
Abstract
CHD7 and CHD8 are chromatin remodeling proteins that regulate several neurodevelopmental events. Mutations in these chromatin remodeling genes occur in neurodevelopmental disorders including CHARGE Syndrome and Autism Spectrum Disorders. Kismet (Kis) is the sole Drosophila homolog of CHD7 and CHD8. We investigated the possibility that Kis influences retrograde synaptic signaling given that Kis restricts the synaptic levels of several cell adhesion molecules and facilitates endocytosis. Our data indicate that Kis restricts synaptic pMad while facilitating the localization of pMad to presynaptic motor neuron nuclei. While the increase in pMad at kis mutant synapses may contribute to the loss of Endophilin B, it may not influence the mislocalization of glutamate receptors relative to active zones or the locomotor phenotypes observed in kis mutants. Kis may antagonize Polycomb Repressive Complex 2 (PRC2) signaling to restrict synaptic pMad. Kis, including its chromatin remodeling/ATPase activity, is required in presynaptic motor neurons for proper synaptic pMad levels. In contrast, an ATPase-deficient Kis can rescue synaptic pMad when expressed in all tissues. Similarly, expression of human CHD7 in all tissues of kis mutants rescues synaptic pMad. Our data suggest a model where Kis restricts synaptic pMad both by transcription-dependent and transcription-independent mechanisms. These data may aid in a better understanding of the importance of chromatin remodeling for synaptic structure and function and the molecular changes correlated with neurodevelopmental disorders.
Introduction
The nervous system is composed of an intricate array of neurons and glial cells, which collectively mediate diverse processes including perception, movement, and higher cognitive processes such as learning and memory. Neuronal communication consists not only of anterograde signaling to postsynaptic cells but also retrograde signaling and signaling between neurons and glia. 1 Retrograde signaling mechanisms provide feedback to modulate presynaptic functions and use signaling molecules like brain-derived neurotrophic factor (BDNF), 2 corticosteroids, 3 endocannabinoids, 4 and the cell adhesion molecule Neuroligin 3, 5 among others. At the Drosophila neuromuscular junction (NMJ), a model glutamatergic synapse similar to mammalian central nervous system (CNS) synapses, 6 retrograde bone morphogenetic protein (BMP) signaling enhances synaptic growth, stability, and plasticity. 7
BMP ligands and receptors belong to the transforming growth factor-beta (TGF-β) superfamily, which is highly conserved across metazoan evolution. 8 BMP signaling components are expressed in several tissues throughout development and adulthood. 9 As such, BMP signaling is critical for diverse developmental processes such as bone formation, 10 embryonic body plan establishment, 11 and limb patterning. 12 In the developing nervous system, BMP signaling helps establish the anterior-posterior and dorsal-ventral brain axes 13 and restricts neuronal migration in the cortex. 14 Although BMP ligands and receptors are expressed in adult brains and spinal cords, the role of BMP signaling in the adult nervous system is poorly understood. 9
BMPs are secreted as dimers and mediate signaling between neurons, muscles, and glia. 15 The BMP ligand glass bottom boat (Gbb), which is most similar as BMP7, 6 is secreted from postsynaptic muscles of Drosophila larvae. 16 Gbb binds to presynaptic tetrameric receptors that include the Type II receptor Wishful Thinking (Wit) in a complex with the Type I receptors Saxophone or Thick veins (Tkv). Activated receptors phosphorylate the R-Smad, Mothers against decapentaplegic (Mad). 8 Phosphorylated Mad (pMad) binds with the Co-Smad, Medea, and the complex translocates to the nucleus to regulate transcription. 17 This signaling promotes synaptic stability 18 and increases the amount of glutamate released as the NMJ grows. 19 Gbb is also released from presynaptic motor neurons where it acts as an autocrine signal to facilitate homeostatic synaptic plasticity. 20
Synaptic signaling pathways are governed by developmental- and tissue-specific gene expression, which requires chromatin remodeling. Chromatin remodeling enzymes, including those of the chomodomain helicase DNA binding (CHD) family, are critical for neurodevelopment 21 and regulate the expression of genes required for cell adhesion, neurotransmission, and synaptic plasticity.22 -25 There are 9 CHD proteins, which are divided into 3 subfamilies. Both CHD7 26 and CHD8, 27 which are members of subfamily III, are important for early neurodevelopmental events including neural migration and differentiation. Although both proteins are expressed in human adult cortical neurons, 28 their roles in mature neurons are less well understood.
Kismet (Kis) is the sole Drosophila homolog of CHD subfamily III proteins including CHD7 and CHD8. 29 CHD8 enhances dendritic complexity in the adult mouse hippocampus 30 and spontaneous excitatory neurotransmission in the prefrontal cortex. 31 Loss of function mutations in Chd8 leads to impaired association, 32 spatial, 33 and contextual fear learning 34 in adult mice. Similarly, Kis promotes spontaneous neurotransmission at the Drosophila NMJ 35 and immediate recall memory in adult flies. 36 Kis also restricts the synaptic localization of cell adhesion molecules, including Fasciclin II, Integrins, and Neuroligins, 37 and facilitates both clathrin-mediated and activity-dependent bulk endocytosis. 38
CHD proteins and BMP signaling may influence one another but whether this relationship occurs in mature neurons has not been examined. CHD7 interacts with SMAD1/5/8 and these complexes are found bound to enhancers in response to BMP signaling in mouse embryonic cardiac cells. 39 CHD-7 is downstream of the DAF-1 receptor and R-Smad DAF14 and transcriptionally regulates the BMP components dbl-1 and sma-2 in C. elegans. 40 Both BMP signaling 41 and CHD7 transcriptional activity 26 are required for migration of neural crest cells. Chd8 is considered a high confidence risk gene for autism spectrum disorders (ASDs), 42 which are characterized partly by aberrant BMP/TGF-β signaling. 43 Finally, Kis binds near coding and/or regulatory regions of mad, tkv, and wit in Drosophila intestinal stem cells 44 and loss of function mutations in Chd7 lead to reduced bmp5 transcripts. 45 Therefore, we investigated whether Kis influences BMP signaling at the Drosophila third instar NMJ. We found increased synaptic pMad despite unchanged transcript levels of several BMP signaling components in kis mutants. There is no change, however, in pMad in motor neuron nuclei in kis mutants. These data, coupled with our finding that Kis is required in motor neurons for appropriate levels of synaptic pMad, indicates that defective BMP receptor trafficking is likely responsible for the accumulation of synaptic pMad in kis mutants.
Materials and Methods
Fly Husbandry and Stocks
Fly stocks were maintained at 25°C in a Percival DR-36NL incubator with a 12:12 h light dark cycle. Flies were fed Jazz-Mix Drosophila food (Fisher Scientific AS153). w 1118 (BL 3605), 24B-Gal4 (BL 1767), Actin5c-Gal4 (BL 9431), D42-Gal4 (BL 8816), elav-Gal4 (BL 8760), E(z) G4251 (BL 27149), gbb G12649 (BL 28388), kis k13416 (BL 10442), miR-965 K01 (BL 58946), nwk 1 (BL 51626), Pc KG05398 (BL 14002), tkv 1 (BL 427), UAS-tkv CA (BL 36537), and wit A12 (BL 5173) stocks were obtained from the Bloomington Drosophila Stock Center. UAS-kis RNAi.b flies were obtained from the Vienna Drosophila RNAi Center (#46685) while the kis LM27 protein null and UAS-kisL stocks were obtained from the laboratory of Dr. Daniel Marenda. 46 UAS-CHD7 and UAS-kis K2060R were generated as described in Hendricks and Liebl. 38
Male and female larvae were used for all experiments. Outcrossed controls for tissue-specific expression experiments were generated by crossing flies containing the UAS or Gal4 transgenes with w 1118 . w 1118 was used as a control for all other experiments. Because kis LM27 is homozygous lethal, these flies were crossed with kis k13416 to obtain kis LM27 /kis k13416 .
Dissections and Immunocytochemistry
Drosophila third-instar larvae were fillet dissected in Rodger’s Ringer (135 mM NaCl, 5 mM KCl, 4 mM MgCl2-6H2O, 1.8 mM CaCl2 2H2O, 5 mM TES, 72 mM sucrose, and 2 mM glutamate) on Sylgard (World Precision Instruments 501986)-coated 60 mm dishes. Larvae were fixed for 30 min with 3.7% paraformaldehyde (Fisher Scientific BP531-500) or Bouin’s fixative (Fischer Scientific 112016). The latter fixative was only used for Bruchpilot (Brp) and GluRIIC immunolabeling experiments. After fixation, larvae were washed 3 times for 10 minutes each in PTX (1 × PBS + 0.1% Triton, Fisher Scientific AAA16046AP) and 2 times for 30 minutes each in PBTX (PBS + 0.1% Triton + 1% Bovine Serum Albumin, Fisher Scientific BP1600-100). Primary antibodies were diluted in PBTX and incubated overnight at 4°C. Several primary antibodies were used including mouse α-Brp (1:50, Developmental Studies Hybridoma Bank [DSHB] nc82), rabbit α-Endophilin B (1:200, Li-Mei Pai lab), rabbit α-GluRIIC (1:5000, generated by Genscript using the sequence described in Marrus et al 47 ), rabbit α-Nwk (1:1000, Kate O-Connor Giles lab), rabbit α-pMad (1:100, Abcam ab52903), mouse α-SCAR (1:50, DSHB P1C1), and mouse α-WASp (1:10, DSHB P3B1). Larvae were then washed 3 times for 10 minutes each in PBTX followed by 2 times in PBTX for 30 minutes. Species-specific secondary antibodies were applied along with horseradish peroxidase (HRP) and incubated at room temperature for 2 hours. All secondary antibodies (1:400) and HRP (1:125) were obtained from Jackson Immunoresearch Laboratories and included α-mouse FITC (113-095-003), α-mouse TRITC (115-025-003), α-rabbit FITC (106-095-003), Cy3-HRP (123-165-021), and A647-HRP (123-605-021). Larvae were next washed 3 times for 10 minutes each in PBTX followed by 2 times in PBTX for 30 minutes. Larvae were subsequently mounted on slides using Vectashield Mounting Media (Vector Laboratories H1000).
Larvae were imaged using an Olympus Fluoview 1000 confocal microscope. Images of A3 or A4 6/7 NMJs were obtained using the 60× oil immersion objective. Images of ventral nerve cords were obtained using the 40× oil immersion objective. Controls were used to set imaging parameters for all experimental animals. Approximately equal numbers of controls and experimental animals were imaged each day. All immunocytochemistry experiments included at least 12 larvae and at least 2 biological replicates. The number of larvae used for each experiment are represented as data points in figure bar graphs.
RNA Isolation and RT-qPCR
Third instar larval CNSs or muscle pelts were dissected in Rodger’s Ringer and placed in RNAlater (Fisher Scientific AM7020) in nuclease-free 1.5 mL centrifuge tubes. Both male and female larvae were used for dissections. Dissected tissues were stored at −20°C and RNA was isolated using the Invitrogen Purelink RNA Mini Kit (Fisher Scientific 12-183-025). RNA concentrations were determined using an Implen Nanophotometer N50. Each biological replicate included RNA isolated from 30 third instar larval CNSs or 8 muscle pelts.
RT-qPCR was performed using the iTaq Universal SYBR Green One Step Kit (Bio-Rad, 1725151) and a CFX Connect Real-Time PCR Detection System (Bio-Rad). Primers were designed around intron-exon borders using PerlPrimer (v. 1.1.21). Primers were validated by creating standard curves based on reactions with serial dilutions followed by melt curve analysis. 100 ng of RNA was added to each reaction. Three biological replicates, each of which included 3 technical replicates, were used to calculate 2−ΔΔ C(t) values. 48 The latter were determined by first calculating ΔC(t)s, the difference between the C(t) value of the target transcript reaction and the C(t) value for Glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Next, the difference between the control and kis mutant ΔC(t)s was calculated and log transformed. The data points in Figure 4C represent technical replicates.
Behavior
Third instar larvae of both sexes were selected from vials and briefly cleaned with a damp brush to remove any debris. Five larvae were collected and assayed during each 30 second recording interval. Larvae were then allowed to wander on a 1.6% agar plate for 30 seconds to acclimate to the crawling surface before being transferred to a backlit, 1.6% agar-coated behavior arena for overhead recording. The crawling behaviors of 5 larvae were recorded for 30 seconds at 29.97 frames per second using a Cannon EOS M50 camera and recording began only when all larvae were engaged in forward-directed motion. The first 899 frames of each recording were opened and analyzed in Fiji (NIH Image J) using the wrMTrck plugin (written by Jesper S. Pedersen). 49 Values for distance traveled, average velocity, maximum velocity, and body lengths per second were recorded for each animal assayed. The X and Y coordinates of larval crawling paths, representing the mean distances traveled for each genotype, were exported and plotted in Microsoft Excel to generate tracings. Representative path tracings were overlayed to accommodate all tracings and do not reflect larval interactions or path crossings. At least 25 animals were used for all behavioral experiments.
Experimental Design and Statistical Analyses
Post-acquisition image processing was performed in Fiji (NIH Image J) 50 by converting z-stack slices to max intensity projection images and generating a binary projection for a NMJ region of interest (ROI) selection. Max projection images of each channel were analyzed to obtain the mean fluorescence intensity values of synaptic and background ROI’s. For each image analyzed, background fluorescence intensity was subtracted from synaptic fluorescence intensity to obtain the reported mean relative fluorescence intensity value. All reported values were normalized to controls. Colocalization of Brp and GluRIIC was determined by obtaining red-green intensity profiles from max z-projection images using Fiji. Lines were drawn on boutons perpendicular to the NMJ branch and the distance between the maximum peak for Brp and GluRIIC was obtained. Maximum peak distances for 5 terminal boutons per NMJ were averaged for each larva.
Sample sizes for each genotype in an experiment are indicated by data points on the bar graphs. Homogeneity of variance was assessed using Bartlett’s Test. Unpaired t-tests were used when comparing 1 experimental group to 1 control group. When more than 1 control group was required, experimental groups were compared to their respective controls through one-way ANOVA with post hoc Tukey’s test and P-values were corrected to account for multiple comparisons. All statistical tests were performed with GraphPad Prism (v. 10.0.1). Statistical significance is indicated on the graphs where *P < .05, **P < .001, and ***P < .0001. All error bars represent standard error of the mean (SEM).
Results
Synaptic pMad is Increased in kis Mutants But May Not Contribute to Some kis Mutant Phenotypes
Kis restricts the synaptic levels of several cell adhesion molecules including integrins, neuroligins, and Fasciclin II. 37 These data, coupled with the capacity of Kis to facilitate endocytosis, 38 could be partly mediated by retrograde synaptic signaling mechanisms. We first examined this possibility by assessing the synaptic levels of pMad in kis k13416 mutants, which are adult viable hypomorphs, and kis LM27 /kis k13416 mutants, which are third instar lethal due to the null kis LM27 allele. 46 Both kis mutants exhibited an increase in synaptic pMad (Figure 1A). Therefore, we sought to determine whether increased BMP signaling could mimic kis mutant phenotypes. To enhance BMP signaling, we expressed a constitutively active Tkv, TkvCA, in motor neurons using the D42-Gal4 driver. Expression of TkvCA in presynaptic motor neurons induces expansion of the NMJ and increases the number of satellite boutons, 51 both of which are observed in kis mutants.35,52 We found a decrease in Endophilin B (EndoB), a BAR-domain-containing protein required for endocytosis in Drosophila oocytes, 53 when TkvCA was expressed in motor neurons compared with the driver-specific outcrossed control but not the UAS-specific outcrossed control (Figure 1B). EndoB is also reduced in kis mutants as Kis transcriptionally regulates endoB. 52 These data suggest that the increase in synaptic pMad in kis mutants may influence the phenotypes observed at kis mutant synapses.

Kis limits synaptic pMad and constitutively active BMP signaling mimics the kis mutant phenotype. (A) High resolution confocal micrographs of terminal 6/7 NMJ boutons show presynaptic motor neurons (HRP, magenta) and pMad (green). Quantification of pMad fluorescence in genotypes listed. *P = .02 and **P = .0014. (B) Confocal images of terminal 6/7 NMJ boutons show presynaptic motor neurons (magenta) and EndoB (green). A constitutively active BMP receptor (TkvCA) was expressed in motor neurons using the D42-Gal4 driver. Quantification of EndoB in genotypes listed. *P = .011 and ***P = .0008; Scale bars = 5 µm.
Kis promotes larval locomotion and the appropriate apposition of postsynaptic glutamate receptors to presynaptic active zones. 35 If Kis restricts synaptic pMad accumulation to ensure proper locomotion and apposition, we would expect to observe the opposite phenotypes in gbb, tkv, and wit loss of function mutants as that of kis mutants. Contrary to this and similar as kis mutants, 35 gbb G12649 and tkv 1 mutants showed a decrease in the glutamate receptor subunit, GluRIIC, at the synapse (Figure 2A and B) compared with controls. Also similar as kis mutants, 35 gbb G12649 and wit A12 mutants showed an increase in the distance between the active zone protein, Bruchpilot (Brp), and GluRIIC compared with controls (Figure 2A and B). Synaptic Brp levels, which are unaffected in kis mutants, 35 were decreased in both tkv 1 and wit A12 mutants compared with controls (Figure 2A and B). Thus, Kis may regulate synaptic pMad and the levels and localization of Brp and GluRIIC independent of BMP signaling.
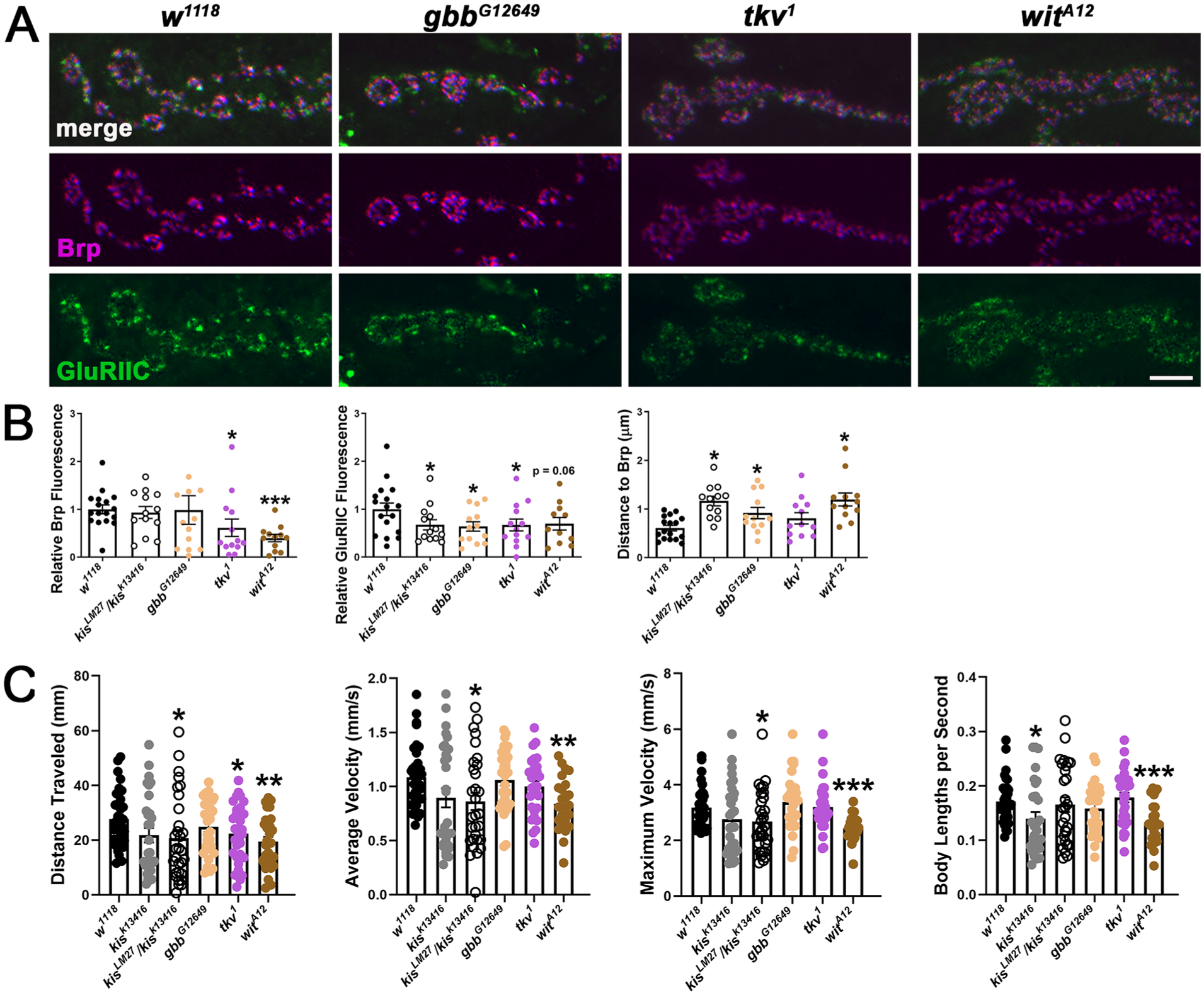
Mutations that diminish BMP signaling partially phenocopy kis mutants. (A) High resolution confocal micrographs of terminal 6/7 NMJ boutons show presynaptic Brp (magenta) and the glutamate receptor subunit, GluRIIC (green). Scale bar = 5 µm (B) Quantification of Brp and GluRIIC fluorescence and the distance of GluRIIC to Brp in genotypes listed. (C) Larval crawling behavior for 30 seconds quantified by wrMTrck. Histograms show total distance traveled, average velocity, maximum velocity, and body lengths per second.
We next examined broader conserved functions at the synapse in the regulation of larval motor function. Larval crawling is a relatively complex behavior as it requires the proper engagement and synchrony of system-wide excitatory and inhibitory motor circuits. 54 As such, quantification of larval locomotion can provide general insights into overall synaptic health and circuit function. Kis is required in either presynaptic motor neurons or postsynaptic muscle for proper motor function. 38 We found that both kis LM27 /kis k13416 and wit A12 mutants exhibited impaired locomotion as evidenced by decreased distance traveled and average and maximum velocities compared with controls (Figure 2C). Although tkv 1 mutants showed reduced distance traveled compared with controls, average and maximum velocities were unaffected. Gbb G12649 mutants did not exhibit impaired locomotion. These data indicate that Kis may independently regulate locomotion and synaptic pMad levels.
To ensure the increase in synaptic pMad did not occur due to a background mutation in kis mutants, we knocked down Kis in all tissues using the Actin5c-Gal4 driver. This produced a similar increase in synaptic pMad as observed in kis LM27 /kis k13416 mutants compared with outcrossed controls (Figure 3A and B). We further confirmed that Kis was responsible for the increase in pMad by examining synaptic pMad in miR-965 knock outs (miR-965 KO1 ). The first intron of kis includes the microRNA, miR-965, 55 raising the possibility that the increase in synaptic pMad may occur in kis mutants because of the loss of miR-965. Synaptic pMad is unchanged in miR-965 KO1 mutants compared with controls (Figure 3C and D). These data indicate that Kis may restrict BMP signaling at the synapse but the increase in pMad in kis mutants is independent of most kis mutant phenotypes examined.

Synaptic pMad is increased in Kis KD but unchanged in miR-965 knock outs. (A) and (D) High resolution confocal micrographs of terminal 6/7 NMJ boutons show presynaptic motor neurons (HRP, magenta) and pMad (green). Scale bar = 5 µm. (B) and (C) Quantification of pMad fluorescence in genotypes listed.
The Increase in Synaptic pMad in kis Mutants May be Independent of Nwk and Transcription of BMP Signaling Components
We next sought to identify the mechanism by which Kis influences pMad. Kis was first identified in a suppressor screen for Polycomb Group (PcG) mutant phenotypes. 56 PcG proteins form polycomb repressive complexes (PRCs), which regulate transcription 57 and are antagonized by Trithorax Group proteins, including Kis. 58 There are 2 PRC complexes, PRC1 and PRC2, which include the chromodomain protein, Polycomb (Pc), and the methyltransferase, E(z), respectively. 59 Therefore, we examined synaptic pMad in larvae with loss of function mutations in E(z), E(z) G4251 ,and in Pc, Pc KG05398 . 60 In contrast to kis LM27 /kis k13416 mutants, E(z) G4251 mutants showed reduced synaptic pMad (Figure 4A and B) suggesting that Kis may repress E(z)/PRC2 activity to influence synaptic pMad.

Kis does not influence transcript levels of BMP signaling components but promotes nuclear VNC pMad localization. (A) Confocal images of terminal 6/7 NMJ boutons showing the presynaptic motor neuron (HRP, magenta) and pMad (green). Scale bar = 5 µm. (B) Quantification of presynaptic BMP in genotypes listed. (C) Expression of transcripts that encode BMP signaling components in kis mutants is shown relative to the control, w 1118 . Relative expression was assessed via RT-qPCR and 2−ΔΔC(t) values are shown. Data points represent technical replicates. (D) Confocal images of VNCs showing presynaptic motor neurons (HRP, magenta) and nuclear pMad. Scale bar = 50 µm. Right, quantification of nuclear pMad levels.
Kis binds near or within regulatory regions of mad, tkv, and wit in Drosophila intestinal stem cells. 44 If Kis represses E(z)/PRC2 activity to regulate synaptic pMad levels, we might observe increased transcript levels of BMP signaling components in kis mutants. In addition to gbb, mad, tkv, and wit, we examined babo and daw, which encode other NMJ TGF-β signaling components, 61 and twit, which encodes a glycosylphosphatidylinositol (GPI)-anchored protein expressed in motor neurons via Wit-mediated transcription. 62 None of the transcripts examined in either the CNS or postsynaptic muscle showed differential expression in kis k13416 or kis LM27 /kis k13416 mutants compared with controls (Figure 4C). Thus, Kis may not influence the transcription genes encoding these BMP signaling components.
Retrograde trafficking of R-Smad/Co-Smad complexes is required for BMP signaling transcriptional regulation. 17 Given that there is no increase in twit in the CNS of kis mutants and Kis promotes several vesicle trafficking steps in neurons,52,63 we examined nuclear levels of pMad in presynaptic motor neurons. Presynaptic motor neuron cell bodies are contained within the ventral nerve cord (VNC), which is part of the Drosophila third instar larva CNS. Although pMad is not trafficked from synapses to VNC nuclei, endocytosed dimerized BMP receptors are retrogradely trafficked in dynein-dependent manner to the VNC nuclei. Mad is subsequently phosphorylated to serve as a transcription factor. 64 Mutations that produce increased synaptic pMad including liquid facets, 65 spartin, 51 and abelson interactor 66 also show increased pMad in presynaptic motor neuron nuclei. However, we found no differences in kis mutant presynaptic motor neuron nuclear pMad levels compared with controls (Figure 4D) suggesting that Kis may promote retrograde trafficking of BMP receptors.
We next sought to determine whether kis mutants possess an impaired capacity to endocytose BMP receptors thereby leading to the lack of increased pMad in VNC nuclei. Endocytosis of dimerized BMP receptors is mediated by a direct interaction between Tkv and Nervous Wreck (Nwk), which also interacts with the endocytic proteins Dynamin and Dap160.67,68 Nwk influences endocytosis partly by stimulating the activity of Wiskott-Aldrich syndrome protein (WASp), 68 which activates Arp2/3 to enable the branching of actin filaments. 69 SCAR also activates Arp2/3 and loss of function mutations in the SCAR component, abelson interactor, results in the accumulation of synaptic pMad. 66 We did not observe changes in the synaptic levels of Nwk, SCAR, or WASp in kis mutants compared with controls (Figure 5). These data suggest that Kis may not influence trafficking of BMP receptors by Nwk-dependent mechanisms.

Synaptic levels of Nwk, SCAR, and WASp are unchanged in kis mutants. Confocal images of terminal 6/7 NMJ boutons showing the presynaptic motor neuron (magenta) and Nwk (A), SCAR (B), or WASp (C). Scale bars = 5 µm. Right histograms show quantification of relative fluorescence intensities in controls and kis mutants.
Kis and Its ATPase Domain are Required in Neurons for Appropriate Synaptic pMad Levels
Kis is expressed ubiquitously early in development but later is progressively restricted to specific tissues including the brain and VNC. 70 To determine the tissue-specific requirements of Kis for appropriate synaptic levels of pMad, we restored kis expression in kis mutants by expressing the long isoform of kis, kisL, in all tissues using the Actin5c-Gal4 driver, in neurons using the elav-Gal4 driver, or in postsynaptic muscles using the 24B-Gal4 driver. Expression of kisL in all tissues or in neurons rescued synaptic pMad levels compared with outcrossed controls. Expression of kisL in postsynaptic muscle cells, however, produced an increase in synaptic pMad levels similar as that observed in kis LM27 /kis k13416 mutants (Figure 6A and B). These data indicate that Kis is required in presynaptic motor neurons but not postsynaptic muscles for appropriate BMP signaling.

Kis, including its ATPase activity, is required in neurons for proper synaptic pMad levels. (A, C, and E) High resolution confocal micrographs of terminal 6/7 NMJ boutons showing the presynaptic motor neuron (HRP, magenta) and pMad (green). The UAS-kisL (A), UAS-kis K2060R (C), or UAS-CHD7 transgenes were expressed in all tissues using the Actin5c-Gal4 driver, in neurons using the elav-Gal4 driver, or in postsynaptic muscle using the 24B-Gal4 driver in kis mutants. Scale bars = 5 µm. (B, D, and F) Quantification of pMad fluorescence intensities in genotypes listed.
The ATPase activity of CHD proteins is required for their chromatin remodeling activity. 71 If Kis remodels chromatin to enable appropriate synaptic pMad levels, then we would expect that expression of an ATPase deficient Kis would fail to rescue synaptic pMad. Mutation of a Lys residue within the conserved ATPase domain 72 of CHD8 renders it incapable of hydrolyzing chromatin and remodeling chromatin. 73 Therefore, we mutated the Lys residue within the Kis ATP binding site to an Arg (KisK2060R) and confirmed that KisK2060R was expressed in the nuclei of both presynaptic motor neurons or postsynaptic muscles when expressed with the elav-Gal4 or 24B-Gal4 drivers, respectively. 38 Surprisingly, expression of kis K2060R in all tissues using the Actin5c-Gal4 driver rescued synaptic pMad levels (Figure 6C and D). In contrast, neuronal expression of kis K2060 failed to rescue the accumulation of synaptic pMad. Expression of kis K2060 in either neurons or postsynaptic muscle cells resulted in an increase in synaptic pMad compared to outcrossed controls similar as the increase observed in kis LM27 /kis k13416 mutants (Figure 6C and D). Collectively, these data indicate that Kis, including its chromatin remodeling activity, is required in neurons for appropriate synaptic levels of pMad. The chromatin remodeling activity of Kis, however, is not required in all tissues for appropriate synaptic levels of pMad.
Kis is 63% identical to human CHD7 with increased conservation in its major functional domains. 29 CHD7 regulates the expression of genes required for cell adhesion, neurotransmission, and synaptic plasticity. 23 Given their similar structure and function, we examined whether expression of human CHD7 could rescue synaptic pMad levels in kis mutants. We constructed a human CHD7 codon-optimized for Drosophila and verified that it was localized to presynaptic motor neuron or postsynaptic muscle nuclei when expressed under the control of the elav-Gal4 or 24B-Gal4 drivers, respectively. 38 Synaptic pMad levels were rescued by UAS-CHD7 expression in all tissues using the Actin5c-Gal4 driver in kis LM27 /kis k13416 mutants compared with outcrossed controls (Figure 6E and F). Expression of CHD7 in either neurons or postsynaptic muscles of kis LM27 /kis k13416 mutants, however, failed to rescue synaptic pMad (Figure 6E and F). Thus, CHD7 is required in both neurons and postsynaptic muscle to functionally compensate for the loss of Kis but cannot compensate when expressed in neurons or muscle alone.
Discussion
CHD proteins are required for neurodevelopmental events including migration of neural crest cells 26 and cell cycle exit of neural progenitor cells. 32 Both CHD7 and CHD8 are expressed in mature neurons 28 but how they influence the activity of mature neurons is not well understood. CHD7 is downstream of BMP signaling in both mouse embryonic cardiomyocytes 39 and in C. elegans where CHD-7 regulates the transcription of the BMP components dbl-1 and sma-2. 40 Similarly, Kis may regulate mad, tkv, and wit transcription in Drosophila intestinal stem cells. 44 Collectively, these data suggest a relationship between CHD7 and BMP signaling but this relationship has not been explored. We found that Kis limits the accumulation of pMad at the synapse (Figure 1A) and facilitates the localization of pMad to VNC motor neuron nuclei (Figure 4D). Our data suggest a model where Kis regulates BMP receptor trafficking via a transcription-dependent mechanism (Figure 7).

Kis restricts synaptic pMad and may promote retrograde trafficking of BMP receptors. In wild type (WT) neurons, binding of ligand to BMP receptors induces phosphorylation of Mad (pMad). Ligand-bound receptors are also endocytosed and trafficked to the nucleus where pMad induces transcription of BMP target genes including trio and twit. Kis mutants show increased synaptic pMad but reduced nuclear pMad indicating that retrograde trafficking of BMP receptors is likely impaired.
Kis most likely limits synaptic and promotes nuclear pMad localization by influencing BMP receptor trafficking. BMP receptors are endocytosed from the plasma membrane by interactions with Nwk, 67 which links receptor complexes to the endocytic machinery and actin regulatory proteins like WASp and SCAR. 74 Although kis mutants do not exhibit altered levels of Nwk, WASp, or SCAR (Figure 5), endocytosis is compromised in kis mutants. 38 Impaired endocytosis would enhance BMP signaling by increasing the number of and/or length of time receptors are localized to the synaptic plasma membrane. Indeed, loss of function mutations in genes that encode proteins involved in BMP receptor endocytosis including nwk, endoA, 67 epsin1/liquid facets, 65 spartin, 51 and abelson interactor 66 lead to increased synaptic pMad. Once endocytosed, activated BMP receptors are directed to endosomal populations including early endosomes and multi-vesicular bodies, where BMP receptor signaling is amplified, and late endosomes/lysosomes. In addition, some BMP receptors are trafficked to the nucleus 15 where Mad is phosphorylated. 64 Notably, liquid facets, 65 spartin, 51 and abelson interactor 66 mutants show increased pMad both at the synapse and in the presynaptic motor neuron nuclei. kis mutants, however, showed no change in VNC nuclear pMad levels compared with controls (Figure 4D) indicating that retrograde trafficking of activated BMP receptors may be impaired in kis mutants.
Kis may transcriptionally promote the expression of gene products that enable appropriate BMP receptor trafficking in presynaptic motor neurons. In support of this possibility, expression of the long isoform of Kis in kis mutant neurons rescues synaptic pMad levels (Figure 6A) and Kis’ chromatin modifying/ATPase activity is required for rescue (Figure 6B). Additional work is required to identify specific, Kis regulated gene products that promote BMP receptor trafficking. Candidates include Rabs, which are GTPases that mediate directional membrane trafficking between specific endomembrane system compartments. 75 Kis binds to predicted regulatory regions of several genes that encode Rabs. 44 Kis also promotes rab11 transcription in Drosophila third instar neurons and enhances Rab11 at the synapse. 52 Other potential candidates include lipid regulatory enzyme gene products as subpopulations of specific lipids assist in membrane trafficking by enabling membrane recruitment and/or directly regulating membrane dynamics. 76 Lipid binding gene products were downregulated in Chd7 knock out spinal cord tissues. 77
It is also possible that Kis may regulate BMP signaling by indirectly influencing the expression of BMP signaling components. CHD proteins are recruited to genomic loci by specific chromatin modifications. For example, CHD7, 78 CHD8, 79 and Kis 44 are associated with posttranslational methylation of the histone 3 Lysine 4 (H3K4) residue. Kis also promotes multiple histone modifications including H3K27 acetylation, H3K36 methylation, 80 and H4K16 acetylation. 36 These data suggest a model whereby CHD proteins like Kis are recruited to enhancers and/or other regulatory elements by specific chromatin modifications and regulate transcription by subsequently recruiting additional chromatin modifiers. Consistent with this, Kis antagonizes PcG activity at H3K36 in Drosophila polytene chromosomes by recruiting the methyltransferases Ash1 and Trithorax to chromatin thereby promoting methylation of H3K36, 80 which physically interferes with PcG binding. Our data suggest that the functional antagonism between Kis and PRC activity may influence synaptic pMad as Kis restricts while PRC2 enhances pMad (Figure 4A and B). This functional antagonism may or may not be dependent on the transcriptional activity of Kis and PRCs given that Kis did not affect the expression of genes encoding BMP signaling components (Figure 4C). The kis mutants we used are not homozygous null mutants as the latter are embryonic lethal. Thus, Kis may be required for the expression of some BMP responsive gene targets. Both CHD7 39 and PRC-dependent modifications 81 are found at SMAD binding/response elements as directed by BMP signaling. It is also possible that Kis suppresses BMP transcriptional activity both by limiting nuclear pMad and by interfering with the derepression of BMP signaling by PRCs. Mel18, a PRC1 subunit, represses transcription of negative regulators of BMP signaling in mouse embryonic stem cells 82 thereby enabling BMP signaling. Similarly, inhibition of the PRC2 subunit, Ezh2/E(z), results in demethylation of H3K27 and activation of BMP signaling as evidenced by SMAD phosphorylation in mouse osteoblast cell lines. 83 Thus, the loss of Kis could maintain the transcriptional repression of negative regulators of BMP signaling by PRCs.
Our data also suggest that Kis regulates synaptic pMad levels independent of its chromatin remodeling activity. Synaptic pMad was rescued either by expression of KisL in all tissues (Figure 6A) or expression of an ATPase-deficient Kis, Kis K2060R , in all tissues (Figure 6B). CHD protein ATPase activity is required to reposition histones relative to DNA. 84 We previously showed that Kis is required in postsynaptic muscles for endocytosis and expression of KisK2060R in kis mutant muscle cells restored endocytosis. Further, expression of either Kis or KisK2060R in kis mutant presynaptic neurons or muscles restores locomotor behaviors. 38 The size of stem cell clones in kis 10D26 loss of function mutants is also partially rescued by KisK2060R expression. 44 The activity of chromatin remodeling proteins is not limited to their capacity to reposition nucleosomes. CHD7 contributes to cardiac neural crest cell development both by remodeling nucleosomes and by recruitment of histone modifying enzymes, including the histone methyltransferase subunit, WDR5, to genomic regulatory regions. 85 CHD proteins are also important because they are critical organizers of large, multisubunit complexes, which vary over developmental time in a tissue-specific manner.86,87 Although our data provide additional insight into the function of CHD complexes in mature neurons, additional work will be required to elucidate the mechanisms by which BMP signaling is specifically regulated.
Footnotes
Acknowledgements
We thank Daniel Marenda for generously providing the UAS-kisL and kis LM27 fly stocks and the Bloomington Drosophila Stock Center for additional fly stocks. We thank the Li-Mei Pai lab for the EndoB antibody and the Developmental Studies Hybridoma Bank (University of Iowa) for additional antibodies. We thank Ireland Smith and Nadine Assi for assistance with data analyses. Finally, we thank Dave Featherstone for his guidance and mentorship.
Ethical Considerations
This article does not contain any studies with human or animal participants.
Author Contributions
RAK, ELH, and FLWL were responsible for conceptualization and writing. RAK, JAP, ELH, and FLWL were responsible for performing experiments and data analyses.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institutes of Health, NINDS 2R15NS101608-02A1 (FL), and Southern Illinois University Edwardsville’s Competitive Graduate Award (RK and EH). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.