attentional and perceptual processing differences frequently documented in ADHD, ASD and dyslexia, which may compromise the ability to focus on task-relevant information (Alloway et al., 2014; Johnson et al., 2007; Lewandowska et al., 2014). Furthermore, the increased distractibility, implied by fewer task-relevant fixations, aligns with accounts of greater susceptibility to distractions in conditions such as ADHD (Osborne et al., 2023).
It is worth noting that ADHD, ASD and dyslexia traits were all positively correlated together in our sample, in line with previous research showing high rates of co-occurrence between these conditions (Russell & Pavelka, 2013), however neither ICL nor ECL significantly correlated with any of the individual scales measuring ND traits. The presence of multiple co-occurring conditions within this group may have had a cumulative effect on cognitive load, resulting in significant differences when compared as a whole to the NT group, but the unique contribution of each condition to cognitive load may be less pronounced when examining the trait scales individually in the correlation analysis. In addition, the correlation analysis assumes a linear relationship between the variables. However, the relationship between ND traits and cognitive load may follow a more complex pattern. For example, there might be a threshold effect, where cognitive differences only become apparent above a certain level of trait severity (Goldberg, 2023). Finally, the relationship between neurodiversity and cognitive load may be influenced by interactions among the different traits or with other factors not captured in the study. These interactions could obscure the direct correlations between individual trait scales and cognitive load measures. Rather than treating this heterogeneity as a confound, we view it as a reflection of the complexity of real-world neurodivergence, where diagnostic overlap is common and samples representing single diagnoses are rare (Luo et al., 2019). Designing for such variability is essential to developing inclusive learning strategies.
Positive correlations were observed between ADHD traits and both the TAR and midfrontal theta power. This is consistent with evidence suggesting that individuals with pronounced ADHD traits may experience elevated cognitive load in certain contexts (Lewis & Brown, 2012; Mukherjee et al., 2021). Furthermore, a medium negative correlation was also identified between ADHD traits and occipital alpha. As the literature on occipital alpha power in ADHD presents varied results (Le Cunff et al., 2023), further research is needed to investigate the exact parameters underlying this relationship. Dyslexia traits also positively correlated with both TAR and midfrontal theta power. As the video involved both reading and auditory comprehension of educational content, this finding aligns with theories proposing that dyslexia is characterised by deficits in phonological processing, potentially leading to increased cognitive load during learning tasks (Kizilaslan & Tunagür, 2021; Knoop-van Campen et al., 2018). ASD traits did not show significant correlations with any neurophysiological measures of cognitive load. This is consistent with prior research indicating that while NT individuals exhibit modulation of theta and alpha power according to cognitive load, such modulation might not be prevalent in ASD (Larrain-Valenzuela et al., 2017).
The EBR, another indicator of cognitive load, did not show differences between groups and showed no correlations with neurodiversity trait scores for ADHD, ASD or dyslexia. This result was unexpected, as a reduced EBR is often associated with increased cognitive load (Siegle et al., 2008). Specifically, cognitive load and EBR have been found to be inversely correlated in tasks involving visual attention (Ledger, 2013). A lower EBR is hypothesised to help decrease the chance of missing critical information when focusing on tasks involving visual attention (Hoppe et al., 2018; Nakano et al., 2009; Ranti et al., 2020). This result also contrasts with existing research suggesting atypical oculomotor function, including EBR, in ADHD (Groen et al., 2017), ASD (Goldberg et al., 1987; Hornung et al., 2019) and dyslexia (Tooze, 2022; Vagge et al., 2015). It is possible that the content, duration or complexity of the learning material might not have elicited the same EBR patterns as found in previous studies. Some participants with ADHD might also have been receiving medication treatment, which can affect oculomotor function, including EBR (Fried et al., 2014).
Importantly, no difference in LP was observed between NT and ND students. University students with ADHD, ASD or dyslexia may have developed compensatory learning strategies allowing them to perform at a level comparable to their NT peers despite experiencing higher cognitive load (Alexander, 2024; Livingston et al., 2019; Sedgwick-Müller et al., 2022). The ND group also had comparable WMC to their NT peers, which has been found to positively affect learning performance and may help them succeed in their higher education studies (Swanson & Alloway, 2012). This null finding for WMC warrants consideration given prior research showing working memory impairments in ND populations (Kofler et al., 2020; Rabiee et al., 2020; Wang et al., 2022). The small sample size may have limited statistical power to detect group differences, or alternatively, university students may represent a highly selected sample who have successfully navigated educational systems requiring substantial working memory resources. This selection effect could obscure group differences that might be more apparent in broader population samples.
The observed variations in cognitive load provide preliminary evidence that ND students engage with online educational content in distinct ways, highlighting the need to consider cognitive processes alongside behavioural outcomes. Collectively, these findings suggest a nuanced relationship between neurodiversity and cognitive load in online learning and highlight the importance of evaluating multiple objective neurophysiological measures rather than solely relying on a single indicator.
However, the relatively small sample size increases the risk of findings that may not generalise to larger populations (Button et al., 2013; Cohen, 1988). Larger, more diverse samples are needed to replicate these findings and provide a more comprehensive understanding of how online learning influences cognitive load across different neurocognitive profiles, ultimately supporting the development of evidence-based, accessible online education. Additionally, future research would benefit from incorporating subjective cognitive load self-report scales (Hart & Staveland, 1988; Klepsch et al., 2017; Leppink et al., 2013; Paas, 1992) to complement objective neurophysiological measures. Such measures would enable direct comparison with established literature on cognitive load in educational settings and provide convergent validation of EEG and eye-tracking indicators. The combination of objective neurophysiological measures with subjective ratings would strengthen interpretations of cognitive load differences and facilitate translation of findings to educational practice.
An important methodological consideration highlighted by these findings is the conceptualisation of neurodiversity. Significant between-group differences emerged when comparing NT and ND students as categorised based on prior diagnosis. However, more nuanced relationships between variables were observed when examining relationships dimensionally using validated scales. This is consistent with research suggesting that cognitive load manifests heterogeneously across neurodevelopmental conditions (Papaioannou et al., 2021). Such heterogeneity in cognitive processing may contribute to null results or inconsistent correlations, as individuals within a diagnostic category can vary greatly in their cognitive profiles and responses to online learning tasks. Moreover, the relatively small sample size in this study likely exacerbated the effects of this heterogeneity, reducing the statistical power to detect subtler relationships (Button et al., 2013). In small samples, individual differences have a larger impact on group-level analyses, and trends or patterns may not reach statistical significance even if they are present in the population (Maxwell, 2004). This heterogeneity, paired with a small sample, could help explain why several correlations between ND traits and cognitive load measures failed to remain significant after correction (Lenroot & Yeung, 2013; Luo et al., 2019). Future research would benefit from larger, more diverse samples to better account for this inter-individual variability and provide a more comprehensive understanding of how cognitive load manifests across the spectrum of ND traits. An additional consideration is the demographic composition of our sample, which was predominantly female. Research suggests that men and women can differ in their responses to cognitive load (Park et al., 2017). Studies with more balanced gender distributions would help clarify whether the patterns we observed generalise across sexes or whether gender-specific interventions might be beneficial for optimising online learning experiences in ND populations.
Overall, these findings suggest a complex relationship between neurodiversity and cognitive load in online learning, and support the value of adopting dimensional, trait-based approaches alongside categorical diagnostic classifications in the study of neurodiversity, enabling the examination of cognitive load across traditional diagnostic boundaries. Importantly, several of our results, while not statistically significant after correction, showed moderate-to-large effect sizes and consistent directional patterns across independent measures (e.g., TAR, ICL, and ECL). In pilot neuroscience studies, marginal but theory-aligned findings are often the first indicators of true effects, particularly when convergent across modalities (Button et al., 2013). Overcorrecting at this stage risks discarding valuable signals that can guide future work.
While this study uncovered potential differences in how ND students experience cognitive load in online learning, there were some limitations. The current sample comprised only university students, limiting generalisability to non-academic populations. Future research with more diverse participants is necessary to explore whether similar relationships between ND traits and cognitive load exist across different online education contexts. Another limitation of the present study is the absence of medication status and intellectual quotient measures, which are known to contribute to inter-individual variance in cognitive performance (Deary et al., 2007; Milioni et al., 2017; Wolff et al., 2022; Wong & Stevens, 2012). As described in the Methods section, the design prioritised ecological validity and participant comfort by limiting data collection to variables directly relevant to the research aims. In particular, collecting detailed information about stimulant or other pharmacological treatments would have required disclosure of sensitive medical data and, given the small pilot sample, would not have enabled meaningful analysis. Nonetheless, variation in medication use may have contributed to variability within the ND group or reduced observed group differences (Swanson et al., 2011). Future studies with larger and more clinically detailed samples could explicitly assess medication type, timing and dosage to clarify their potential effects on cognitive load during online learning. Including these variables would allow for a more comprehensive understanding of the relationship between ND traits, cognitive load, and learning performance.
Our ND group included a small proportion of self-diagnosed participants alongside those with professional diagnoses. This inclusion was deliberate as formal diagnosis remains inaccessible for many students due to long wait times (Lang, 2024) and is particularly delayed in women (Quinn & Madhoo, 2014), so excluding self-identified students would therefore risk systematically under-representing key subgroups. A potential limitation is that self-diagnosis may not always reflect a neurodevelopmental condition, as overlapping symptoms can arise from other factors such as chronic stress or other environmental exposures (Erdal et al., 2025). However, all participants completed validated trait measures (ASRS, AQ and ARHQ), and the ND group scored significantly higher than NT peers across these instruments, supporting the construct validity of the grouping. We consider this inclusive approach co-designed with our Research Advisory Board to be appropriate for capturing the lived reality of university cohorts, while recognising that larger future studies could examine diagnostic pathways and potential confounds in more detail.
Although the study interface visually simulated a standard online learning environment, participants did not interact with it beyond passively watching a lecture video. This design choice allowed us to focus specifically on perceptual and attentional aspects of cognitive load during content viewing, improving interpretability of eye-tracking and EEG measures. However, this necessarily limited ecological validity as some challenges of online learning – such as task management, navigation or multitasking – were not captured. These executive demands might be especially relevant for ND students and could influence their cognitive load (Le Cunff et al., 2024b). Future work should incorporate more interactive elements to model the full range of challenges present in real-world online learning environments. In addition, the physical laboratory environment itself may have introduced limitations. ND participants may experience distinct sensory, emotional and cognitive processing differences that can impact their experimental responses. Even in a room designed to minimise distractions and standardise settings across participants, factors such as room temperature or ambient noise might be more salient for certain ND individuals (Le Cunff, Glover et al., 2024), independently of the cognitive load imparted by the experimental task.
The use of EEG features such as TAR to measure cognitive load comes itself with several limitations. While TAR has been widely used to study cognitive load, it may not fully capture the complexity of cognitive load in ND populations where neural processing patterns may differ significantly from those of NT individuals (Le Cunff et al., 2023). Additionally, TAR and similar EEG-based measures are often sensitive to individual differences in baseline neural activity, which may further complicate their interpretation in small or heterogeneous samples (Lopez et al., 2023). To mitigate these limitations, relative power measures were used, which consider the power of specific frequency bands in relation to the total power across all frequencies (Klimesch, 1999). This approach helps to account for individual differences in baseline neural activity by normalising power within each subject, providing a more stable measure across participants (Allen et al., 2004; Knyazev, 2007). However, while this method reduces variability, it may still be influenced by inter-individual differences in brain structure, neural processing or response to environmental factors – which might be particularly relevant in ND populations (Barry et al., 2003; Lenartowicz et al., 2021). Because we aimed to examine group differences during a naturalistic learning task and because standard rest conditions alter the sensory context in ways that make them difficult to use as neutral baselines for audiovisual tasks (Keil et al., 2014; Laufs et al., 2003), resting-state EEG was not collected in this pilot study. This decision was also informed by input from the Research Advisory Board to simplify the protocol and minimise participant fatigue (Le Cunff, Glover et al., 2024). However, this design choice limits the ability to determine whether observed group differences are fully task-evoked or partly reflect pre-existing neural traits. This is particularly relevant given evidence that ADHD is associated with elevated frontal theta during rest, which has been linked to cognitive performance differences (Hermens et al., 2005). Future studies could include a brief, eyes-open resting baseline matched in luminance and auditory input to the learning task to enable within-subject comparisons and better separate baseline activity from task-related neural responses in heterogeneous samples. Finally, the correlational nature of this study precludes causal conclusions.
Nevertheless, this study provides a valuable proof of concept for the methodological approach used to investigate the relationship between neurodiversity and cognitive load in online learning, and its findings offer novel insights into the distinct ways ND students engage with online educational content. In light of the increasing prevalence of online education, we think that these insights should be relevant for researchers and educators aiming to create inclusive online learning environments. While some individual differences were not as robust as expected and further research is needed to expand upon these findings, this study contributes to the body of knowledge at the intersection of neurodiversity and online education and the approach demonstrated here provides a foundation for future research that could explore these patterns in larger, more diverse samples – ultimately contributing to more inclusive, evidence-based online education for neurodiverse classrooms.
Conclusion
This pilot study developed in collaboration with ND students offers a proof of concept for investigating the relationship between neurodiversity and cognitive load in online learning using a multimodal approach. Through the integration of EEG, eye-tracking and validated trait scales, it highlights potential differences in how ND and NT learners engage with online educational content. While some effects were only marginal after statistical correction, the consistency and directionality of the results, alongside moderate-to-large effect sizes, suggest patterns that warrant further exploration. As online learning becomes increasingly widespread, understanding how different learners experience cognitive load is essential for designing accessible and effective educational materials. These preliminary results contribute to ongoing efforts to investigate online learning processes and experiences in diverse student populations and demonstrate the potential of multimodal approaches in neuroeducation research.
Supplemental Material
sj-pdf-1-ndy-10.1177_27546330251400677 - Supplemental material for Neurodiversity and Cognitive Load in Online Learning: A Pilot EEG and Eye-Tracking Study
Supplemental material, sj-pdf-1-ndy-10.1177_27546330251400677 for Neurodiversity and Cognitive Load in Online Learning: A Pilot EEG and Eye-Tracking Study by Anne-Laure Le Cunff, Eleanor J Dommett and Vincent Giampietro in Neurodiversity
Footnotes
Acknowledgements
The authors would like to thank their Research Advisory Board for providing feedback on the protocol and recruitment documents for this study. For the purposes of open access, the authors have applied a Creative Commons Attribution (CC BY) licence to any Accepted Author Manuscript version arising from this submission.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The Research Advisory Board for this study was supported by funding from UK Research and Innovation (UKRI).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.