
Abstract
Background.
Mycobacterium tuberculosis has become the leading cause of morbidity and death in humans worldwide. Thus, genetic variability of the host plays a major role in human susceptibility to the pathogen, among others. Therefore, the objective of this finding was to assess the association of genetic polymorphisms of cytokines with tuberculosis infection.
Method.
A cross-sectional study was conducted between January and May 2018. Five ml of whole blood was collected and extracted the genomic DNA through simple salting out method. The patterns of genetic polymorphism were determined by amplification refractory method PCR using specific primers. Finally, the PCR run on electrophoresis of agarose gel and the band was visualized under UV light. A logistical regression model has been adapted to assess the association of genetic polymorphisms with tubercular infection. In order to determine the association between the explanatory and outcome variable, the odds ratio with 95% CI was calculated. P < 0.05 is a statistically significant value.
Result.
In present study, the frequency of TNF-α -308 G allele and GG genotype OR (95% CI)= 0.20 (0.11-0.37), and OR (95% CI)= 0.29 (0.18-0.46)), respectively) and IFN-γ +874 A allele and AA genotype OR (95% CI)= 3.80 (2.11-6.86) and (OR (95% CI)= 1.61(1.13-2.28), respectively) were significantly associated with tuberculosis incidence. In contrast, there is no significant correlation between IL-10 -1082 A and AA of allele and genotype, respectively in tuberculosis patients (p > 0.05) was evident.
Conclusion.
From our finding, the genetic variability of TNF-α -308 A and IFN-γ +874 alleles are the potent host genetic risk factors associated with tuberculosis infection.
Background
Tuberculosis (TB) is a disease caused by the Mycobacterium tuberculosis complex, including Mycobacterium tuberculosis (M. tb). Mycobacterium tuberculosis is commonest causative agents of TB. 1 In 2018 the report indicates, 106 people fell ill from TB, among those 1.76 were dying. In many countries with a high TB burden, including Ethiopia, Kenya, Lesotho, Namibia, Russian, Tanzania, Zambia, and Zimbabwe, TB exceeded 4% per year since 2010. 2
Furthermore, there is an estimation that about a third of the global population has M. tuberculosis infection, however, the active disease develops only in about 10% of the infected individuals. 3 Some patients which have underlying risk factors such as HIV infections and diabetes have a higher susceptibility of infection with TB. 4 On the other hand the development of active TB can be a result of environmental and genetic predisposition. 5 Furthermore, host genetic factor is a great contributor to the control of tuberculosis globally. 6 Evidences from genetic concordances in monozygotic and dizygotic twins as well as ethnic variations are important proofs showing that host genetic factors are relevant determinants of the susceptibility to M. tb infection. 7,8 The populations in the African setting, particularly the sub-Saharan Africa, are importantly informative in terms of genetic diseases fine mapping because of the huge level of diversity genetically and the demographic history. 9,10 Research using human mitochondrial DNA and nuclear DNA markers has consistently shown that Africa is the world most genetically diverse region. 11 Taking in to this account, The environment in this continent plus the TB naïve host genetic structures of the region may have contribute the rapid spread of the disease. 12 The immunological reactions to M. tb is due to the interaction of bacterial and environmental factors with the human host immune system (host genetics). Both genetics and acquired defects in host immune response pathways substantially raised the probability of infection severity. 9
Thus, genetic variation is the basis for human diversity and plays an important role in human diseases. 13 There are many genes encode for various numbers of cytokines, which have a crucial role in host susceptibility to tuberculosis. 14 Association between cytokine gene polymorphisms and the risk of TB development was examined in different forms with a different explanation. 15
Tumor Necrosis Factor-alpha (TNF-α) is a pro-inflammatory cytokine that found in chromosome 6, MHC III. 15 It is primarily secreted by a different immune cells, including macrophage, lymphocyte, neutrophils, mast cell, and endothelial cell. 16 The effect of TNF-α on the human body varies from the regulation of inflammatory response, activation of macrophage, regulation of interferon γ production and stimulating the production of IL-1and IL-6. 17 Therefore, TNF-α contributes in the control of M. tb infection by the induction of reactive nitrogen and reactive oxygen intermediates by a macrophage and early induction of chemokines. 18 A variety of single nucleotide polymorphisms (SNPs) have been found in the promoter region of the TNF-α gene. Among these, SNP at -308 at (G/A) position is the most important gene polymorphism, which associates with altered levels of pro-inflammatory mediators and the progression of numerous communicable diseases like TB. 19
Another important cytokine in the immunity of TB is interferon-gamma (IFN-γ), which is produced by activating T cells, natural killer cells, and macrophage. Its production plays a critical role in activating macrophages and promotes cell proliferation, adhesion and apoptosis. 18 IFN-γ induce a large number of ROIs and Nitric oxide production for controlling M. tb infection. 20,21 There is SNP +874 T/A located at the 5’ -end of a CA repeat at the first intron of human IFN-γ that can be associated with TB disease. 22,23 IL-10 is also an immunoregulatory cytokine that down-regulate the response of pro-inflammatory cytokine. 22 It can be involve by a number of intracellular microbial, including M. tb to hamper the host immune response, which results failure of subsequent clearance from the host. 24 In the IL-10 gene promoter region, SNP was associated with modified levels of circulating IL-10. 21 These polymorphisms were considered to be potentially predisposing factors for tuberculosis. With an increasing number of TB patients in Ethiopia, knowledge of genetic background and marker, which contribute to the etiology and progression of TB is important in the intervention and management of the disease. To date, as a controversial, it has been difficult to determine the exact relationship between These cytokine genotypes with respect to TB. Therefore, aim of this finding was to assess the association of genetic polymorphisms of TNF-α -308 G/A, IFN-γ +874 T/A and IL-10 -1082 G/A with tuberculosis infection.
Methods and Materials
Study Design, Sample Collection & Processing Procedure
An institutional based cross-sectional study was done between January and May 2018. Data was collected by using a structured questionnaires and interviews. For the study of the laboratory test, using EDTA coated tubes 5 ml of whole blood was collected from each study participant. The molecular technique was done in the molecular laboratory of immunology and the institute of the Biotechnology laboratory. The inclusive value of the laboratory work was maintained by strictly following the standard operating procedure.
DNA and Genotype Isolation
By simple salting out method according to standard protocol genomic DNA was extracted from human blood. 25 Purity of genomic DNA was estimated by the Nanodrop at 260/280 nm absorbance ratio having between 1.8 to 2.0 indicate the quality of DNA. 25,26 DNA was amplified in a different PCR reaction. We used specific designed primer as follows in Table 1.
Primers for Amplifications of TNF-α (-308), IFN-γ (+874) and IL-10-(-1082), 2018.

Genotyping of each cytokine gene was done using ARMS PCR with appropriate specific primers and PCR reaction conditions. The PCR reaction was performed in a total volume of 50 µl containing 200 ng of genomic DNA, 200 µM of each dNTP, 50 mM KCl, 1 mM each of the primers, and 1.5 U of Taq polymerase. The amplification of the PCR product was accomplished by denaturation for 5 minutes at 95°C, annealing for 1 minute at 52°C, and extension for 1 minute at 68°C to72°C with a repeated the cycle 35 times.
The amplified products were stained with ethidium bromide, run on gel electrophoresis, and analyzed in a UV light gel documentation system to determine allele and genotype frequency of TNF-α (- 308), IFN -γ (+874), and IL-10 (-1082) locus (Figure 1).

Shows the agarose gel electrophoresis image.
Statistical Analysis
Data were prepared and double entered in Epi Info 7 and transported to SPSS version 20 for data interpretation. The results were summarized and presented through tables. The variations of genotypes and allele frequencies were analyzed by means of Chi-square test using sing the co-dominant model. A Logistic regression test was done to determine the association between different cytokine gen polymorphism and tuberculosis infection. Odds ratios (ORs) with 9595% CIs were also considered. A P value < 0.05 was considered statistically substantial for all tests.
Result
A total of 200 age and sex-matched study subjects, 100 cases and 100 controls, were enrolled in this finding. Among those 58% (58/100) in cases and 57% (57/100) in controls were male. The mean age was 34.9
The Socio-Demographic Characteristics of Study Groups.

Association of TNF-α -308 G/A Polymorphism and Susceptibility to TB
The frequency of TNF-α -308 A (wild type) to G (mutant type) in TB patients and healthy group illustrated in Table 3. The frequency of TNF-α allele G in TB patients and control groups were 77% and 48.5% ((P = 0.00, OR (95% CI) =0.20 (0.11-0.37)), respectively. In addition, the GG genotype frequency was 64% and 20% in TB patients and control groups (P
Relationship Between TNF-α -308 Locus Polymorphisms and the Odds of Developing TB, 2018.

Note: TB: Tuberculosis; N: Number; OR: Odd<b>s</b> ratio, *P significant < 0.05
Association Between IFN γ+874 T/A and Susceptibility to TB
The frequency of IFN-γ +874 A allele was higher in TB patients than healthy controls, which were 62% and 47.5%, respectively. While the AA genotype (mutant) was higher in TB patients (41%) compared to healthy controls 25% ((P < 0.001, OR (95% CI)= 3.80 (2.11-6.86)), Therefore, this result describes that IFN-γ +874 A allele and AA genotype (mutant) has highly associated with TB infection than T allele (38%) and TT genotype (wild) (27%) ((P < 0.008, OR (95% CI)= 1.61(1.13-2.28)) (Table 4).
The Relationship Between IFN-γ +874 Gene Variation and the Odds of Developing TB, 2018.
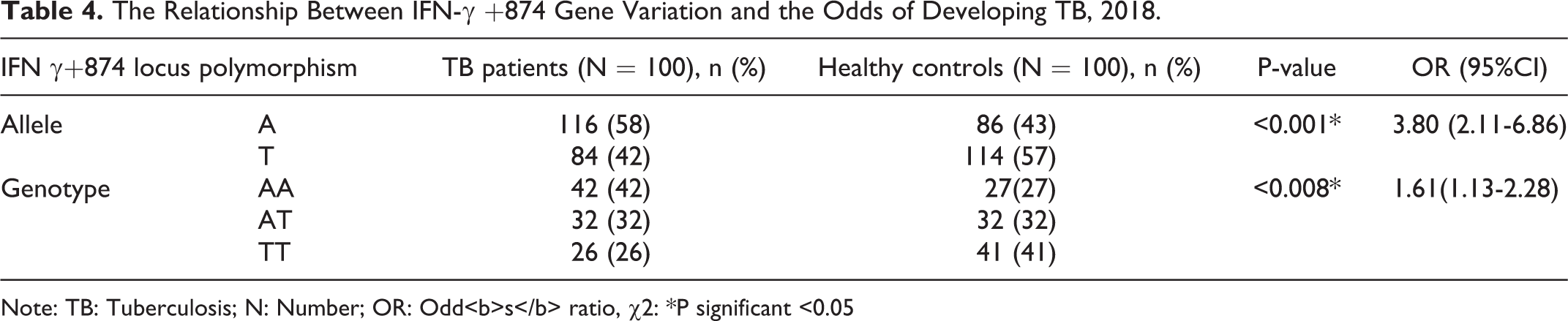
Note: TB: Tuberculosis; N: Number; OR: Odd<b>s</b> ratio, χ2: *P significant <0.05
IL-10 -1082 G/A Associated with TB Susceptibility
There is no significance difference in frequency of A allele/ AA genotype and G allele/GG genotype of IL-10 at -1082 locus between healthy controls and TB patients (P = 0.20, OR (95% CI)= 0.70 (0.40 -1.22)), Thus, IL-10 -1082 Aallele and AA genotype (mutant) has no significant association with TB infection ((P = 0.05, OR (95% CI)= 0.72 (0.52-0.99) (Table 5).
Relationship between IL-10 -1082 Locus Polymorphism and Odds of Developing TB, 2018.

Note: TB: Tuberculosis; N: Number; OR: Odd<b>s</b> ratio
Discussion
Host genetics remain the main players of susceptibility and/or resistance to many communicable diseases including TB. Thus, TB infection risk was associated with several candidate gene polymorphisms. Moreover, the genes encoding for the regulatory and Th1 signature mediators may play vital roles in host susceptibility to TB, as the interplay between these mediators might determine the immunological status of individuals in question. The interaction of the cytokine gene variation with TB susceptibility was explored in different ethnic groups and came up with different results. Therefore, in the era of personalized medicine, understanding the genetic polymorphism of the cytokines of our interest in TB patients that has a different genetic makeup than Europeans and Asians is paramount importance.
In this study, we found that TNF-α allele G and genotype GG frequency was higher in TB patients than the control group, respectively (P = 0.00 and P
In addition, the existence of A allele and AA genotype (mutant type) in IFN-γ was higher in TB patients than control groups (P < 0.001, and P < 0.008), respectively. It is consistent with the finding from China 30,31 , Spain 32 , Iran 17,33 , Hong Kong, 30 India, Han Taiwan, and Pakistan. 3,34,35 This may be due to the A allele and AA genotype at +874 position corresponds to lower IFN-γ expression, macrophage activation, and cellular proliferation and apoptosis. Therefore, a lower IFN-γ expression can impair macrophage activation, which limits the production of ROIs and RNIs result in favorable conditions for the progression of TB disease. However; the study conducted in south India, Houston and western Africa (Ghana) indicated that there was no significant difference between TB patients and controls in A allele and AA genotype frequency. 36,37 This might be partly attributed to the genetic variation of the society in these countries; the other probable reason for the disparity might be the sample size difference between the different studies.
In addition, the frequency of A allele and AA genotype (mutant type) in IL-10 was also assessed. Thus, the finding of this research revealed that a high frequency of A allele and AA genotype in healthy controls than TB patients (P = 0.20 and P = 0.05, respectively). This result is inconsistent with a study conducted in Iran (Azari population) and Turkey (p = 0.001). However, allele G and genotype GG frequency was higher in TB patients 56.5% and 44.5%, respectively, than healthy controls (42% vs 33%, respectively), in agreement with study in Iran and Turkey. 31,38 This may be due to A allele and AA genotype at -1082 positions has a minor effect in down-regulating Th1 driving pro-inflammatory cytokines, and less interruption of host immune response. Subsequently, pro-inflammatory cytokine augments the immune response against TB. In contrast, the G allele and GG genotype (wild type) at -1082 position leads to decrease Th1 driving cytokine, macrophage deactivation and preventing the release of ROIs/RNIs. In consequence, it is favorable to the development of TB. In contrast, our finding is different with a study done in Korea, A allele was significantly related with TB patients than controls (P = 0.001). 39
Conclusion
Our result shows that TNF-α -308 G/A and IFN-γ +874 A/T genetic polymorphisms in TB patients demonstrate higher frequencies of GG and AA genotype, respectively as compared to healthy controls. It implies that TNF-α and IFN-γ gene polymorphisms are related to the risk of TB infection. However, the frequency of AA genotype was lower in TB patients than healthy controls, though, it was not clearly showing the protective roles to TB infection.
Footnotes
Abbreviations
Acknowledgment
We would like to express our deepest and heartfelt gratitude to department of Immunology and Molecular Biology, School of Biomedical and Laboratory Science, College of Medicine and Health Science, University of Gondar to give the opportunity to do this project. Our acknowledgment also goes to the institute of Biotechnology, University of Gondar particularly to Professor Nega Berhane and Laboratory assistance who helps us to conduct the laboratory work. Finally, we would like to thank the University of Gondar Comprehensive Specializes Hospital and study participants for their collaboration during sample collection.
Author Contribution
GA generates the research idea, develop the research protocol and draft the manuscript. GA, BA, TS did laboratory activities. GA, DD, DG, ML, TS, MKT and BA were involved in the research protocol development, and critically reviewed the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethics Approval and Consent to Participate
The study was approved by the School of Biomedical and laboratory Science, University of Gondar (SBMLS892/10) ethical review committee. Each study participant was informed about the objective of the study and written informed consent was obtained from each participant.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was supported by the University of Gondar, Gondar, Ethiopia. These funds were used to collect data and purchase laboratory material and reagents.