
Abstract
Spinal cord injury (SCI) triggers secondary pathological processes, including inflammation, oxidative stress, and mitochondrial dysfunction, that contribute to progressive neuronal damage. Because mitochondria play a central role in cellular homeostasis, mitochondrial impairment is considered an important component of SCI pathophysiology. We investigated the effects of exenatide, a glucagon-like peptide-1 (GLP-1) receptor agonist, on mitochondrial dysfunction after SCI using a rat contusion model and C6 glioma cells. Exenatide was associated with increased OPA1 expression and enhanced Drp1 phosphorylation at Ser637, suggesting a shift in mitochondrial dynamics toward fusion-dominant regulation. Exenatide also tended to increase the expression of Bcl-2, HO-1, and p62. In vitro, exenatide attenuated H2O2- and CCCP-induced reductions in cell viability, preserved mitochondrial membrane potential, and reduced the proportion of dead cells. These findings suggest that exenatide may mitigate secondary injury after SCI in association with mitochondrial protective responses.
Introduction
Spinal cord injury (SCI) is a devastating form of central nervous system (CNS) trauma that imposes substantial physical, psychological, and vocational burdens on affected individuals and their families.1,2 SCI can be categorized into primary and secondary injuries: the former results directly from the initial traumatic mechanical insult, whereas the latter arises from a complex cascade of vascular, cellular, and biochemical events. This cascade induces necrosis and apoptosis in surrounding cells, thereby contributing to the progressive expansion of tissue damage. 3
Inflammation represents a key element of secondary injury, playing a pivotal role in shaping the pathological progression of both acute and chronic SCI, as well as influencing the body’s regenerative capacity. 4 Following SCI, the inflammatory response in the CNS is mediated by peripherally derived immune cells and activated glial cells that proliferate and migrate to the lesion site. 5 Infiltrating inflammatory cells promote excessive production of reactive oxygen species (ROS), which are released into the extracellular space and contribute to extensive cell death involving neurons, glial cells, and oligodendrocytes. Inflammation and oxidative stress are key components of secondary injury, underscoring the clinical importance of limiting these pathological responses. 6
A glucagon-like peptide-1 (GLP-1) receptor agonist has been shown to exert neuroprotective effects in a variety of neurodegenerative disorders.7–9 In our previous studies using a rat model of SCI, exenatide, a GLP-1 receptor agonist, alleviated endoplasmic reticulum (ER) stress, promoted macrophage polarization, and improved hindlimb motor function.10,11
Mitochondria serve as the principal bioenergetic organelles in most cells; however, secondary injury after SCI impairs mitochondrial function, leading to reduced adenosine triphosphate (ATP) production, increased ROS generation, and activation of mitochondria-mediated apoptotic signaling pathways. These changes ultimately contribute to progressive neuronal cell death and tissue damage.12,13 Mitochondria continuously undergo fusion and fission, thereby maintaining their morphology and quality. Fusion refers to the process in which two mitochondria merge into a single organelle through the joining of their outer and inner membranes. This process promotes the exchange of mitochondrial DNA, proteins, and metabolic intermediates. 14 In contrast, fission can be understood as the division of mitochondria to increase mitochondrial number and is therefore the process opposite to fusion. Fusion and fission are not independent events; rather, excessive mitochondrial fission accelerates mitochondrial damage.15,16 In addition, post-injury oxidative stress disrupts mitochondrial membrane potential, amplifies ROS generation, and impairs ATP synthesis through cytoplasmic calcium (Ca2+) accumulation.17,18 Furthermore, the activation and increased permeability of mitochondrial transition pores induce mitochondria-mediated apoptosis. 19
In the myocardium, GLP-1 receptor (GLP-1R) signaling has been reported to exert cardioprotective effects through the Epac/PI3K/Akt pathway. 20 GLP-1R transcripts and protein are expressed in neuronal cell bodies throughout the gray matter, with particularly high levels of GLP-1R protein observed in neurons that contact the cerebrospinal fluid. 21 GLP-1R is also regarded as an organizer of signaling nanodomains, or signalosomes, and has been suggested to improve mitochondrial function through the regulation of Ca2+ and ATP fluxes. 22
Taken together, these findings suggest that GLP-1R signaling may influence mitochondrial homeostasis in the CNS, providing a rationale for examining whether exenatide affects mitochondrial dysfunction after SCI. To date, no research has investigated how exenatide influences mitochondrial dysfunction following SCI.
Materials and Methods
Rat model of SCI
All animal procedures were performed in accordance with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals and were approved by the Animal Experimentation Committee of Tokai University School of Medicine (approval no. 231051). Ten-week-old female Sprague–Dawley (SD) rats were obtained from CLEA Japan Inc. (Kanagawa, Japan). Under anesthesia with 4% isoflurane, the back was shaved and prepared under aseptic conditions. Using the same surgical procedure as in our previous experiments, a midline skin incision was made, and the subcutaneous tissue and paraspinal muscles were carefully dissected to expose the lower thoracic vertebrae. After laminectomy at the 10th thoracic vertebra, contusive SCI was induced using an Infinite Horizon Impactor (Precision Systems & Instrumentation, Lexington, KY, USA) with a force of 0.2 N. The paraspinal fascia and skin were then sutured, and the animals were allowed to recover from anesthesia.10,11
For the in vivo experiments, animals were assigned to three groups: a sham group (S group), which underwent laminectomy without spinal cord contusion; a control group (C group), which underwent SCI and received phosphate-buffered saline (PBS); and an exenatide-treated group (Ex group), which underwent SCI and received exenatide. To determine the dose used in the subsequent analyses, exenatide was initially administered subcutaneously at 10, 20, or 40 μg/day twice daily beginning immediately after SCI. Based on the behavioral results, 40 μg/day was selected for the subsequent experiments and was defined as the Ex group. Animals in the C group received 0.1 mL PBS per dose on the same schedule. The exenatide used in this study was Byetta® (AstraZeneca Pharmaceuticals LP, Cambridge, UK), an approved therapeutic agent for type 2 diabetes mellitus. Although the dose used in rats is not directly translatable to safe administration in humans, it was selected to evaluate the therapeutic potential of exenatide in this pre-clinical SCI model.
To manage post-injury urinary dysfunction, bladder expression was performed twice daily until spontaneous voiding was confirmed. Animals showing signs of cystitis or wound infection were excluded from the study. All animals were handled and housed in accordance with the protocol approved by the Life Science Support Center of Tokai University School of Medicine.
Sample collection
At 3 and 7 days post-injury (dpi), rats were reanesthetized with 4% isoflurane, and a 5-mm segment of spinal cord tissue centered on the lesion site was harvested under microscopic observation. The excised tissue was immediately rinsed with ice-cold PBS. The samples were then homogenized using a ShakeMaster NEO homogenizer (BMS, Japan), and the resulting homogenates were aliquoted for RT-qPCR and Western blot analyses.
RT-qPCR
Total RNA was extracted from spinal cord tissue using the RNeasy Mini Kit (Qiagen, Japan) and reverse-transcribed into cDNA using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems™, Thermo Fisher Scientific, Japan). RT-qPCR was performed using a LightCycler® 480 system (Roche Diagnostics, Japan) with Fast SYBR™ Green Master Mix (Applied Biosystems™, Thermo Fisher Scientific, Japan). The expression levels of Drp1, OPA1, Bcl-2, HO-1, and p62 were analyzed, and β-actin was used as an internal control. The following primers were designed for the Rattus norvegicus genome: DRP1 (forward: 5′-ACAACAGGAGAAGAAAATGGAGT-3′, reverse: 5′-GATGGATTGGCTCAGGGCTT-3′), OPA1 (forward: 5′-TTCTTCACTGCGGGTACACCT-3′, reverse: 5′-CTTCCTGTGGTGGATGCCTT-3′), Bcl-2 (forward: 5′-AGGATAACGGAGGCTGGGATG-3′, reverse: 5′-CTCACTTGTGGCCCAGGTAT-3′), HO-1 (forward: 5′-GCTAGCCTGGTTCAAGATACTACC-3′, reverse: 5′-CGCCAACAGGAAACTGAGTG-3′), p62 (forward: 5′-GCTGAGTCGGCTTCTGCT-3′, reverse: 5′-ACTTTCTGCAGAGGTGGGTG-3′), and β-actin (forward: 5′-CCGCGAGTACAACCTTCTTG-3′, reverse: 5′-CGTCATCCATGGCGAACTGG-3′). For each sample, measurements from two wells were averaged, and spinal cord samples from five animals per group were analyzed. Relative gene expression was calculated using the 2−ΔΔCt method after normalization to the mean Ct value of β-actin. Expression levels in the C and Ex groups are presented relative to those in the S group, which was set to 1.
Western blot assay
Excised spinal cord samples were immediately washed with ice-cold PBS, and the dura mater was removed as completely as possible. Protein extracts were prepared using freshly made lysis buffer containing 50 mM Tris/HCl (pH 7.4), 1 mM CaCl2, 0.5 mM phenylmethylsulfonyl fluoride, and 1% Nonidet-P (NP)−40, followed by sonication with an ultrasonic device (BIORUPTOR, Sonic Bio Inc., Samukawa, Japan). The lysates were centrifuged at 12,000 rpm for 5 min at 4°C, and proteins were collected from the supernatants. All extraction procedures were performed on ice, and the samples were stored at −80°C until analysis. Protein concentration was determined using DC Protein Assay Reagent (BIO-RAD, #1310-73-2, #5000114, and #500-0115) and measured at 700 nm with a microplate reader (VersaMax™, MOLECULAR DEVICES).
Protein samples were mixed with 5 × sample buffer and 2-mercaptoethanol (2-ME), heated at 100°C for 5 min, and separated on polyacrylamide gels using MES running buffer (Invitrogen NuPAGE™, MES SDS Running Buffer [20×], NP0002) at 20 mA for 80 min. After electrophoresis, proteins were transferred onto a polyvinylidene fluoride membrane (Immobilon®-P, 0.45 μm; Merck Millipore Ltd., Burlington, VT, USA) at 20 V for 60 min. The membranes were washed three times for 10 min each, blocked with 3% bovine serum albumin (BSA) (Probumin® Bovine Serum Albumin Universal Grade; Merck Millipore Ltd.) at 37°C for 30 min, and then washed with Buffer G containing 0.1% gelatin, 0.1% BSA, 1 mM MgCl2, and 0.1% NaN3 in PBS. The membranes were incubated overnight at 4°C with the following primary antibodies: DRP1 (abcam, ab184247, 1:1000), phospho-DRP1 (Ser637) (Cell Signaling, #6319, 1:1000), OPA1 (abcam, ab157457, 1:1000), Bcl-2 (abcam, ab59348, 1:1000), p62/SQSTM1 (MBL, PM045, 1:1000), and HO-1 (abcam, ab68477, 1:1000). After washing with TBST (50 mM Tris, pH 7.6, 150 mM NaCl, 0.1% Tween-20), the membranes were incubated for 60 min at room temperature with horseradish peroxidase-conjugated anti-rabbit IgG (GE Healthcare, NA9340, 1:3000) or anti-mouse IgG (GE Healthcare, NA9310, 1:3000). Protein signals were detected using a FUSION Solos imaging system (Vilber, Paris, France). β-actin was used as the loading control. Spinal cord samples from five animals per group were analyzed. Protein expression levels in the C and Ex groups are presented relative to those in the S group, which was set to 1.
Behavioral analysis
Hindlimb motor function was evaluated using the Basso, Beattie, and Bresnahan (BBB) locomotor rating scale before injury, on the day after surgery, and daily thereafter up to 7 dpi. 23
Behavioral assessments were performed between 9:00
Blood glucose level and body weight
Blood glucose levels were measured immediately before injury, immediately after injury, on the day after surgery, and daily thereafter up to 7 dpi. Approximately 1.1 μL of blood was collected from the caudal vein using a 20-G needle, and glucose levels were measured with a LAB Gluco blood glucose meter (Research & Innovation Japan Inc., Chiba, Japan). Body weight was recorded immediately before injury, immediately after injury, and at 3 and 7 dpi. Five animals were analyzed in each group.
Cell culture and culture conditions
The rat C6 glioma cell line was obtained from the American Type Culture Collection (Manassas, VA, USA). Cells were seeded in 10-cm tissue culture dishes and maintained at 37°C in a humidified atmosphere of 5% CO2 and 95% air for 48 h. The culture medium consisted of Dulbecco’s modified Eagle’s medium (DMEM; Gibco/Life Technologies, Carlsbad, CA, USA) supplemented with 10% (v/v) fetal bovine serum and 1% (v/v) penicillin–streptomycin.
Cell proliferation assay
When the cells reached 70–80% confluence, they were detached by trypsinization and adjusted to a density of 5,000 cells per well. A total of 120 μL of cell suspension was seeded into each well of a 96-well plate and incubated for 48 h. Excessive ROS production is a major contributor to secondary injury after SCI. 24 H2O2 was therefore used as an in vitro model of oxidative stress. Cells were then treated with exenatide or vehicle in the presence of 0–500 μM H2O2 or 0–100 μM carbonyl cyanide m-chlorophenyl hydrazone (CCCP) for 2 h. After treatment, 10 μL of Cell Counting Kit-8 (CCK-8) solution (CK04; Dojindo, Kumamoto, Japan) was added to each well, and the optical density at 450 nm was measured after 1 h of incubation using a microplate reader. Each condition was assessed in 5 wells.
Measurement of mitochondrial membrane potential and dead cells
Mitochondrial membrane potential (MMP) was assessed by flow cytometry using the MT-1 MitoMP Detection Kit (Dojindo). Briefly, rat C6 glioma cells were stained with MT-1 dye for 30 min at 37°C and then incubated for 2 h with DMEM or 50 μM CCCP. After treatment, the supernatant was collected and centrifuged. The adherent cells were washed three times with 1× PBS, detached with trypsin/EDTA, and combined with the recovered supernatant fraction. Relative cellular fluorescence was quantified using a BD LSRFortessa™ Cell Analyzer (BD Biosciences, Franklin Lakes, NJ, USA), and the data were analyzed with BD FACSDiva v9.0 and FlowJo v10. To detect dead cells, propidium iodide (PI; Sigma-Aldrich, USA) was prepared as a 1 mg/mL stock solution in PBS and added to each sample at a final dilution of 1:1000. The proportion of dead cells was subsequently determined by flow cytometry. To confirm reproducibility, the experiments were performed twice independently with three samples per experiment, and the combined data (total n = 6) were used for analysis.
Statistical analysis
All statistical analyses were performed using GraphPad Prism version 10.3.0 (GraphPad Software, CA, USA). Data are presented as mean ± standard deviation (SD). Differences between two groups were analyzed using a two-tailed unpaired Student’s t-test. Comparisons among multiple groups were performed by one-way or two-way analysis of variance (ANOVA), as appropriate, followed by Tukey’s multiple comparisons test. A p-value <0.05 was considered statistically significant.
Results
Exenatide group demonstrated improved hindlimb motor function
The BBB scale evaluates the motor functionality of the hindlimbs using a range from 0 to 21, where elevated scores indicate superior motor performance. Exenatide was delivered subcutaneously twice daily at doses of 10, 20, and 40 μg/day. At 7 dpi, the 40 μg/day group showed significantly improved hindlimb motor function (p < 0.05, Fig. 1A). The administration of exenatide for subsequent experimental trials involving rats was established at a dosage of 40 μg/day, with this cohort designated as the Ex group. The initial BBB scale scores in both the Ex and C groups started at 0 immediately after the injury and demonstrated a progressive improvement over time. Nevertheless, from 5 dpi onward, a pronounced enhancement in the scores was noted within the Ex group compared with the C group (p < 0.05, Fig. 1B).
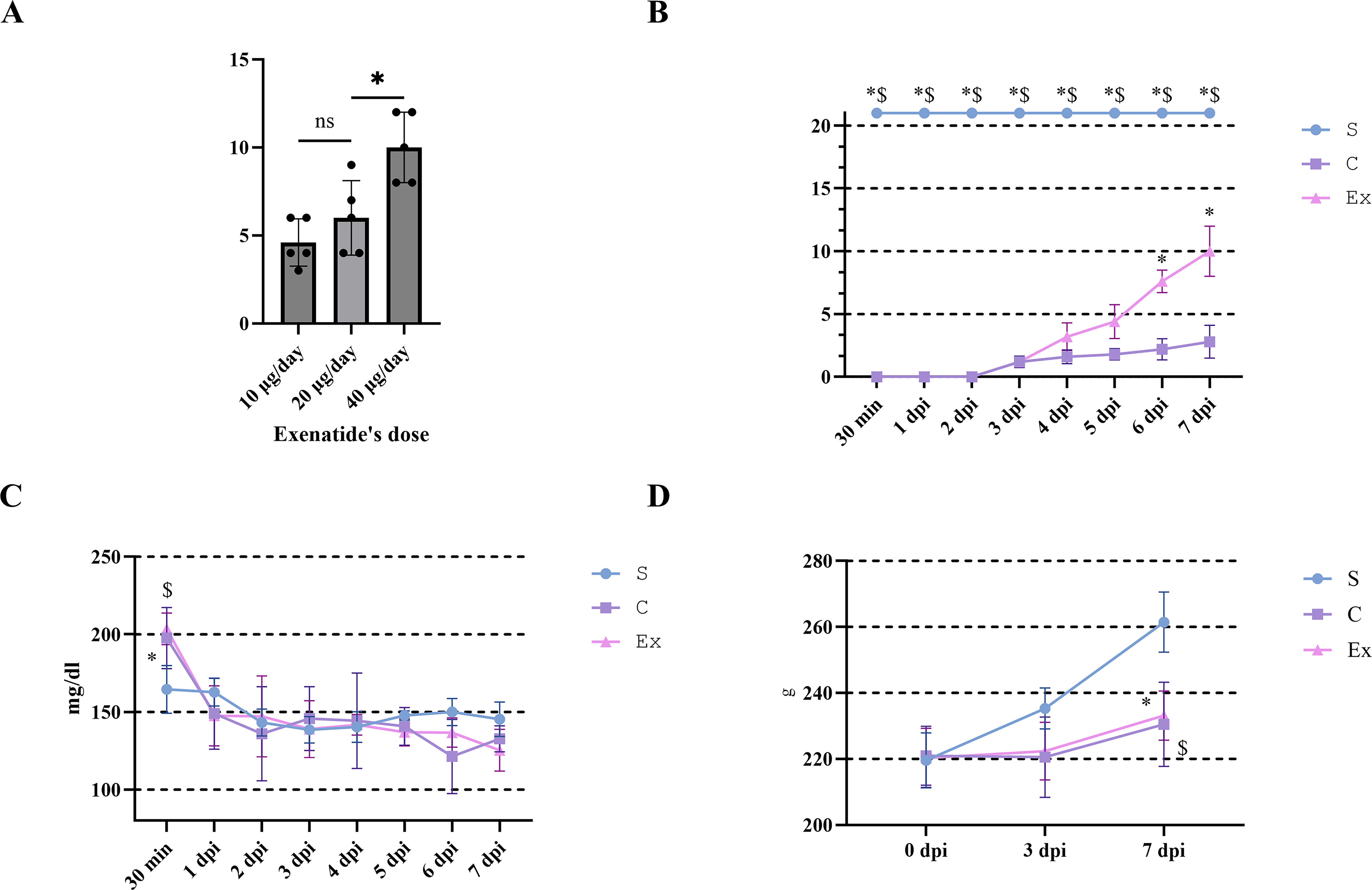
Effects of exenatide on hindlimb motor function, blood glucose, and body weight after SCI.
Exenatide did not cause excessive decreases in blood glucose or body weight after SCI
Blood glucose levels, which are normally approximately 120 mg/dL in Rattus norvegicus, increased to nearly 200 mg/dL at 30 min after injury in both groups and then rapidly returned to baseline. No significant differences were observed between the groups from 1 dpi onward, indicating that exenatide did not induce an excessive reduction in blood glucose levels (Fig. 1C). Body weight was significantly reduced in both groups compared with the sham group at 7 dpi. However, no significant difference was detected between C and Ex groups, suggesting that exenatide did not exacerbate SCI-associated weight loss (Fig. 1D).
Effects of exenatide on Drp1 and OPA1 in a rat model of SCI
Spinal cord samples were analyzed by RT-qPCR and Western blotting to assess the expression of Drp1 and OPA1, two key regulators of mitochondrial fission and fusion. Drp1 mRNA expression was significantly lower in the C group than in the sham group at both time points, whereas it was significantly higher in the Ex group than in the C group (Fig. 2A). The ratio of phosphorylated Drp1 (Ser637) to total Drp1, an indicator of fission suppression, did not differ significantly between the sham and C groups, but was significantly increased in the Ex group at 7 dpi (Fig. 3A). OPA1 mRNA expression was significantly decreased in the C group compared with the sham group at both 3 and 7 dpi. Although no significant differences were detected between the C and Ex groups, OPA1 expression tended to be higher in the Ex group at both time points (3 dpi: p = 0.54; 7 dpi: p = 0.46; Fig. 2B). At the protein level, OPA1 was significantly downregulated in the C group at both time points. Notably, exenatide significantly restored OPA1 protein expression at 3 dpi, while a nonsignificant trend toward recovery was observed at 7 dpi (p = 0.08; Fig. 3B). Together, these findings suggest that exenatide may shift mitochondrial dynamics toward fusion rather than fission, help preserve cristae structure, and thereby contribute to the suppression of apoptosis.
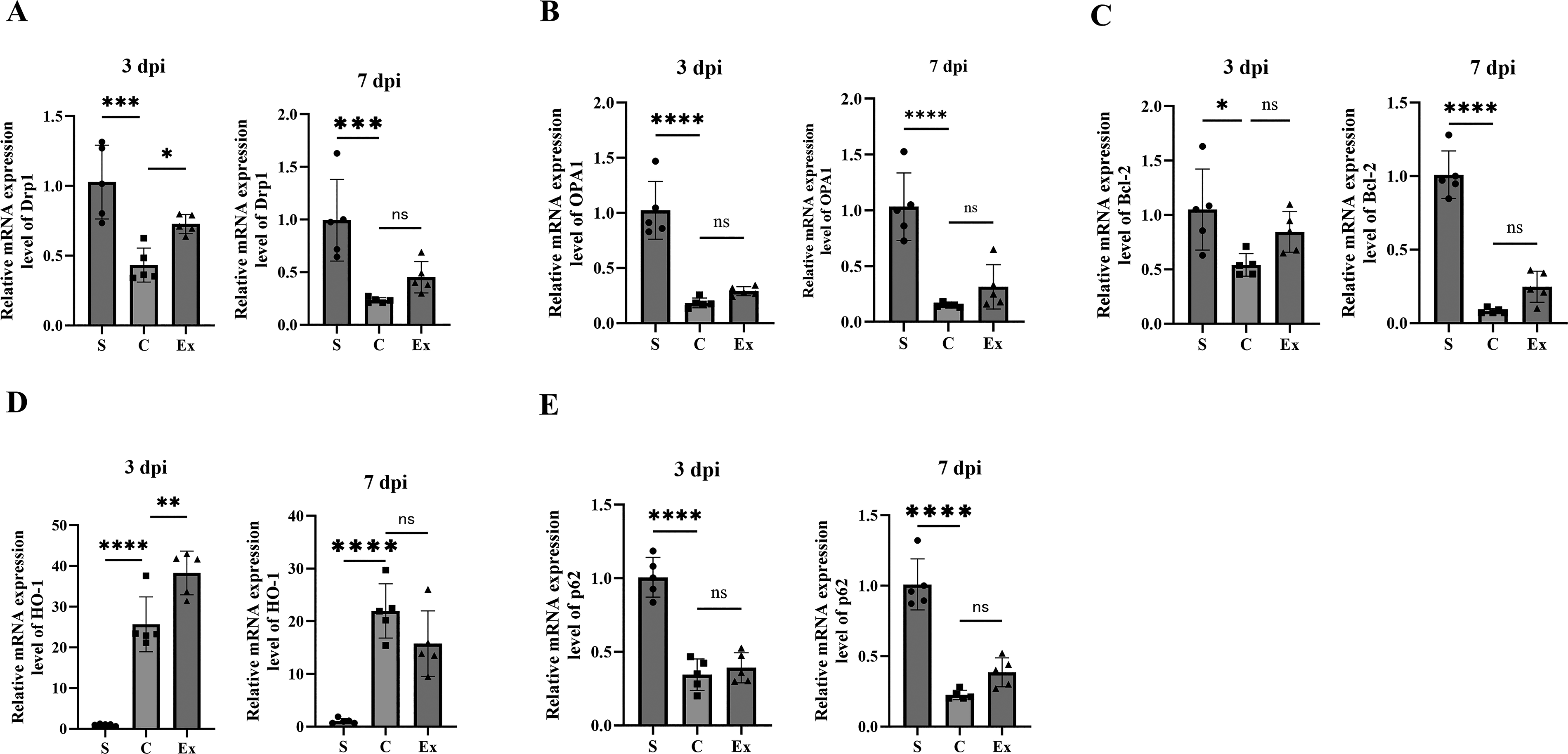
Effects of exenatide on mRNA expression of genes related to mitochondrial dynamics and mitophagy in injured spinal cords. RT-qPCR was performed to assess the mRNA expression of
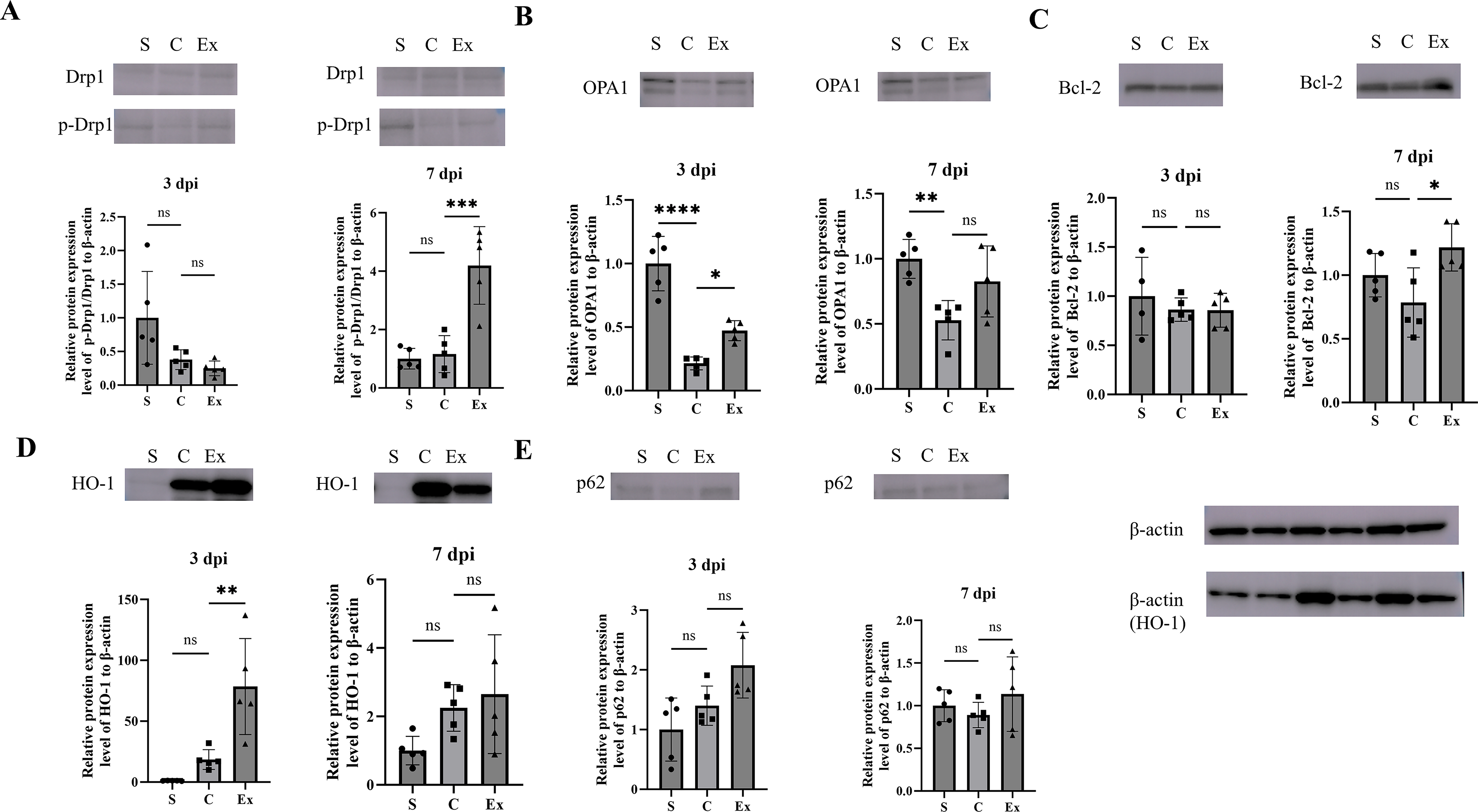
Effects of exenatide on the expression of markers related to mitochondrial dynamics and mitophagy in injured spinal cords. Spinal cord samples were analyzed by Western blot at 3 and 7 dpi to evaluate the protein expression levels of
Effects of exenatide on Bcl-2, HO-1, and p62 expression in a rat model of SCI
Spinal cord samples were analyzed by RT-qPCR and Western blotting to evaluate the expression of Bcl-2, HO-1, and p62, which are associated with mitochondrial dynamics and mitophagy. Bcl-2 mRNA expression was significantly lower in the C group than in the S group at both 3 and 7 dpi. Although no significant differences were observed between the C and exenatide-treated (Ex) groups, Bcl-2 expression tended to be higher in the Ex group at both time points (3 dpi: p = 0.17; 7 dpi: p = 0.09; Fig. 2C). At the protein level, Bcl-2 expression did not differ significantly among the groups at 3 dpi; however, at 7 dpi, Bcl-2 protein levels were significantly increased in the Ex group compared with the C group, suggesting enhancement of antiapoptotic signaling by exenatide (Fig. 3C). HO-1 mRNA expression was significantly elevated in the C group relative to the S group at both time points and was further increased in the Ex group at 3 dpi. Specifically, HO-1 mRNA levels were significantly higher in the Ex group than in the C group at 3 dpi, whereas no significant difference was detected at 7 dpi (3 dpi: p < 0.05; 7 dpi: p = 0.13; Fig. 2D). A similar pattern was observed at the protein level, with significantly higher HO-1 expression in the Ex group at 3 dpi but not at 7 dpi (3 dpi: p < 0.05; 7 dpi: p = 0.84; Fig. 3D). These findings suggest that exenatide may enhance the early protective response to oxidative stress after SCI, although whether this effect is sustained over time remains unclear. In contrast, p62 mRNA expression was significantly reduced in the C group compared with the S group at both time points. No significant differences were found between the C and Ex groups at either 3 or 7 dpi, although p62 expression tended to be higher in the Ex group at both time points (3 dpi: p = 0.80; 7 dpi: p = 0.13; Fig. 2E). At the protein level, no significant differences were observed between the C and Ex groups at either time point; however, p62 expression consistently showed an upward trend in the Ex group (3 dpi: p = 0.10; 7 dpi: p = 0.40; Fig. 3E). Collectively, these results suggest that exenatide may facilitate the clearance of damaged cellular components through autophagy-related pathways, thereby contributing to cytoprotection.
Exenatide attenuates H2O2- and CCCP-induced apoptosis and MMP loss in C6 glioma cells
H2O2 was applied at increasing concentrations for 24 h, and cell viability was assessed using CCK-8 assay. The treatment with 400 μM and 500 μM H2O2 significantly reduced cell viability compared with the control group (p < 0.05, Fig. 4A). When 200 nM exenatide was coadministered with 500 μM H2O2, cell viability was significantly improved compared with H2O2 treatment alone (p < 0.05, Fig. 4B). Based on these results, 200 nM exenatide was used in the subsequent in vitro experiments.
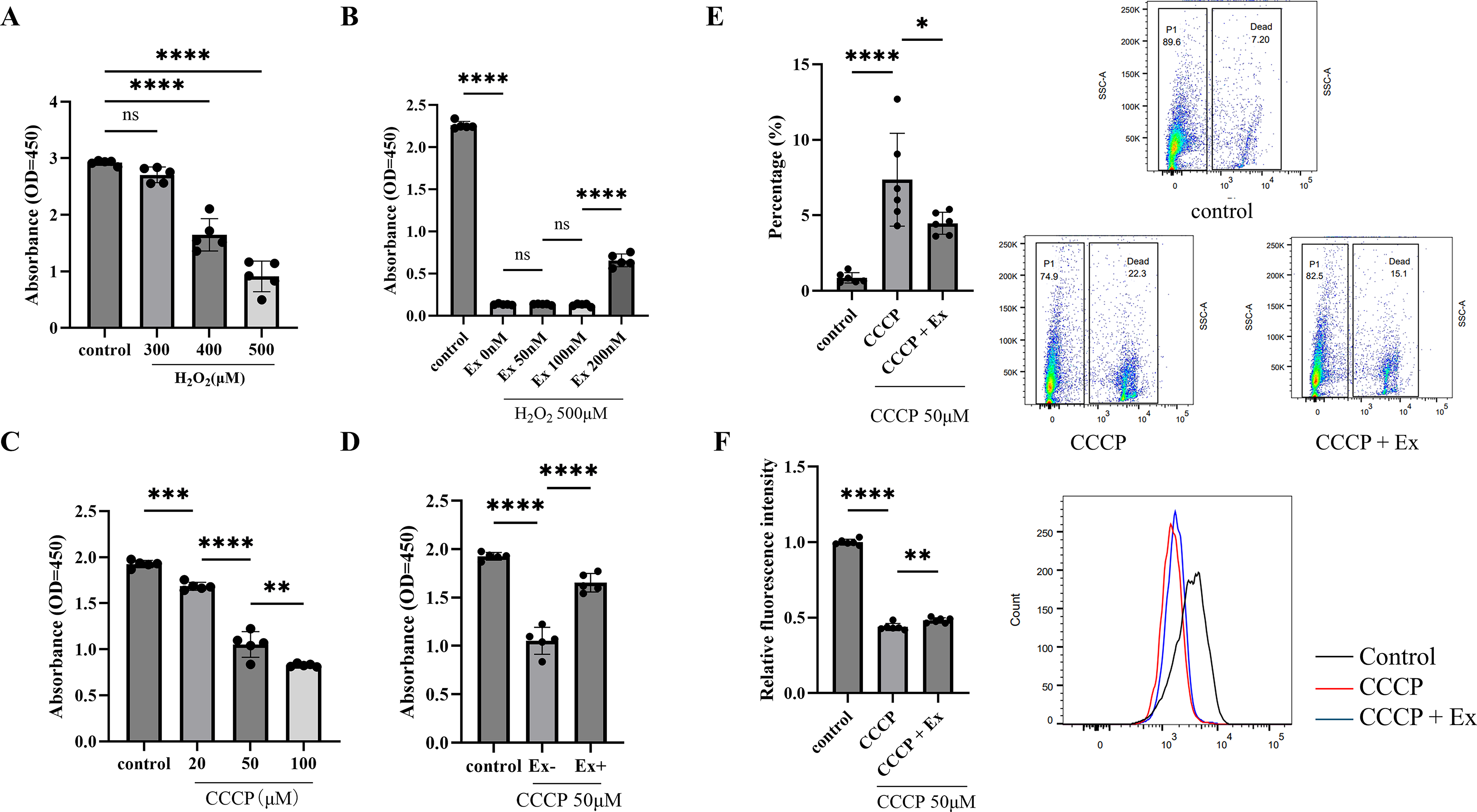
Effects of exenatide on cell viability, mitochondrial membrane potential, and cell death in C6 glioma cells exposed to H2O2 or CCCP.
CCCP was then applied at concentrations of 0–100 μM for 2 h, and cell viability was assessed in the same manner. All CCCP-treated groups showed significantly lower cell viability than the control group (p < 0.05, Fig. 4C). In cells exposed to 50 μM CCCP, cotreatment with 200 nM exenatide significantly improved cell viability compared with CCCP treatment alone. In cells exposed to 50 μM CCCP, cotreatment with 200 nM exenatide significantly improved cell viability compared with CCCP treatment alone (p < 0.05, Fig. 4D). Under the same conditions, exenatide significantly preserved mitochondrial membrane potential and reduced the proportion of dead cells (Fig. 4E, F).
Discussion
Our previous studies have shown that ER stress is an important component of the secondary injury cascade after SCI and that enhancement of the ER stress response is associated with improved motor recovery.25–29 We first examined whether amiloride, a potassium-sparing diuretic and sodium channel blocker, could enhance the ER stress response. Amiloride reduces apoptosis in oligodendrocyte precursor cells, promotes remyelination by facilitating their differentiation from surviving precursor cells, and improves motor function after SCI. 27 However, diuretics may not be suitable for patients in the acute phase of SCI because SCI-related autonomic dysfunction can destabilize blood pressure and circulation. Because GLP-1 receptor agonists have been reported to enhance the ER stress response in myocardial ischemia, cerebral ischemia, renal injury, and hepatic injury,30–33 we focused on exenatide, a GLP-1 receptor agonist that can cross the blood–brain barrier, in the present SCI model. In our previous work, exenatide reduced ER stress, promoted macrophage polarization, and improved hindlimb motor recovery after SCI. Mitochondria are a major source of ROS and play a central role in oxidative stress. They also contribute to intracellular iron homeostasis.34,35 Damaged mitochondria are removed or recycled through fusion, fission, and mitophagy, thereby maintaining mitochondrial network integrity through mitochondrial quality control.
Mitochondrial dysfunction is considered an important contributor to secondary injury after spinal cord trauma. Abnormalities in mitochondrial morphology and dynamics can impair mitochondrial function and compromise neuronal survival.36,37 Mitochondrial fusion and fission are essential for maintaining mitochondrial integrity and energy production in neurons. Previous studies have identified key regulators of these processes, including OPA1, Drp1, and Bcl-2, and have suggested that their modulation may represent a potential therapeutic target.38,39 Mitochondrial dynamics depend on the balance between fusion, mediated by proteins such as OPA1 and mitofusins, and fission, which is primarily regulated by Drp1 and its associated proteins.36,40 After SCI, increased Drp1 activity together with reduced or cleaved OPA1 has been reported, indicating disruption of this balance.38,41 Phosphorylation of Drp1 at Ser637 is an important regulatory event that suppresses mitochondrial fission and has been associated with neuroprotection. In the present study, exenatide was associated with partial restoration of the disturbed balance between Drp1 and OPA1 expression after SCI. These findings suggest that exenatide may shift mitochondrial dynamics toward fusion-dominant regulation and thereby help limit mitochondrial damage and cell death after injury.
HO-1 is involved in the cellular response to oxidative stress and has been linked to cytoprotective and anti-apoptotic processes. In SH-SY5Y neuronal cells, activation of the Keap1/Nrf2/HO-1 signaling pathway has been shown to attenuate oxidative stress-induced apoptosis by reducing ROS levels and stabilizing mitochondrial membrane potential. 42 HO-1 has also been reported to influence the expression of apoptosis-related proteins. In human retinal pigment epithelial cells, upregulation of HO-1 suppresses caspase-3 activation while increasing the expression of antiapoptotic proteins such as Bcl-2. 43 In the present study, exenatide enhanced HO-1 expression at 3 dpi, which may reflect an early protective response to oxidative stress after SCI. However, because this increase was not maintained at 7 dpi, the longer-term significance of this response remains unclear.
p62 is also involved in mitophagy, a selective form of autophagy that targets damaged mitochondria for degradation. Dysregulation of p62-mediated mitophagy can aggravate mitochondrial dysfunction and has been implicated in neurodegenerative disorders.44,45 In the present study, exenatide was associated with a tendency toward increased p62 expression at both the mRNA and protein levels, although these changes did not reach statistical significance. This finding may indicate a potential relationship between exenatide treatment and autophagy-related clearance of damaged cellular components. However, because p62 can have context-dependent roles and the present study did not directly assess mitophagic flux, the contribution of p62 to the observed protective effects should be interpreted with caution. MMP reflects mitochondrial functional and metabolic states. Disruption of MMP impairs ATP production, disturbs calcium homeostasis, and increases ROS generation.46,47 These changes can lead to excitotoxicity, oxidative damage, and activation of necrotic and apoptotic pathways. 48 MMP is also closely related to mitochondrial ROS production, and reduced MMP has been associated with impaired respiratory chain activity and enhanced oxidative stress in mitochondrial disorders.49–53 In addition, loss of MMP is generally associated with neuronal cell death.54,55 In our in vitro experiments, exenatide attenuated the CCCP-induced decline in MMP and reduced the proportion of dead cells. These findings support the possibility that exenatide helps preserve mitochondrial function under conditions of mitochondrial stress.
We used the C6 rat glioma cell line as an in vitro model to examine cellular responses to oxidative and mitochondrial stress in the presence of exenatide. Although C6 glioma cells are derived from a glial tumor and do not fully reproduce the behavior of primary neural cells, they provide a reproducible system for the initial evaluation of mitochondrial dysfunction, oxidative stress, and cell viability. Because glial responses, including reactive astrogliosis and neuroinflammation, are important components of SCI pathology, the use of a glial cell line may still provide useful preliminary information. Nevertheless, these in vitro findings should be regarded as complementary to the in vivo observations rather than as a definitive model of SCI pathophysiology. Further studies using primary astrocytes, oligodendrocyte precursor cells, neurons, or spinal cord-derived cell populations will be necessary to extend these findings.
Taken together, the present findings suggest that exenatide is associated with improved hindlimb motor recovery after SCI and with molecular changes related to mitochondrial dynamics and mitochondrial homeostasis. In particular, exenatide treatment was associated with increased OPA1 expression, enhanced Drp1 phosphorylation at Ser637, preservation of MMP, and reduced cell death under mitochondrial stress conditions. These in vitro findings complement the in vivo observations and support a potential link between exenatide treatment and mitochondrial protective responses after SCI.
This study has several limitations. Because the in vivo analyses were performed using homogenized spinal cord tissue, we could not identify the specific cellular targets of exenatide in the injured spinal cord. Histological assessments, including lesion volume, tissue sparing, axonal preservation, and cell-specific staining, were also not performed, and therefore, the molecular changes observed here could not be directly linked to structural tissue protection in vivo. In addition, the in vitro experiments were conducted using C6 glioma cells. Although this model was useful for the initial assessment of oxidative stress-related and mitochondrial stress-related responses, it does not fully reflect the properties of primary neural cells. Finally, we did not perform in vivo pathway-specific intervention studies to determine whether mitochondrial regulation is required for the protective effects of exenatide. For this reason, our findings should be regarded as showing an association between exenatide treatment and mitochondrial protective responses after SCI, rather than proving a causal mechanism. Additional studies using histological analyses, primary neural cell systems, and in vivo mechanistic interventions will be needed to define these relationships more clearly.
Transparency, Rigor, and Reproducibility
All animal experiments were approved by the Animal Experimentation Committee of Tokai University School of Medicine and conducted in accordance with the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. Statistical analyses were performed using GraphPad Prism version 10.3.0, as described in the Methods section. A sample size of five animals per group was selected based on an anticipated effect size of 1.5, which was estimated to provide 80% power to detect a significant difference using two-way analysis of variance with repeated measures on one factor. A p-value <0.05 was considered statistically significant. Research team members who were familiar with the relevant characteristics of the animals were involved in animal handling. One team member performed SCI induction, and T.K. was blinded during the experimental procedures and data analysis. All equipment and analytical reagents used in this study are described in the Materials and Methods section. Replication experiments are ongoing.
Authors’ Contributions
T.K.: Writing—original draft, conceptualization, data curation, investigation, and methodology. H.K.: Writing, reviewing, and editing, data curation, funding acquisition, methodology, project administration, supervision, validation, and visualization. K.S.: Writing, reviewing, and editing, conceptualization, data curation, investigation, and methodology. N.H.: Writing, reviewing, editing, validation, and visualization. K.O.: Writing, reviewing, editing, validation, and visualization. M.W.: Supervision, writing, review, editing, and funding acquisition.
Footnotes
Acknowledgments
The authors would like to express their profound gratitude to the personnel of the Life Science Support Center (Tokai University, Japan) for their invaluable technical support in protein quantification and fluorescence-activated cell sorting.
Funding Information
The author(s) declare that financial support was received for the research, authorship, and publication of this article. This work was supported by JSPS KAKENHI Grant-in-Aid for Scientific Research (C) numbers