
Abstract
Imperata cylindrica is a globally distributed plant known for its antiepileptic attributes, but there is a scarcity of robust evidence for its efficacy. The study investigated neuroprotective attributes of Imperata cylindrica root extract on neuropathological features of epilepsy in a Drosophila melanogaster mutant model of epilepsy. It was conducted on 10-day-old (at the initiation of study) male post-eclosion bang-senseless paralytic Drosophila (parabss1) involved acute (1-3 h) and chronic (6-18 days) experiments; n = 50 flies per group (convulsions tests); n = 100 flies per group (learning/memory tests and histological examination). Administrations were done in 1 g standard fly food, per os. The mutant flies of study (parabss1) showed marked age-dependent progressive brain neurodegeneration and axonal degeneration, significant (P < 0.05) bang sensitivity and convulsions, and cognitive deficits due to up-regulation of the paralytic gene in our mutants. The neuropathological findings were significantly (P < 0.05) alleviated in dose and duration-dependent fashions to near normal/normal after acute and chronic treatment with extract similar to sodium valproate. Therefore, para is expressed in neurons of brain tissues in our mutant flies to bring about epilepsy phenotypes and behaviors of the current juvenile and old-adult mutant D. melanogaster models of epilepsy. The herb exerts neuroprotection by anticonvulsant and antiepileptogenic mechanisms in mutant D. melanogaster due to plant flavonoids, polyphenols, and chromones (
Keywords
Introduction
Epilepsy is a chronic noncommunicable brain (neurological) disorder associated with excessive neuronal activity and unprovoked seizures referred to as ‘epileptic seizures’ and sensory-motor deficits that cause neurobiological, cognitive, psychological, and social consequences to the patient.1-4 It is one of the most common neurological disorders worldwide,1-5 is among the most common chronic brain diseases, and affects people of all ages. Around 50 million people or more worldwide have epilepsy, with almost 80% of them living in low- and middle-income countries.2-4 The global prevalence of active epilepsy is 0.64%, with a lifetime prevalence of 0.76% and an incidence of 61.4 per 100,000 person-years. The prevalence of the disease increases with age and is higher in people who are socially deprived. 6 Africa is estimated to contribute about 20–23% of the global cases, and the prevalence of epilepsy in Uganda is estimated to be ranging between 0.22 and 13 per 1000 population, with an estimated incidence of 156 cases per 100,000 each year.7,8
Genetic epilepsy arises from mutations in one or multiple genes and can occur in non-ion and ion channel genes. 9 Of huge significance are the voltage-gated sodium ion channels (VGSCs) which are transporters of ions facilitating the depolarization of action potentials and transmission of signals in neurons. The most common types of epilepsy occurring in humans are the genetic forms due to genetic disorder (s) in the VGSCs of the brain (brain sodium channelopathies).9-11 Studies involving animal models with genetic forms of epilepsy due to brain sodium channelopathies are vital for a better understanding of the pathophysiology of neuropathological features of epilepsy associated with these channelopathies. This knowledge can be utilized to develop newer effective therapies for the genetic epilepsies that arise due to channelopathies. 11
Drosophila melanogaster is a good model for studying neurological disease pathways and for utilization during drug discovery for neurological diseases including epilepsy mainly because the flies and humans have similar basic biology, physiology, and neurological properties, with 75% of disease-causing human genes being conserved in the flies. 12 In D. melanogaster flies the VGSCs are mainly coded by a single gene called the paralytic (para) gene.13,14 A mutation in the para gene (overexpression of para) in the neuronal brain tissues of bang-senseless (bss) mutant epileptic Drosophila models (parabss) gives rise to epileptic neuropathological features in the flies that are similar to those in epileptic mammals. This mutation induces a reduction in seizure stimulation threshold in parabss which makes the flies more susceptible to getting seizures following electrical, mechanical, or physical stimulation of the animals.15,16 In addition, gene mutations or alterations due to the para gene in D. melanogaster fly models are associated with defective metabolism and viability in neurons of these flies leading to defective neurophysiology and behavior.17,18 Again, the gene defects are associated with extensive neurodegeneration in the brain of the flies 17 as well as an impairment of sensory-motor activities, and neurobiological, and cognitive deficits in the animals.19,20 Because almost 67% of human disease genes are conserved in Drosophila, 21 the defects and deficits observed in the epileptic fly models (referred to as ‘para neuropathology’) are related to the ‘epileptic syndrome’ in people with genetic epilepsy that is characterized by brain sclerosis, convulsions, cognitive deficits, motor deficits, etc.21-25
Some epileptic patients (almost a third of them) do not get the expected relief from seizures and other neuropathological features of epilepsy after administration with the current anti-epileptic treatments,10,26 also, the anti-epileptic drugs are associated with numerous severe side-effects causing discomfort, drug non-compliance, and drug resistance in the patients.16,27,28 Other challenges faced by epileptic patients include the inadequate number of neurologists and the lack of good healthcare facilities for epilepsy patients in developing countries.29,30 Therefore, herbal extracts are an important source for the development of novel complementary and alternative antiepileptic medicines both in developing and developed countries.30-33
Imperata cylindrica (L.) P. Beauv (Poaceae) is native to tropical regions of Africa, tropical and subtropical zones, southwestern Asia, and some parts of the U.S.A,34,35 is an important neuromedicinal plant in Africa, Asia, and elsewhere, and could be a good source for the development and discovery of novel complementary and alternative antiepileptic medicines globally due to its widespread utilization in treating epilepsy in people with epilepsy.34-36 Phytochemicals from the herb such as the 2-(2-Phenylethyl) and 5-Hydroxy-2-(2-phenylethyl) chromones have been found to possess neuroprotective potential on glutamate-induced neurotoxicity in rat cerebral cortical cells,37,38 additionally plant-based nonenzymatic antioxidants mainly phenols, flavonoids, chromones, and glycosides provide neuroprotective attributes against brain cell damage through their strong antioxidant potentials that improve brain cell biology,37,39 provide anticonvulsant and antiepileptogenic attributes, 40 and alleviate cognitive deficits 41 of animals by the antioxidation mechanism which controls oxidative stress markers in the brain of animals.39-41
Although it is widely used, there is a lack of strong evidence for the efficacy and safety of Imperata cylindrica or I. cylindrica as an anticonvulsant in folk medicine in Africa, Asia etc,35,36,42 therefore, information on evidence-based scrutiny of antiepileptic/anticonvulsant efficacy and safety of this commonly used neuromedicinal (antiepileptic) herb is crucial for establishing targeted and effective community-based intervention policies and programs during the management of epilepsy.32,43,44
The study aimed to establish the neuroprotective effects of phytochemicals and changes in convulsion, cognitive, and histological parameters in brain tissues caused by Imperata cylindrica root extract on the neuropathological features of Drosophila melanogaster mutant models of epilepsy.
Materials and Methods
Plant Materials
Fresh plants of Imperata cylindrica were harvested in Ishaka-Bushenyi Municipality, Bushenyi District, western Uganda (GPS coordinates; Latitude: −0.53828 S 0°32′17.79583″; Longitude: 30.14546 E 30°8′43.64405″), and identified and authenticated by a botanist at the Department of botany, Mbarara University of Science and Technology (voucher No. P. Beauv /11/2020). The plants were cleaned and washed with distilled water to remove all foreign matter and the roots were collected and made into pieces and dried under shade. The methanol root extract was prepared using standard methods.45-47 Briefly, the dried roots were ground using a grinding machine into fine powders that were kept in air-tight glass containers. The methanolic extract of the plant was prepared by soaking 100 g of the powdered sample in 1000 mL of 70% methanol in an Erlenmeyer conical flask (Corning® CLS431145, Darmstadt, Germany). This was placed on an orbital shaker incubator (Amerex®, Gyromax 727, NY, USA) to allow complete mixing (24 h, 100 rpm, 30°C). The debris was removed by centrifugation (CYAN CL008N, USA; 4000 rpm, 15 min) to obtain a fine extract that was then filtered using Whatman number 1 filter papers (Whatman® qualitative filter paper, Grade 1, WHA1001325, Darmstadt, Germany). The extract was concentrated to dryness using a rotary evaporator (Büchi® Rotavapor® R-210, Büchi 23011V010, Darmstadt, Germany) at 53°C to evaporate off the methanol, followed by a concentration to dryness at 53°C using an electric oven (GallenKamp BS300, London, England). The remaining dry concentrated extract was weighed and stored in universal bottles put in a freezer at 4°C (Samsung RF22N9781SR, UPC: 887276259277, Brooklyn, NY, USA) till utilization.45-47
Animals
The study was conducted on the male adult or post-eclosion D. melanogaster flies at 10 days of age at the time of study initiation following previous studies on bang-senseless (bss) D. melanogaster models (parabss) and novel anti-epileptic treatments.15,16 The Drosophila models of the bang sensitive (BS) family used in the study were the bang senseless or bss1 mutants of the paralytic gene (parabss1),48,49 and the wild-type strains used as controls were Oregon-R or OrR and Canton-Special (CS).49-51 Male flies were used as a previous laboratory study has shown differences between female and male bss Drosophila because the parabss is located on the X chromosome, and heterozygous parabss/+ female Drosophila have significantly decreased recovery times. 52 Only animals that underwent seizures were included in the study during behavioral (convulsion and learning/memory tests) and histological assessments. 49
The bang-sensitive mutant flies (parabss1) obtained from Prof. Richard Baines laboratory (University of Manchester, UK)48,49 were used, and the wild-type flies ie, OrR (BDSC stock #25211) and CS (BDSC stock #64349), were obtained from Bloomington Drosophila Stock Center, Indiana University, U.S.A, and all the stocks were delivered to the Institute of Biomedical Research for culturing into sufficient stock colonies. The bss paralytic (para) D. melanogaster mutant flies used in the study have a gain-of-function mutation in the para sodium channel gene located at the para locus in neurons of the mutant flies.52-54
The flies were cultured on a standard yeast/cornmeal/agar diet (distilled water, 6.65% cornmeal, 7.15% dextrose, 5% yeast, 0.66% industrial agar supplemented with 3.4 mL/L propionic acid, and 2.2% nipagin) for Drosophila flies following previous methods,55,56 (Supplementary file 1). The flies were kept in incubators at 25°C, 65% humidity, and on a 12-h light/dark cycle (Fly incubator with programmable day/night cycle; Powers Scientific Inc., S33SD, USA). Flies were transferred to fresh plastic fly vials (Genesee Scientific, 32-116, USA), every 3 days, and fly density was kept to 10 flies per plastic fly vial (Genesee Scientific, 32-116, USA).
Experimental Design and Flies’ Treatment
The study was conducted for 1, 2, 3 h, and 6, 12, and 18 days for acute and chronic experiments respectively based on previous studies.52,57,58 The preliminary acute toxicity tests on D. melanogaster from our laboratory using a previous method, 59 found the lethal concentration 50 (LC50) of I. cylindrica P. Beauv methanol root extract (acute oral exposure for 24 h in 20-day-old post-eclosion Drosophila) to be 3 g/mL of extract in the medium, and doses below 2 g/mL produced no lethality. Mutant animals treated with the extract solution appeared healthy and showed reduced seizure-like behavior compared to the untreated mutant controls. Therefore, we decided to expand the assessments to lower extract concentrations (0.6, 0.8, and 1 g/mL) and (0.0125, 0.025, and 0.05 g/mL) in our acute and chronic experiments respectively. The doses of sodium valproate or SV (0.3 and 0.15 mg/mL, p.o) that were used in standard control groups of the study were based on previous studies that have shown them to provide significant anticonvulsant, anti-epileptic activities, and alleviative control of bss behaviour in epileptic D. melanogaster models eg parabss.15,60,61
The initial experiments assayed the bang-sensitivity and convulsion characteristics of our mutant flies, and also evaluated the extract doses that provide anticonvulsant potential on the mutant flies of the study following short-term and prolonged treatments. To do this, juvenile adult flies (10 days) were anesthetized using CO2 anesthesia (Fly CO2 anesthesia setup; Genesee Scientific, 59-114/54-104M, USA), sexing was performed on a CO2 perfused pad to separate flies into females and males (dissecting stereo microscope; Leica Microsystems, Leica M60 CMO), and the males were randomly assigned to plastic fly vials (Genesee Scientific, 32-116, USA), with 10 flies each. Male Animals were subjected to a vortex stimulus to induce seizures,62,63 seizing males were separated from non-seizing males and the former were then randomly assigned to the different groups (negative/mutant control, standard control, and low and high-dose extract tests groups), and the wild-type flies were used in the normal control groups, Table 1.
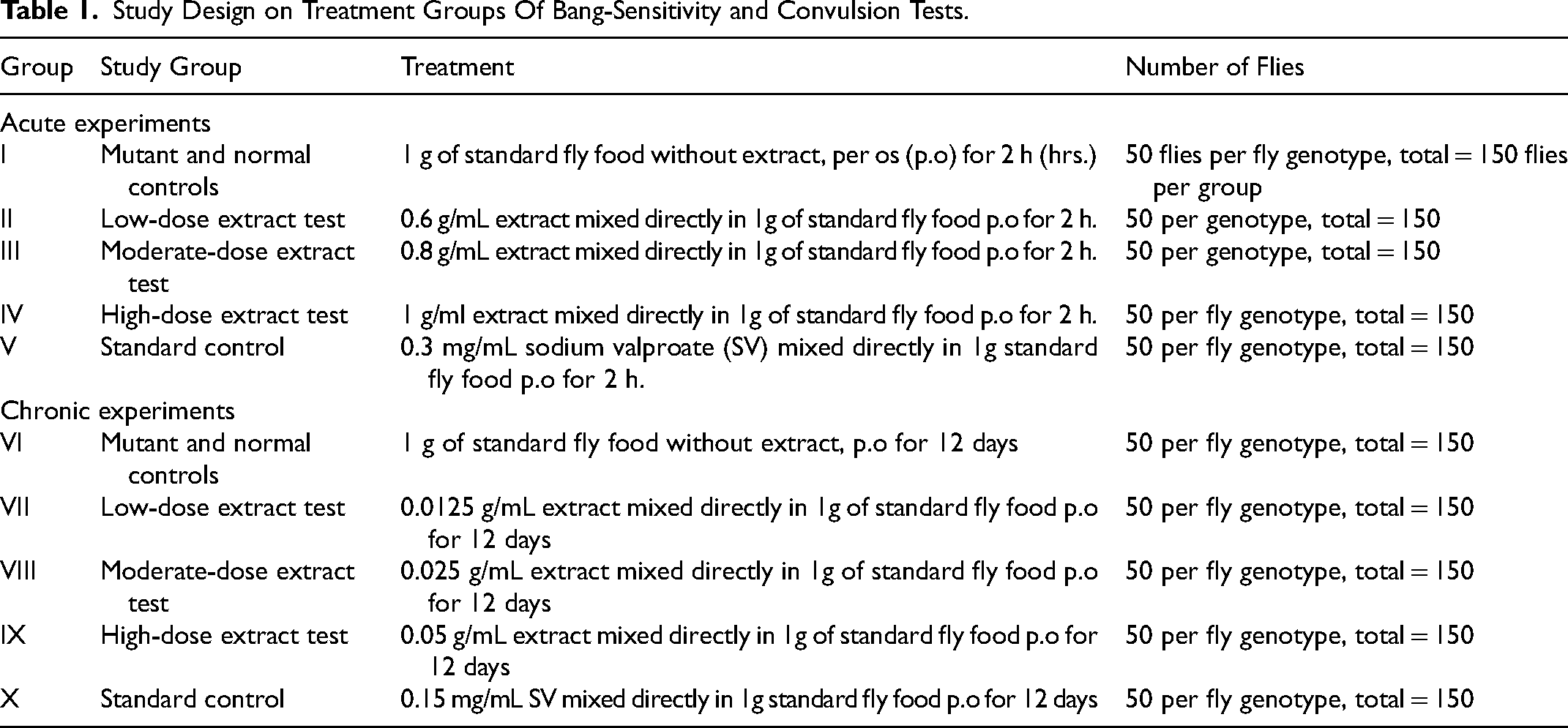
Study Design on Treatment Groups Of Bang-Sensitivity and Convulsion Tests.
Then, during the subsequent learning, memory, and histological tests, juvenile adult flies (10 days) were also anesthetized using CO2 anesthesia with Fly CO2 anesthesia setup (Genesee Scientific, 59-114/54-104M, USA), separated on a CO2 perfused pad into females and males (dissecting stereo microscope; Leica Microsystems, Leica M60 CMO), and the males were randomly assigned to plastic fly vials (Genesee Scientific, 32-116, USA), with 10 flies each. The flies were then tested for bang sensitivity (seizures) and the animals that underwent seizures were then randomly assigned to the different groups (negative/mutant control, standard control, and extract test groups), while wild-type flies were used in the normal control group, Table 2.
Study Design on Treatment Groups of Learning, Memory, and Histological Tests.

Drug Preparation and Administration
I. cylindrica root extract and sodium valproate were administered by mixing them with standard cornmeal fly food (Supplementary file 1), and extract and drug solutions were prepared fresh prior to each experiment. The experiment does are expressed in g of extract per mL of fly food and mg of sodium valproate per mL of fly food. For extract doses (0.6, 0.8, 1, 0.0125, 0.025, and 0.05 g/mL) the dry concentrated root extract was dissolved in distilled water in a beaker at room temperature and warm food (50°C) was added and mixed before aliquoting approximately 1.5 mL per testing vial. For sodium valproate, one uncoated 100 mg tablet (Agog Pharma Ltd, India) was crushed, 0.3 mg or 0.15 mg were suspended in 1 mL distilled water each, and the drug suspension was then mixed with 5 mL of warm food to produce low and high sodium valproate concentrations respectively; SV 0.3 mg/mL and 0.15 mg.
Administration of the extract and standard control drug were done following previous methods.46,49,64,65 For acute study models, 10-day-old flies in replicates of 10 flies per plastic fly vial (Genesee Scientific, 32-116, USA), were starved for 15 h to induce appetite, after which they were fed 1 g of standard yeast/cornmeal/agar media without extract (negative or mutant controls and wild-type controls for acute study), or 1 g of standard media mixed directly with 0.6 g/mL, 0.8 g/mL, or 1 g/mL extract (extract tests for acute study), or 1 g of standard media mixed with 0.3 mg/mL SV (standard controls for acute study), and for the acute tests, the flies were allowed to feed for 1, 2, or 3 h (each treatment was tested for 1-3 h) before being transferred into individual empty plastic fly vials (Genesee Scientific, 32-116, USA) that were capped with a cotton plug. The flies were allowed to sit undisturbed for 20 min before tests were conducted.46,49,64,65 For chronic experiments, 10-day-old flies at the start of experiments in replicates of 10 flies per plastic fly vial (Genesee Scientific, 32-116, USA), were fed 1 g of standard yeast/cornmeal/agar media without extract (mutant/negative controls and wild-type controls for chronic study) or 1 g of standard media mixed directly with 0.0125 g/mL, 0.025 g/mL or 0.05 g/mL extract (extract tests for chronic study), or 1 g of standard media mixed with 0.15 mg/mL SV (standard controls for chronic study), and the flies were transferred to fresh plastic fly vials (Genesee Scientific, 32-116, USA) every 3 days. The flies for chronic studies were allowed to feed for 6, 12, or 18 days (each treatment was tested for 6-18 days) before being transferred into individual empty plastic fly vials (Genesee Scientific, 32-116, USA) which were capped with cotton plug, and the flies were left to sit undisturbed for 20 min after which the tests were conducted.46,49,64,65
Analytical Procedures
Phytochemical Analysis of I. cylindrica Root Extract
Phytochemical analysis of the methanol root extract of the plant was performed in triplicates in the Ethnobotany Laboratory, Department of Biological Sciences, Makerere University in Uganda. Qualitative analysis of the main bioactive compounds of the plant (alkaloids, saponins, tannins, cardiac glycosides, flavonoids, phenols, terpenoids, etc)34,35,66 was carried out using standard qualitative analytical methods described previously.67,68 Quantitative analyses of the isolated compounds of the crude extract of the herb were carried out following standard spectrophotometric analytical methods (PerkinElmer's Lambda 35 UV/Vis, USA) described previously.69-75
In Vivo Anticonvulsant and Antiepileptic Effects of I. cylindrica Root Extract on Mutant Flies
Determination of Bang-sensitivity Behavior and Seizure Parameters of Flies
To determine the bang sensitivity and seizure characteristics of the flies, we subjected the mutant flies (parabss1) and wild-type controls (OrR and CS) in different groups to mechanical stress, then observed for the occurrence of bang sensitivity and measured the time taken during convulsions, paralysis, to recovery from the paralysis.62,63 Bang sensitivity and convulsion parameters (mean convulsion time, CT; mean paralysis time, PT; and mean recovery time, RT, were assessed following previous methods.49,62,63,78 Briefly, on the day of the behavioral assessment, each fly was transferred into an empty plastic cylindrical 10 mL vial capped with a cotton plug, the fly was mechanically stimulated by placing an inverted vial on a bench-top vortex, then vortexed at high speed (maximum speed of 3200 rpm for 10 s, at room temperature of 22- 30°C) with a vortex mixer (Benchmark Scientific, BMK-BV1000, USA). The durations spent during convulsions, paralysis, and recovery for each animal to right itself (recover) were recorded as CT, PT, and RT respectively (QIANZICAI Stopwatch 382 ®). CT is the time taken while the fly is experiencing a convulsion (seizures, shaking, and lying on its back); paralysis is the second phase after a convulsion; PT is the time between the end of a convulsion and recovery; recovery is when the fly is standing and exhibiting normal mobility or flight; RT is the time taken by a fly that exhibits bang-sensitive behavior to recover completely from the convulsion.49,62,63,78 The mean of the convulsion, paralysis, and recovery times (CT, PT, and RT) of the 50 flies in each group were calculated as follows: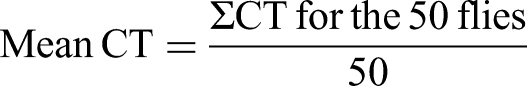


Determination of learning and Memory Pass Rates of Flies
The learning and memory assays were performed at 22–30°C using the Aversive Phototaxic Suppression (APS) assay previously described57,79 with a modified T-maze set-up.
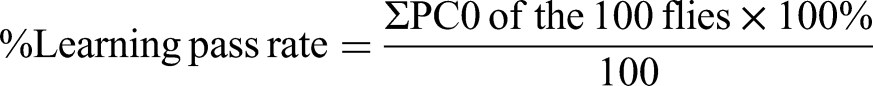

Histopathological Examination of Brain Tissues of Flies
Histopathological tests were performed using standard histological methods as previously described.78,80-82 Briefly, on the day of histological assessments, Drosophila melanogaster flies were anesthetized with CO2 (Fly CO2 anesthesia setup; Genesee Scientific, 59-114/54-104M, USA), and placed on a CO2 perfused pad for collecting. The flies were decapitated under a dissecting stereo microscope (Leica Microsystems, Leica M60 CMO), and the heads were fixed in 10% neutral buffered formalin (NBF) fixative at room temperature (22- 30°C) for 24 h, after which tissues were processed using routine histology techniques by dehydrating with graded alcohols, clearing with xylene, and embedding into paraffin wax using an automated tissue processor (Histokinette-SLEE MAINZ, MTP type). Every tissue was randomly sectioned into six, 6 μm-thick transverse histological sections using a rotary microtome (SLEE MAINZ, CUT4062), and the sections were then placed on slides and stained with hematoxylin and eosin (H &E) and combined Luxol fast blue (LFB) and Nissl (Klüver's) staining techniques following standard protocols.78,80-82 The stained sections were mounted in mounting media and qualitative histological examination of the sections was done and photographed with a light microscope (Nikon Eclipse Ci-L Upright Microscope
A qualitative examination of the tissues was done using previously described methods64,83-85 where brain neurodegeneration was categorized as normal, moderate, or severe based on the size and frequency of brain vacuolations in H and E stained-sections following previous methods.84,85 The non-myelinating Schwann cells in mammals are comparable to the ‘ensheathing’ glial cells within the CNS of Drosophila which encase axons and neuropil of the flies,86,87 therefore, LFB in Klüver LFB stain was used to demonstrate axonal degeneration of neurons rather than axonal demyelination.
88
The nature of the Nissl substance and nerve tracts (axons) were demonstrated using the Klüver LFB-stained tissues following previous methods,64,83 where the nerve tracts were shown by blue color and the Nissl substance was shown by magenta (violet) colour. A weak LFB stain (light blue patchy areas) indicated axonal degeneration of nerve tracts, while a weak cresyl echt violet stain (light violet) indicated abnormalities in Nissl substance,64,83 supplementary file 2
Statistical Analysis
The data was recorded and organized into an excel spreadsheet (version 219), then imported into Graph Pad Prism Version 6 software for statistical analysis. Data were subjected to one-way analysis of variance (ANOVA) and two-way ANOVA for convulsion and learning/memory parameters respectively and Tukey's multiple comparison tests were used to determine sources of variation, and significant differences (P < 0.05) at a 95% confidence interval were indicated with different superscript letters (a, b, c, d, e, f, and g). The information was expressed as mean ± SD and presented in graphs and tables. For easy graphical visualization and representation of brain histopathological findings of the flies, histological data were also graphically represented using the GraphPad Prism software, and the grouped interleaved bars were used to represent the general brain morphology, and grouped stacked bar graphs were employed to represent the brain axonal morphology of flies.
Results
Phytochemical Composition of I. cylindrica Root Extract
Phytochemical analysis of the methanol I. cylindrica root extract showed the availability of saponins, flavonoids, tannins, cardiac glycosides, polyphenols, alkaloids, and chromone derivatives (
Total Phytochemical Content in the Methanol Root Extract of I. cylindrica.

Values represent means ± standard deviations (SD) for triplicate experiments.
Juvenile-Adult and Old-Adult bss Paralytic Mutant Flies Portray Acute and Chronic Bang-Sensitivity and Seizure Phenotypes
Bang-sensitive assays revealed that the wild-type control flies (OrR and CS) were unaffected by mechanical stress (did not experience bang-sensitivity), while the mutant control flies of acute and chronic studies in the same groups (I and VI) displayed bang-sensitivity. This indicates that juvenile adults (10-day-old post-eclosion) and old-adults (16-28-day-old post-eclosion) mutant flies show acute and chronic bang-sensitivity, unlike the wild-type controls which show normal phenotypes (Figure 1A).
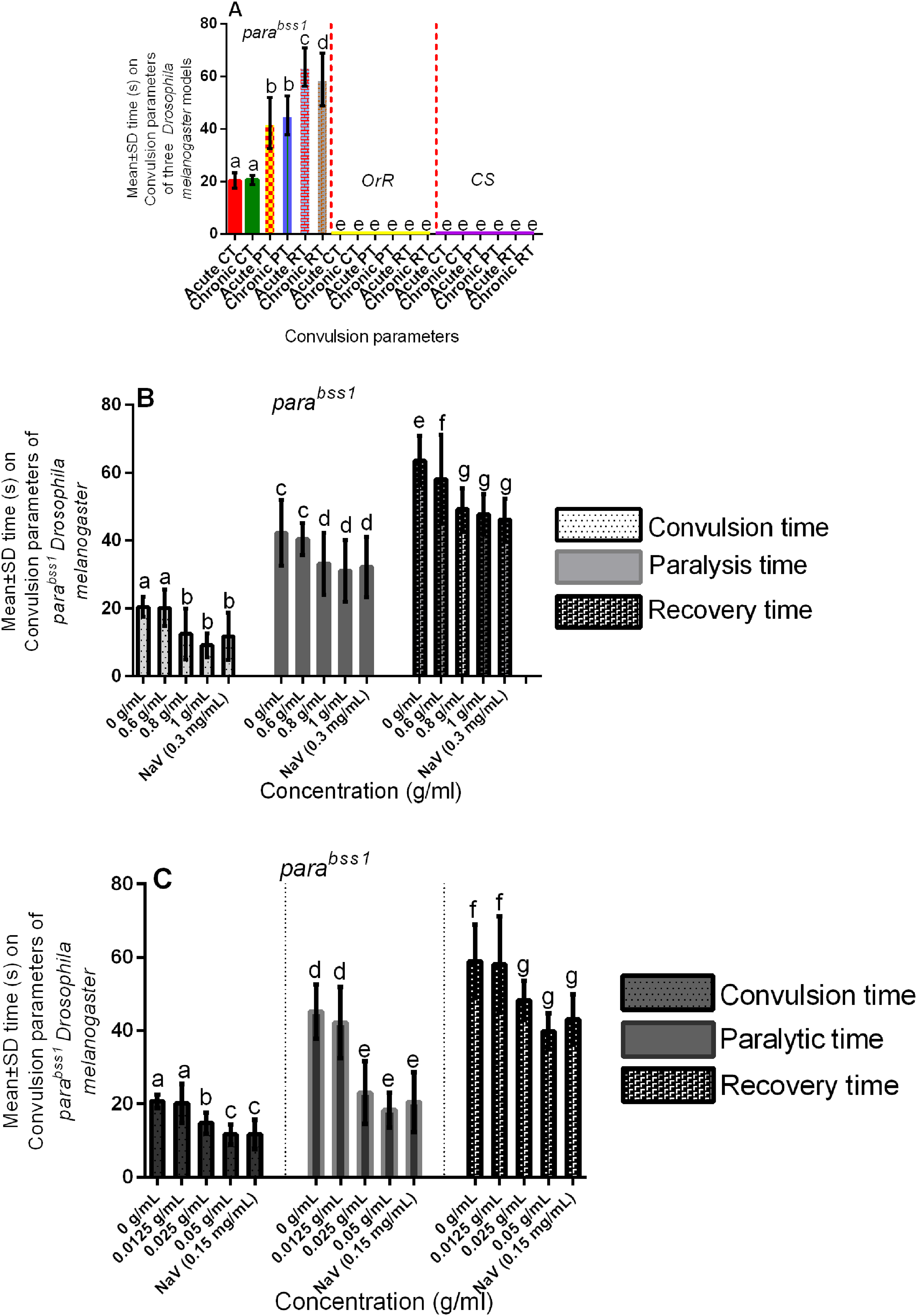
Acute and chronic effects of methanol I. cylindrica root extract on bang-sensitivity and seizure phenotypes of male juvenile-adults (10-day-old) and old-adults (22-day-old) bang-sensitive mutant Drosophila (parabss1). A. Acute and chronic seizure parameters (CT, PT, and RT) of juvenile-adults and old-adults; mean-time (y-axis) plotted for the three parameters. B. Acute (2-h) extract treatment alleviates acute seizures of juvenile adults. C. Chronic (12-day) extract treatment alleviates chronic seizures of old adults; mean time (y-axis) plotted against different treatment groups. OrR and CS = wild-type controls; parabss1 = mutant (test) model. Each value is expressed as mean ± SD; n = 50. Different letters (a, b, c, d, e, f, and g) meant significant differences among groups, (p < 0.05). Abbreviations: BSS = bang-senseless, CS = Canton-Special, CT = mean convulsion time, NaV = sodium valproate, OrR = Oregon R, Para = Paralytic, PT = mean paralysis time, RT = mean recovery time. Juvenile-adult and old-adult bss mutant flies showed bang-sensitivity and seizure phenotypes unlike the wild-type controls (A). The acute bang-sensitivity and seizure phenotypes of mutant flies were alleviated by acute (2 h) extract feeding in a dose-dependent pattern similar to SV (B). The chronic bang-sensitivity and seizures of mutant flies were alleviated by chronic (12 days) extract feeding similar to SV (D).
The bss paralytic mutant Drosophila (parabss1) were used in our subsequent convulsion experiments to determine their convulsion characteristics (convulsion, paralysis, and recovery times), and the wild-type controls that showed neither bang-sensitivity nor convulsion phenotypes were only used for purposes of controlling the experiments. Bang-sensitivity tests revealed that the mean CT, PT, and RT of juvenile-adult mutant controls in group I of the acute experiments were 20.46 s, 42.26 s, and 63.6 s respectively, and the average CT, PT, and RT of the old-adult mutant controls in group VI of the chronic tests were 20.74 s, 45.24 s, and 58.98 s respectively, with no significant (P > 0.05) difference between the acute and chronic CT and PT, but with significant (P < 0.05) difference between the acute and chronic RT
I. cylindrica Root Extract Controlled Acute and Chronic Bang-Sensitivity and Convulsion Phenotypes of bss Paralytic Mutant Flies Following Short-Term and Prolonged Treatments
The mean CT, PT, and RT of the untreated mutant control or negative control group (group I) were significantly (P < 0.05) elevated compared to the moderate and high-dose extract test groups (groups III and IV) in acute experiments, with no significant (P > 0.05) difference between SV group (group V) compared to the moderate and high-dose extract test groups, but with significant (P < 0.05) difference between the low-dose group (II) compared to SV, moderate and high-dose extract test groups (III, IV, and V), (Figure 1B, Table 4, and Supplementary file 3: Table 2). The results indicate that short-term ingestion of the extract similar to SV protects against acute seizures of the mutants in a dose-dependent fashion, with 0.8 mg/ml and 1 g/mL extract, and 0.3 mg/mL SV being effective seizure-controlling doses compared to 0.6 g/mL extract.
Multiple Comparisons of Acute and Chronic Effects of Methanol I. cylindrica Root Extract on Acute and Chronic Seizure Parameters of Male Juvenile-Adult (10-day-old) and old-Adult (22-day-old) Bang-Sensitive Mutant Drosophila (parabss1), n = 50.

The mean CT, PT, and RT of the untreated mutant controls (group VI) were significantly (P < 0.05) elevated compared to the moderate and high-dose extract test groups (groups VIII and IX) in chronic experiments, with no significant (P > 0.05) difference between SV group (X) compared to the moderate and high-dose extract test groups (VIII and IX), but with significant (P < 0.05) difference between the low-dose group (VII) compared to SV group, moderate and high-dose extract test groups (VIII, IX, and X), (Figure 1C, Table 4, and Supplementary file 3: Table 2). The results indicate that prolonged ingestion of the extract similar to SV protects against chronic seizures of the mutants in a dose-dependent fashion, with 0.025 mg/mL and 0.050 g/mL extract, and 0.15 mg/mL SV being effective seizure-controlling doses compared to 0.0125 g/mL extract.
Juvenile-Adult and Old-Adult bss Paralytic Mutant Flies have Acute and Chronic Learning and Memory Impairments
APS assay revealed the average learning and memory pass rates of wild-type controls as being78% and 71% respectively and were significantly (P < 0.05) higher than the learning and memory pass rates of the mutant controls (40% and 35% respectively) in group XI of the acute tests, with no significant (P > 0.05) difference in the average learning and memory pass rates of the mutant flies at 1, 2, or 3 h within the mutant controls of group XI (Figure 2A, Table 5 and Supplementary file 3: Table 3). This indicates that juvenile-adult (10-day-old) mutant flies show acute non-progressive deficits in learning and memory performance compared to the wild-type control group in the acute phases of the study.

Acute and chronic effects of methanol I. cylindrica root extract on acute and chronic learning and memory deficits of male juvenile-adults (10-day-old) and old-adults (16–28-day-old) bang-sensitive mutant Drosophila (parabss1). A. Acute learning and memory deficits of juvenile-adult bang-sensitive mutant Drosophila. B. Acute (1–3-h) extract treatment alleviates acute learning and memory deficits of juvenile-adult mutant flies; mean learning and memory pass rate for mutant flies (y-axis) plotted against time (hours) with respect to the concentration of extract. C. Chronic learning and memory deficits in old-adult bang-sensitive mutant Drosophila; mean learning and memory pass rate (y-axis) plotted against time (days). D. Chronic (6–18-day) extract treatment alleviates chronic learning and memory deficits in old-adult mutant flies; mean learning and memory pass rate for mutant flies (y-axis) plotted against time (days) with respect to the concentration of extract; mean learning and memory pass rates (y-axis) plotted against time with respect to different treatment groups. OrR and CS = wild-type controls; parabss1 = mutant (test) model. Each value is expressed as mean ± SD; n = 100. Different letters (a, b, c, and d) meant significant differences among groups, (p < 0.05). Abbreviations: BSS = bang-senseless, CS = Canton-Special, LPR = learning pass rate, MPR = memory pass rate, NaV = sodium valproate, OrR = Oregon R, Para = Paralytic. Juvenile-adult and old-adult bss mutant flies showed impaired learning and memory function compared to the wild-type controls with normal learning and memory function (A & C). The acute learning and memory functions of the mutant flies were rescued by acute (1–3 h) extract feeding in a duration-independent pattern similar to SV (B). The chronic learning and memory functions of the mutant flies were rescued by chronic (6–18 days) extract feeding in a duration-independent pattern similar to SV (D).
Multiple Comparisons on Acute and Chronic Learning and Memory Deficits of Male Juvenile-Adults (10-day-old) and Old-Adults (16–28-day-old) Bang-Sensitive Mutant and Wild-Type Control Drosophila at 0 g/ml. n = 100.

OrR and CS = wild-type controls; parabss1 = mutant (test) model. p < 0.05 meant significant differences among the groups. Abbreviations: BSS = bang-senseless, CS = Canton-Special, LPR = learning pass rate, MPR = memory pass rate, NaV = sodium valproate, OrR = Oregon R, Para = Paralytic.
Also, the assay revealed that the average learning and memory pass rates of wild-type controls (78% and 71% respectively) were significantly (P < 0.05) higher than that of the mutant controls (35% and 33% respectively) in the group XIV of the chronic tests, with no significant (P > 0.05) difference in the mean learning and memory pass rates of the mutant flies at 6, 12, or 18 days within the mutant controls of group XIV (Figure 2C, Table 5 and Supplementary file 3: Table 4). This indicates that old-adult (16-28-day old) mutant flies show chronic non-progressive deficits in learning and memory performance compared to the wild-type control group in the chronic phases of the study.
I. cylindrica Root Extract Controlled Acute and Chronic Learning and Memory Impairments of bss Paralytic Mutant Flies Following Short-Term and Prolonged Treatments
The average learning and memory pass rates of untreated mutant controls or negative controls (group XI) were significantly (P < 0.05) decreased compared to the extract test group (XII) of acute experiments, with no significant (P > 0.05) difference between the SV group (XIII) and extract test group (XII), and with no significant (P > 0.05) difference of learning and memory pass rates after 1, 2, or 3 h of treatment in the extract test group (XV), (Figure 2B, Table 6 and Supplementary file 3: Table 3). The results indicate that short-term ingestion of 0.8 g/mL extract (similar to 0.3 mg/mL SV) alleviates the acute learning and memory deficits of the mutant flies in a time-independent fashion during the acute phase of the study.
Multiple Comparisons on the Acute and Chronic Effect of Methanol I. cylindrica Root Extract on Acute and Chronic Learning and Memory Deficits of Male Juvenile-Adult (10-day-old) and old-Adult (16–28-day-old) Bang-Sensitive Mutant Drosophila (parabss1). n = 100.

p < 0.05 meant significant differences among groups. Abbreviations: BSS = bang-senseless, CS = Canton-Special, LPR = learning pass rate, MPR = memory pass rate, NaV = sodium valproate, OrR = Oregon R, Para = Paralytic.
The mean learning and memory pass rates of untreated mutant controls (group XIV) were significantly (P < 0.05) decreased compared to the extract test group (XV) in chronic tests, with no significant (P > 0.05) difference between the SV group (XVI) and extract test group (XV), and with no significant (P > 0.05) difference of learning and memory pass rates following 6, 12, or 18 days of treatment in the extract test group (XV), (Figure 2D, Table 6 and Supplementary file 3: Table 4). The results indicate that prolonged ingestion of 0.025 g/mL extract (similar to 0.15 mg/mL SV) protects against the chronic learning and memory deficits of the mutant flies in a duration-independent pattern during the chronic phases of the study
The acute and chronic APS assays in mutant and wild-type control groups showed significantly (P < 0.05) higher average learning pass rates compared to the corresponding average memory pass rates within each experimental group (Figure 2A-D, Table 5, and Table 6). This indicates that in general terms, the learning performance of flies is higher than their memory performance in both acute and chronic phases of the study, this disparity is probably due to the method used to obtain the two parameters.
Juvenile-Adult and Old-Adult bss Paralytic Mutant Flies Show Acute and Chronic Brain Neurodegeneration
Histopathological examination of H & E-stained brain tissues found no significant brain lesions (focal brain vacuolations ˂ 3 µm in diameter) in wild-type controls of group XI, while mutant (negative) controls of the group (XI) showed moderate brain neurodegeneration (multifocal vacuolations, 3-5 µm in diameter) in acute tests, with a similar nature of brain histopathology in mutant controls of group XI at 1, 2, or 3 h (Figure 3A and 4, and Supplementary file 3: Table 5). This indicates that juvenile-adult (10-day-old) mutant flies show acute non-progressive brain neurodegeneration compared to the juvenile-adult wild-type controls which depict normal general brain histology in the acute phases of the study.
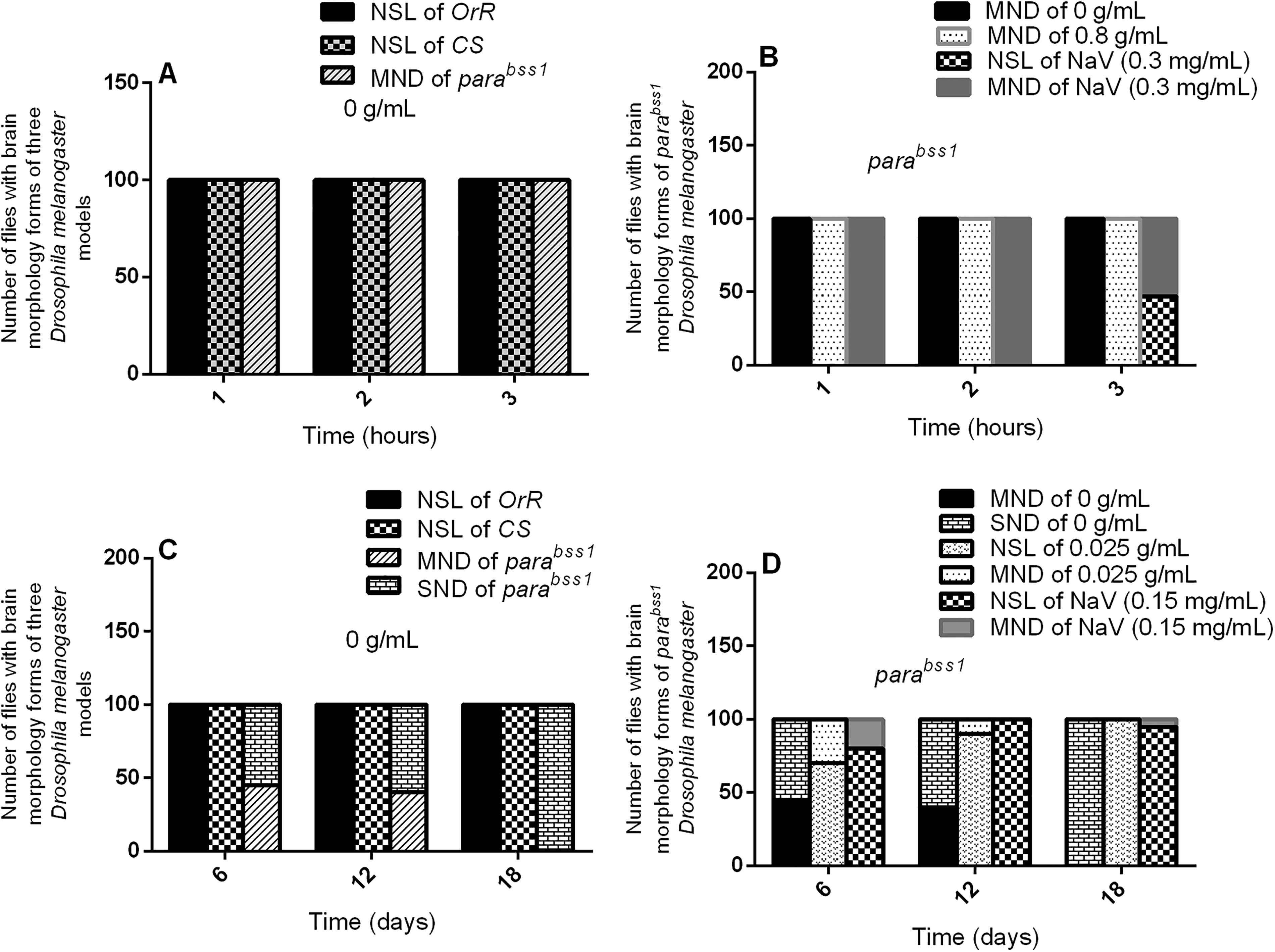
Acute and chronic effects of methanol I. cylindrica root extract on acute and chronic brain neurodegeneration in male juvenile-adult (10-day-old) and old-adult (16–28-day-old) bang-sensitive mutant Drosophila (parabss1). A. Acute brain neurodegeneration of juvenile-adult bang-sensitive mutant Drosophila. B. Effect of acute (1–3-h) extract treatment on acute brain neurodegeneration of juvenile-adult mutant flies. C. Chronic neurodegeneration of old-adult bang-sensitive mutant Drosophila. D. Effect of chronic (6–18-day) extract treatment on chronic brain neurodegeneration of old-adult mutant flies. The number of flies having each type of brain morphology (y-axis) plotted against time concerning different treatment groups; n = 100. OrR and CS = wild-type controls; parabss1 = mutant (test) model. Abbreviations: BSS = bang-senseless, CS = Canton-Special, NaV = sodium valproate, NSL = no significant lesion, MND = moderate neurodegeneration, OrR = Oregon R, Para = Paralytic, SND = severe neurodegeneration. 100% of juvenile-adult (10-day-old) bss mutant flies showed moderate brain neurodegeneration versus the wild-type controls of the same age with normal brain morphology/NSL (A). The acute brain neurodegeneration of mutants was not alleviated by acute extract feeding versus alleviation by acute feeding with SV (B). 100% of old-adult (16–28-day-old) bss mutant flies showed moderate-severe brain neurodegeneration versus the wild-type controls of the same age with NSL (C). The chronic brain neurodegeneration of mutants was rescued in 100% of mutant flies by chronic (18 days) extract feeding similar to feeding with SV (D).

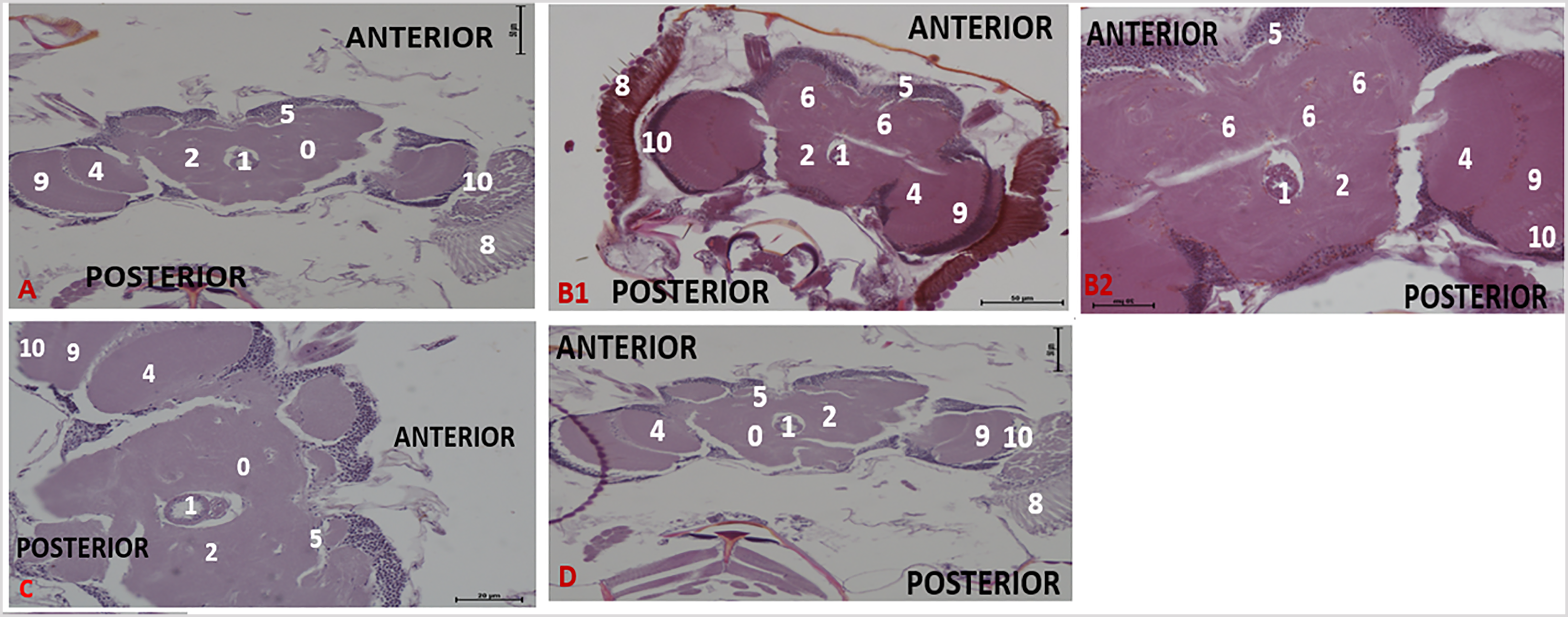
Effect of acute treatment with methanol I. cylindrica root extract on acute brain neurodegeneration of male juvenile-adults (10-day-old) bang-sensitive mutant Drosophila (parabss1). n = 100, (H&E; transverse sections at level of mid-brain, x200 or x400). Bar, 20 µm or 50 µm. A. Wild-type control, x200. B. Mutant control, x400. C. Standard control, x200. D. Methanol extract test, x200 (D1), x400 (D2). Categories of brain neurodegeneration: NSL (vacuolations <3 µm in diameter) = Normal; MND, (vacuolations 3–5 µm in diameter); SND, (vacuolations >5 µm in diameter). OrR and CS = wild-type controls; parabss1 = mutant (test) model. 0 = No significant lesion in the neuropil, 1= Oesophagus; 2= Central brain; 3= Moderate vacuolations in neuropil; 4= Optic lobe; 5= Cell bodies; 7= Ventral nerve cord; 8= Compound eyes; 9= Medulla; 10= Lamina. Abbreviations: BSS = bang-senseless, CS = Canton-Special, NaV = sodium valproate, NSL = no significant lesion, MND = moderate neurodegeneration, OrR = Oregon R, Para = Paralytic, SND = severe neurodegeneration. 100% of juvenile-adult (10-day-old) bss mutant flies showed moderate brain neurodegeneration (B) versus wild-type controls of the same age with NSL (A). The acute brain neurodegeneration of mutants was not alleviated by acute extract feeding (D) versus alleviation by acute feeding with SV (C). See the bar graphs in Figure 3 above for a summary.
Histopathological examination of H & E-stained brain tissues showed no significant brain lesions (focal brain vacuolations ˂ 3 µm in diameter) in wild-type controls of group XIV, while mutant controls of the group (XIV) showed age and duration-dependent progressive brain neurodegeneration during the chronic phase of the study, as follows: 45% of animals showed moderate brain neurodegeneration and 55% of them showed severe brain neurodegeneration on day 6; 40%, moderate brain neurodegeneration and 60%, severe brain neurodegeneration on day 12; and 100%, severe brain neurodegeneration on day 18 (Figure 3C and 5, and Supplementary file 3: Table 6). This indicates that old-adult (16-28-day old) mutant flies show chronic age-dependent progressive brain neurodegeneration, unlike old-adult wild-type controls that depict normal general brain histology during chronic phases of the study.
Effect of chronic treatment with methanol I. cylindrica root extract on chronic brain neurodegeneration of male old-adult (16-28-day-old) bang-sensitive mutant Drosophila (parabss1). n = 100, (H&E; transverse sections at level of mid-brain, x200 or x400). Bar, 20 µm or 50 µm.
I. cylindrica Root Extract Lessened Brain Neurodegeneration of bss Paralytic Mutant Flies Following Prolonged but not Short-Term Treatment
The acute brain histopathological features (with H & E-stained tissues) of untreated mutant controls (group XI) were similar to the extract test group (XII) in acute tests, with a marked difference between histopathological findings of SV group (XIII) and extract test group (XII), ie, SV group showed improved brain histology from moderate brain neurodegeneration to normal brain forms in 47% of the animals after 3 h of treatment with SV (Figure 3B and 4, and Supplementary file 3: Table 5). Therefore, short-term ingestion of 0.8 g/mL extract (unlike SV) does not reduce the acute brain neurodegeneration of juvenile-adult mutant flies during the acute phase of the study.
The brain histopathological features (with H & E stain) of untreated mutant controls (group XIV) were markedly defective compared to the extract test group (XV) in the chronic phase of the study. The extract test group (XV) showed a noticeable alleviation of brain neurodegeneration in a duration-dependent fashion as follows: 70% of animals, showed no significant brain lesions, and 30%, had moderate brain neurodegeneration on day 6; 90%, had no significant brain lesions, and 10%, moderate brain neurodegeneration on day 12; 100%, no significant brain lesions on day 18. SV group (XVI) showed similar brain histological features as the extract test group (XV) during the chronic phases of the study as follows: 80% of animals, no significant brain lesions and 20% moderate brain neurodegeneration on day 6; 100%, no significant brain lesions on day 12; 95% no significant brain lesions and 5% of the animals regressed to moderate brain neurodegeneration on day 18, (Figure 3D and 5, and Supplementary file 3: Table 6). The results indicate that prolonged ingestion of 0.025 g/mL extract (like SV) alleviates chronic brain neurodegeneration of old-adult mutant flies in a duration-dependent fashion during the chronic phase of the study. Also, prolonged administration with SV (beyond 12 days) may be detrimental to the improved brain histology of the mutant flies.
Juvenile-Adult and Old-Adult bss Paralytic Mutant Flies Portray Acute and Chronic Brain Axonal Degeneration
Histopathological examination of axonal morphology and Nissl substance on Klüver Luxol Fast Blue-stained tissues found no significant lesions in axons and Nissl substance of wild-type controls of group XI, while mutant controls of the group (XI) showed axonal degeneration in the brain but with normal Nissl substance during the acute phase of the study, and with no differences in axonal morphology and Nissl substance of mutant flies at 1, 2, or 3 h within mutant control group (XI), (Figure 6A and 7, and Supplementary file 3: Table 5). This indicates that juvenile-adult (10-day-old) mutant flies show acute non-progressive axonal degeneration in the brain, compared to the juvenile-adult wild-type controls that depict normal axonal morphology in the acute phase of the study.
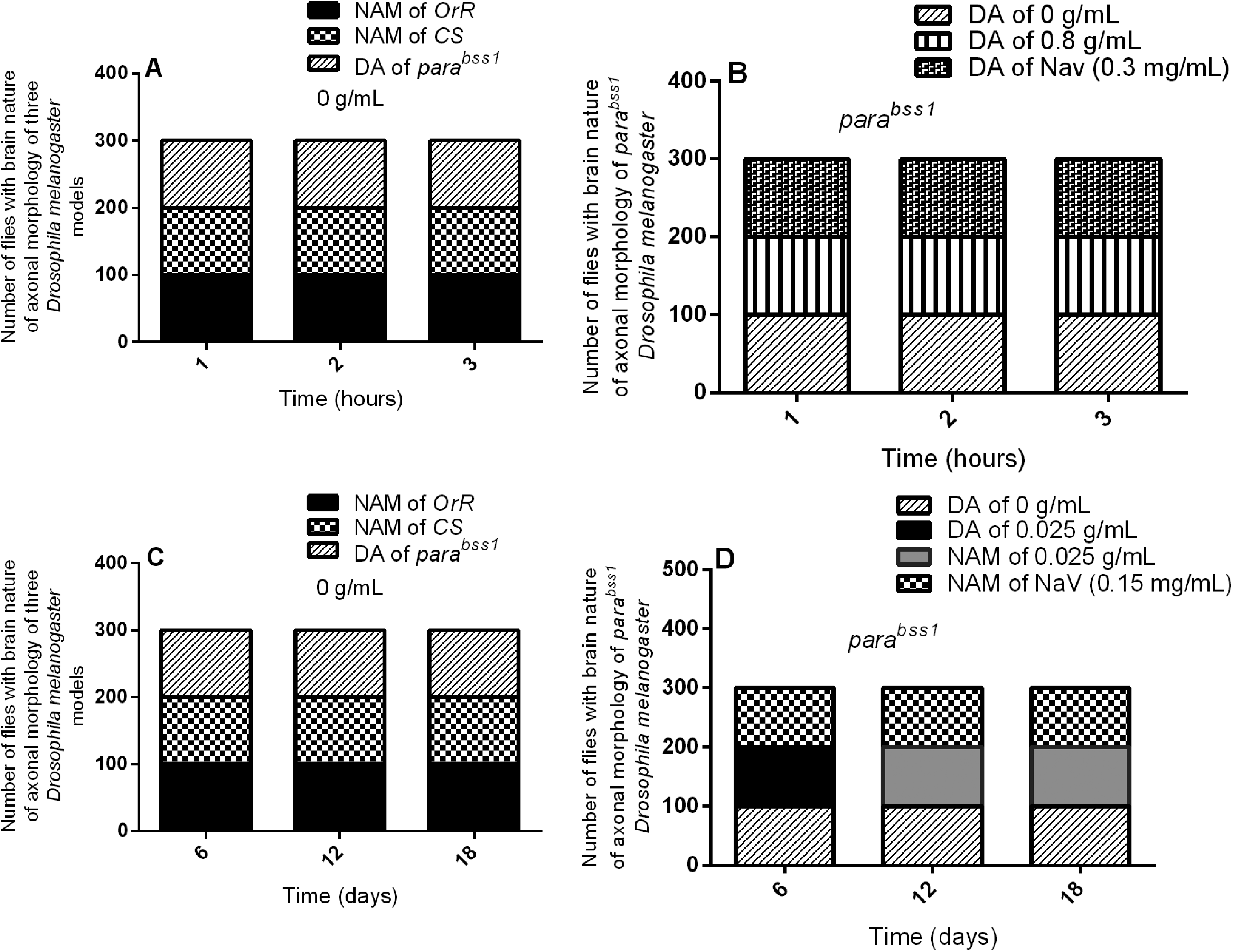
Acute and chronic effects of methanol I. cylindrica root extract on acute and chronic axonal degeneration in the brain of male juvenile adult (10-day-old) and old-adult (16–28-day-old) bang-sensitive mutant Drosophila (parabss1). A. Acute axonal degeneration in the brain of juvenile-adult bang-sensitive mutant Drosophila. B. Effect of acute (1–3-h) extract treatment on acute axonal degeneration in the brain of juvenile-adult mutant flies. C. Chronic axonal degeneration in the brain of old-adult bang-sensitive mutant Drosophila. D. Effect of chronic (6–18-day) extract treatment on chronic axonal degeneration in the brain of old-adult mutant flies. The number of flies with each type of brain axonal morphology (y-axis) plotted against time for different treatment groups; n = 100. OrR and CS = wild-type controls; parabss1 = mutant (test) model. Abbreviations: BSS = bang-senseless, CS = Canton-Special, DA = degenerated axons, NAM = normal axonal morphology, NaV = sodium valproate, OrR = Oregon R, Para = Paralytic. 100% of juvenile-adult (10-day-old) bss mutant flies showed axonal degeneration in the brain versus wild-type controls of the same age with normal brain axonal morphology (A). Acute brain axonal degeneration of mutants was neither alleviated by short-term extract feeding nor short-term feeding with SV (B). 100% of old-adult (16–28-day-old) bss mutant flies showed brain axonal degeneration versus wild-type controls of the same age with normal brain axonal morphology (C). Chronic axonal degeneration of mutants was rescued in 100% of mutant flies by prolonged (6–18 days) extract feeding, similar to feeding with SV (D).

Effect of acute treatment with methanol I. cylindrica root extract on acute axonal degeneration in the brain of male juvenile-adult (10-day-old) bang-sensitive mutant Drosophila (parabss1). n = 100, (Klüver Luxol Fast Blue; transverse sections, x200 or x400). Bar, 20 µm or 50 µm. A low-intensity patchy Klüver LFB stain indicates axonal degeneration/lesion in the Nissl substance. Axons are depicted in blue colour and Nissl substance is shown in magenta/ violet colour.
Histopathological examination of Klüver Luxol Fast Blue-stained tissues for axonal morphology and Nissl substance showed no significant lesions in axons and Nissl substance of wild-type controls in group XIV, however, the mutant controls in the group (XIV) showed brain axonal degeneration but with normal Nissl substance during the chronic phase of the study, and with no marked differences in nature of axon morphonology and Nissl substance of mutant flies at 6, 12, or 18 days within the mutant controls (group XIV), Figure 6C and 8, and Supplementary file 3: Table 6). This indicates that old-adult (16-28-day old) mutant flies show chronic age-independent non-progressive brain axonal degeneration compared to old-adult wild-type controls which depict the normal axonal structure and Nissl substance in chronic phases of the study.
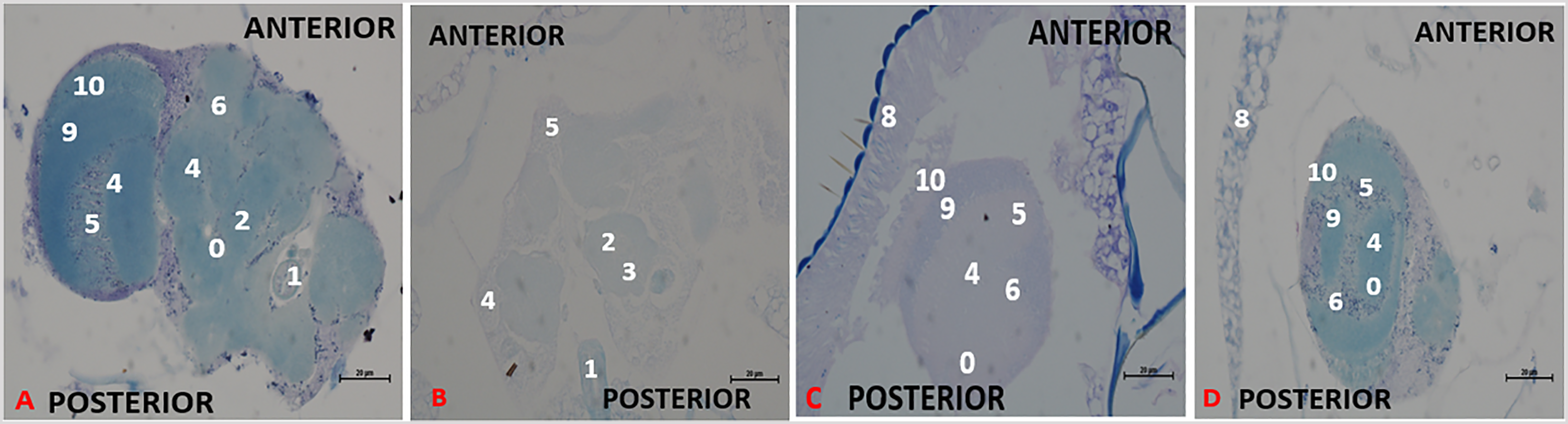
Effect of chronic treatment with methanol I. cylindrica root extract on chronic axonal degeneration in the brain of male old-adult (16-28-day-old) bang-sensitive mutant Drosophila (parabss1). n = 100, (Klüver Luxol Fast Blue; transverse sections, x200 or x400). Bar, 20 µm or 50 µm. A low-intensity patchy Klüver LFB stain indicates axonal degeneration/lesion in the Nissl substance. Axons are depicted in blue colour and Nissl substance is shown in magenta/ violet colour.
I. cylindrica Root Extract Alleviated Brain Axonal Degeneration of bss Paralytic Mutant Flies Following Prolonged but Not Short-Term Treatment
The acute brain axonal degeneration seen with Kluvers LFB-stained tissues of untreated mutant controls (group XI) was similar to the extract test group (XII) of the acute study, with no differences between SV group (XIII) and extract test group (XII), (Figure 6B and 7, and Supplementary file 3: Table 5), indicating that short-term ingestion of 0.8 g/mL extract (like 0.3 mg/mL SV) did not alleviate the acute brain axonal degeneration of juvenile-adult mutant flies during the acute phase of the study.
The chronic axonal degeneration was found in untreated mutant controls of the chronic study (group XIV), whereas the extract test group (XII) of the chronic study showed normal axonal morphology in 100% of treated mutant animals after days 6, 12, or 18 of treatment, and similar changes in axonal morphology were observed with the SV group (XVI), (Figure 6D and 8, and Supplementary file 3: Table 6), indicates that prolonged ingestion of 0.025 g/mL extract (like 0.15 mg/mL SV) alleviates the chronic brain axonal degeneration of old-adult mutant flies in a duration-independent pattern during the chronic phase of the study.
Discussion
The number of epilepsy cases has increased tremendously within the past few years and between 2019 and 2020 around 50 million people had the condition globally and almost 80% of these live in low- and middle-income countries.2,3 Imperata cylindrica (L.) P. Beauv (Poaceae) is a famous neuromedicinal, anticonvulsant, and antiepileptic herb in traditional therapy due to its receptor inhibition and antioxidant properties,32,34-38 however, to the best of our knowledge little is known regarding the neuroprotective efficacy of I. cylindrica as an antiepileptic herb and few studies have demonstrated the neuroprotective potential of the plant on glutamate-induced neurotoxicity in rat cerebral cortical cells.37,38 The study demonstrated that phytochemicals of I. cylindrica root extract had neuroprotective roles on convulsions, cognitive deficits, and brain histopathology of Drosophila melanogaster bang senseless mutant models of epilepsy. This is vital because little is known about the neuroprotective role of the phytochemicals,35,36,42 and in this study, the plant phytochemicals are thought to have upregulated endogenous antioxidants and inhibited receptor or voltage-gated sodium ion channels (VGSCs) which are linked to increased antioxidant status, decreased inflammation and apoptosis, increased tissue repair, and improved cell biology in the brain of the mutant flies faced with epilepsy-like neuropathological features.
The high contents of phytochemicals isolated from the methanol Imperata cylindrica (L.) P. Beauv (Poaceae) root extract (flavonoids, polyphenols, chromones
The current study has demonstrated that bang-senseless (bss) D. melanogaster mutant models (parabss) depict marked acute and chronic abnormal brain phenotypes (age-dependent progressive brain neurodegeneration and axonal degeneration) and significant neuropathological symptoms (acute and chronic convulsions and cognitive deficits or learning and memory deficits) in juvenile and old-adults arising from a gain-of-function point mutation in a para sodium channel gene at the para locus (up-regulates and increases the activity of the gene) in neurons of the current mutant D. melanogaster models of epilepsy (parabss1), unlike the wild-type controls (Oregon R and Canton-Special) which showed normal brain morphology, cognitive functions, and without bang sensitive and convulsion behaviors. Therefore, the abnormal brain phenotype and neuropathological features that were demonstrated in the mutant flies (parabss1), validate a mutation in the para gene (overexpression of para) in the neurons of brain tissues of juvenile and old-adult D. melanogaster bss mutants15,16 and is similar to the expression of a para gene demonstrated in neuronal brain tissues of adult D. melanogaster when UAS-para is driven by ELAV-GAL4 driver,101-104 and the expression of the mutant genes has been linked to human-like brain neurodegenerative disorders in Drosophila.101,103,105,106
Altered gene expression and tissue-specific expression of specific neurodegenerative mutant genes such as epilepsy mutant genes21,107 in D. melanogaster cause neuropathological phenotypes of the neurodegenerative disorder (in our case, epilepsy)21,108-110. The tissue-specific expression of mutant genes and transgenes in neurons has been linked to neuropathological phenotypes such as neurodegeneration, defects in mean lifespan, and locomotion in D. melanogaster models of neurodegenerative diseases and epilepsy.101-106,108,109,111 The expression of tissue-specific mutants and transgenes provides robust and precise models with which to study human neurodegenerative diseases, epilepsy, and possible preventive and curative treatments for neurological disorders.21,108-110
The neuropathological findings of the current study concur with previous studies that demonstrated bang-sensitive behaviour, convulsions in mutant paralytic D. melanogaster models of seizures overexpressing para gene (parabss1) due to a mutation of the gene15,16 and resulting reduction of seizure-stimulation threshold arising from increased expression of the VGSCs in the neurons of the mutants.15,16 The impaired learning and memory functions demonstrated in the current parabss1 mutant Drosophila compared to the wild-type controls are similar to the cognitive phenotypes previously shown in the model (parabss1),15-17 and concur with other findings that have shown cognitive deficits in Drosophila models of neurodegeneration (ELAV-GAL4 > UAS-GFP and ELAV-GAL4 > UAS-TAU). 57 Cognitive deficits in bang-sensitive paralytic D. melanogaster are attributable to a mutation of the para gene in neurons of the flies which disrupts the metabolic pathways and neuronal activities leading to bang-sensitive and seizure behaviors that originate in the brain of the flies (epileptogenic zone),15,53,83,112 and the seizures and their effects spread to secondary brain centers including the motor centers and learning and memory centers in mushroom bodies (cognitive centers) hence distorting the learning and memory functions (cognitive deficits) in the brain of D. melanogaster. 113 The study demonstrated that in general, the current parabss1 mutants have less magnitudes of memory function than the corresponding levels of learning function in all the study groups, the observed general differences in the two parameters come from the method (APS assay) that was employed to assess the parameters, ie, memory tests were performed 6 h after performance of the learning tests without re-training the flies to the task learnt. 57
The current parabss1 mutants show progressive age-related brain neurodegeneration (neurovacuolation) and axonal degeneration concurring with previous studies on bss and excitability Drosophila (parabss, jus, eas, tko, ses B) that found age-related brain neurodegeneration due to gene mutations in neurons of the flies.22,114,115 The axonal degeneration found in the current mutant models is typical of axonal-type neurodegenerative disorder (axonopathy) found in Drosophila models of human genetic neurodegenerative disease,116-118 and axonal degeneration is a cause of disease pathogenesis, disability, and loss of function in the CNS during epilepsy.119-121 It is characterized by a reduction in the density and number of nerve tracts as demonstrated previously in D. melanogaster bang-sensitive mutants (parabss, ses B, eas, jus, tko), 115 and is due to a mutation of the para gene at the para locus in neurons of bss para D. melanogaster models of epilepsy which induces defective nerve fibers.20,83,84,115,122 Similarly, mammalian forms of genetic epilepsy due to gene mutations in VGSC β subunit genes (SCN1B-SCN4B) and α subunit genes (SCN1A, SCN2A, SCN3A, and SCN8A) in the brain have been linked to neurodegeneration and pathology found in epilepsy.9-11,123
The neurodegeneration (degeneration of nerve tracts) in the brain of the bang senseless paralytic flies distorts electrical activity and conduction of impulses,124-126 contributes to the development of seizure and epileptic phenotypes (bang-sensitivity, convulsions, learning, and memory deficits) observed in the brain and peripheral parts of the current mutants, features of which are consistent with previous studies.124-126 In conclusion, the mutant flies of the study (parabss1) display fascinating epileptic phenotypes and behaviors similar to mutants of the BS paralytic family such as sdaiso,7,8 easPC, 80 and tko25t mutants, as well as other Drosophila models of human seizures10,15,17,22,114,115 ie, progressive age-related brain neurodegeneration and axonal degeneration of the mutant flies, are linked to bang-sensitive behavior, seizures, and defective cognitive functions, consistent with CNS neurodegenerative and axonopathy features of bss paralytic D. melanogaster seizure models.10,17,83,114,124-127
To the best of our knowledge, the study is the first of its kind to demonstrate the staining affinity of CNS axons for combined Luxol fast blue (LFB) and Nissl (Klüver's) stain in Drosophila models indicating that Drosophila neurons and axonal sheaths could be containing biological compounds mimicking the phospholipids or lipoproteins of the myelin sheath and Schwann cells of myelinated neurons. However, Drosophila axons are non-myelinated and not surrounded by Schwann cells, 117 instead, they are encased in ‘ensheathing glial cells’ within the CNS of Drosophila , 87 and although they are non-myelinated and not surrounded by Schwann cells,117,128,129 the current data concurs with previous studies which demonstrated Schwann-like (Drosophila) or Schwann cells (mammals) to be existing in unmyelinated axons in form of grooves but not surrounding the axons, closely interdependent with the axons to provide structural integrity and function of the axons.117,130-132 Also, Schwann cells have been reported to secrete neurotrophic factors and myelin-related proteins.130,133 The non-myelinating Schwann cells of mammals are comparable in function to the ‘ensheathing’ and ‘wrapping’ Drosophila glial cells of the CNS and peripheral nervous system of Drosophila respectively and are responsible for encasing axons and neuropil of the flies.86,87,134 Future studies should focus on the utility of combined Luxol Fast Blue and Nissl (Klüver's) staining technique in experimental examination of the basic neuronal structure in the CNS of Drosophila to establish the staining properties of the nervous tissues of Drosophila.
Acute and chronic extract treatment significantly reduced the neuropathological features of the mutant flies (convulsions, learning and memory deficits, and brain histopathology) in the extract test animals and the standard control group treated with SV in duration and dose-dependent fashions compared to the mutant (negative) control group, and the values of the parameters (CT, PT, RT, learning/memory pass rate, brain, and axonal neurodegeneration) of the extract test and standard control groups were similar to (not significantly different from) the wild-type control group indicating that our I. cylindrica extract induces significant neuroprotective activities (alleviates the neuropathological features) in the nervous tissue of the mutant flies, with a similar neuroprotective capacity to the standard control drug of our study (SV).
However, the acute and chronic age-related brain neurodegeneration and axonal degeneration of the mutant flies were alleviated by prolonged (6-18 days) extract treatment but not by short-term (1-3 h) treatment and the standard antiepileptic (SV) depicted better alleviative potential on the brain histopathology than the plant extract since the former modulated the brain histological defects of the mutant flies following both short-term (3 h) and prolonged (6-12 days) treatments compared to the extract which showed modulatory potential only after prolonged treatment (6-18 days). The standard antiepileptic showed a side-effect on brain morphology since further feeding with SV (18 days) caused brain histopathological features in the previously improved brain histology of the mutant flies indicating that although antiepileptic molecules have marked beneficial roles, they may cause detrimental side effects on the CNS, especially in cases of chronic antiepileptic therapy.135,136
The neuroprotective attributes of the current herb (I. cylindrica) are consistent with previous findings that demonstrated the phytochemicals of the herb to be possessing neuroprotective attributes on the neuropathology of rat brain cells37,38 and is in tandem with previous studies that suggest plant-based nonenzymatic antioxidants notably phenols, flavonoids, chromones, and glycosides to be contributors neuroprotection on brain cell damage by the provision of significant antioxidant activities (antioxidation mechanism controls oxidative stress markers in the brain of animals) that are linked to improved brain cell biology,37,39 thereby inducing anticonvulsant and antiepileptogenic pathways,40,137 and alleviate cognitive deficits of animals. 41 The antiepileptic similarity of the herb compared to standard anticonvulsant (SV) indicates that the extract similar to standard antiepileptics can alleviate the bang-sensitive behaviour of Drosophila models of epilepsy by improving brain cell biology, seizure-control, and cognitive-function modulatory pathways in concurrence with previous findings.15,61,127
Again, the potential of our extract to modulate the brain histopathology of the mutant flies is in tandem with a review by Trojnar et al, (2002) who demonstrated novel standard antiepileptics (lamotrigine, tiagabine, and topiramate) as having neuroprotective effects on CNS morphology of epileptic animals, 138 and concurs with previous studies that reported similar neuroprotectant potential in antioxidant compounds and antiepileptic drugs on epileptic animals.137,139 However, our findings contradict with findings of Qiao et al, (2000) where Vigabatrin demyelinated the brain of epileptic rats 140 indicating that antiepileptic molecules have side effects. 135 The differences and similarities between the current data and those by Trojnar et al, (2002), Qiao et al, (2000), and Cavanna et al, (2010) are striking but not surprising since antiepileptics can depict neuroprotective, neutral, or detrimental CNS activities in epileptic models.135,138,140
Oxidative stress has been strongly related to the pathogenesis of neurodegenerative disorders such as Alzheimer's disease, Parkinson's disease (PD), and epilepsy.115,141-146 Bang-sensitive Drosophila models of epilepsy are vital models for oxidative stress-related neurological defects, and the abnormal phenotypes in the models such as parabss, jus, eas, and tko are attenuated by feeding with an antioxidant. 115 In addition, attenuation and rescue of neurological defects by natural antioxidants have been reported in Drosophila models of epilepsy 143 and Drosophila is an emerging model organism for plant-derived antioxidants.147-149 Most importantly, the potential role of natural antioxidants from plant extracts as neuroprotectants for treating and preventing neurodegenerative diseases and epilepsy has been shown in vitro, in vivo, and in clinical settings.89,137,150 In addition, hyperactivity of the VGCs due to mutation (s) in the para gene (gain-of-function mutation in the para sodium channel gene at the para locus) in neuronal brain tissues of bang-sensitive (BS) Drosophila models of epilepsy is strongly linked to the neuropathological features of epilepsy (brain histopathology, seizures, and cognitive deficits).15,16
The brain has very high metabolic activities and is the most aerobically active organ in the body, the organ is very susceptible to oxidative stress, causing oxidative-induced brain injuries and neuronal hyperexcitability,
139
such as in bang-sensitive Drosophila and the defects can be rescued via the antioxidant pathways (feeding with neuroprotectant antioxidants),115,139 and by treating with molecules that inhibit the VGSCs.151,152 Therebefore, the proposed mechanisms by which I. cylindrica provides its neuroprotective attributes are the ‘antioxidant’ and ‘receptor inhibition’ mechanisms115,139,151,152 arising from the main phytochemicals of the plant (polyphenols, flavonoids, and chromones), these provide important biological activities within the brain cells including the improvement of antioxidant status, tissue repair, decreased inflammation and apoptosis, improvement of brain cell biology,37,39 and binding and inhibition of VGSCs and receptors (eg, 5-HT2B) by chromones
Given the weight of the flies tested (1 mg per fly) and that they can consume between 20–100 µg a day and twice this amount when starved for 14–17 h, 161 the doses used can be extrapolated to the human context eg 0.8 g/mL utilized in acute therapy, and 0.025 g/mL used in chronic therapy correspond to 16 g/kg and 0.5 g/kg respectively when projected to a human adult.
The current study had some limitations: it did not focus on the evaluation of the neuroprotective molecular mechanisms via the antioxidant signaling and receptor binding inhibition mechanisms, did not emphasize the gender-related differences associated with the abnormal phenotypes and effect of the herb on the phenotypes of the mutant flies, and only focused on the effect of the herb on a genetic form of epilepsy while leaving out acquired/induced convulsions like the antimuscarinic and pentylenetetrazole kindling models that are often studied137,162; a prospective study will be conducted to assess the practical attributes of antioxidant and receptor inhibition pathways related to the neuroprotective effects of I. cylindrica root extract using bss Drosophila, other genetic models and kindling models of convulsions.
Conclusion
The juvenile-adult and old-adult mutant flies (parabss1) show age-related progressive brain neurodegeneration and axonal degeneration and associated acute and chronic seizure and cognitive deficit phenotypes. The methanol root extract of I. cylindrica in a similar magnitude to sodium valproate exerts neuroprotective properties on the neuropathological features of the mutant flies by proposed antioxidant and receptor inhibition mechanisms arising from the main phytochemicals of the herb (Polyphenols, flavonoids, and chromones
Therefore, the methanolic extract contains high amounts of non-enzymatic antioxidants (phenols, flavonoids, and chromones
Supplemental Material
sj-docx-1-chp-10.1177_2515690X231160191 - Supplemental material for Attenuation of Seizures, Cognitive Deficits, and Brain Histopathology by Phytochemicals of Imperata cylindrica (L.) P. Beauv (Poaceae) in Acute and Chronic Mutant Drosophila melanogaster Epilepsy Models
Supplemental material, sj-docx-1-chp-10.1177_2515690X231160191 for Attenuation of Seizures, Cognitive Deficits, and Brain Histopathology by Phytochemicals of Imperata cylindrica (L.) P. Beauv (Poaceae) in Acute and Chronic Mutant Drosophila melanogaster Epilepsy Models by Fred Ssempijja, Samuel Sunday Dare, Edmund E. M. Bukenya, Keneth Iceland Kasozi, Ritah Kenganzi, Edgar Mario Fernandez and Marta Vicente-Crespo in Journal of Evidence-Based Integrative Medicine
Supplemental Material
sj-docx-2-chp-10.1177_2515690X231160191 - Supplemental material for Attenuation of Seizures, Cognitive Deficits, and Brain Histopathology by Phytochemicals of Imperata cylindrica (L.) P. Beauv (Poaceae) in Acute and Chronic Mutant Drosophila melanogaster Epilepsy Models
Supplemental material, sj-docx-2-chp-10.1177_2515690X231160191 for Attenuation of Seizures, Cognitive Deficits, and Brain Histopathology by Phytochemicals of Imperata cylindrica (L.) P. Beauv (Poaceae) in Acute and Chronic Mutant Drosophila melanogaster Epilepsy Models by Fred Ssempijja, Samuel Sunday Dare, Edmund E. M. Bukenya, Keneth Iceland Kasozi, Ritah Kenganzi, Edgar Mario Fernandez and Marta Vicente-Crespo in Journal of Evidence-Based Integrative Medicine
Supplemental Material
sj-docx-3-chp-10.1177_2515690X231160191 - Supplemental material for Attenuation of Seizures, Cognitive Deficits, and Brain Histopathology by Phytochemicals of Imperata cylindrica (L.) P. Beauv (Poaceae) in Acute and Chronic Mutant Drosophila melanogaster Epilepsy Models
Supplemental material, sj-docx-3-chp-10.1177_2515690X231160191 for Attenuation of Seizures, Cognitive Deficits, and Brain Histopathology by Phytochemicals of Imperata cylindrica (L.) P. Beauv (Poaceae) in Acute and Chronic Mutant Drosophila melanogaster Epilepsy Models by Fred Ssempijja, Samuel Sunday Dare, Edmund E. M. Bukenya, Keneth Iceland Kasozi, Ritah Kenganzi, Edgar Mario Fernandez and Marta Vicente-Crespo in Journal of Evidence-Based Integrative Medicine
Footnotes
Abbreviations
Acknowledgements
We thank the members of DrosAfrica, TReND (Teaching and Research in Natural Sciences for Development in Africa), and 4Sayansi for the helpful donation of laboratory equipment used in the present study at the Institute of Biomedical Research (IBR), Kampala International University, Western Campus, Bushenyi, Uganda, as well as for the technical support provided through conduction of workshops on the use of Drosophila models in Biomedical research in support of the study. We thank fellow members of our laboratory for their advice and helpful discussions throughout the project, Richard Baines and Bloomington Drosophila Stock Center, U.S.A for providing the mutant (parabss1) and wild-type (OrR and CS) fly stocks respectively.
Authors’ Note
Authors' Contributions
Fred Ssempijja: conceptualization, Principal Investigator, methodology, data collection, validation, data analysis and interpretation, writing—original draft preparation, and writing—review and editing. Samuel Sunday Dare: data analysis and interpretation, writing—review and editing, and supervision of this study. Edmund E. M. Bukenya: validation, writing—review and editing, and supervision of this study. Keneth Iceland Kasozi: validation, data analysis and interpretation, writing—original draft preparation, and writing—review and editing. Ritah Kenganzi: methodology, data collection, validation, writing—original draft preparation, and writing—review and editing. Edgar Mario Fernandez: conceptualization, methodology, and writing—review and editing. Marta Vicente-Crespo: conceptualization, methodology, validation, writing—review and editing, and supervision of this study. All authors have read and agreed to publish the version of the manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
The experimental protocol was approved by the Institutional Review Ethics Committee of Kampala International University, Western Campus, Uganda (study protocol code: MSC-ANA-001/142-DU), and carried out following the guidelines given by the Uganda Council for Higher Education (UCHE).
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Informed Consent
Not applicable
Trial Registration
Not applicable
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.