
Abstract
Metabolic pathways are often spread over several organelles and need to be functionally integrated by controlled organelle communication. Physical organelle contact-sites have emerged as critical hubs in the regulation of cellular metabolism, but the molecular understanding of mechanisms that mediate formation or regulation of organelle interfaces was until recently relatively limited. Mitochondria are central organelles in anabolic and catabolic pathways and therefore interact with a number of other cellular compartments including the endoplasmic reticulum (ER) and lipid droplets (LDs). An interesting set of recent work has shed new light on the molecular basis forming these contact sites. This brief overview describes the discovery of unanticipated functions of contact sites between the ER, mitochondria and LDs in de novo synthesis of storage lipids of brown and white adipocytes. Interestingly, the factors involved in mediating the interaction between these organelles are subject to unexpected modes of regulation through newly uncovered Phospho-FFAT motifs. These results suggest dynamic regulation of contact sites between organelles and indicate that spatial organization of organelles within the cell contributes to the control of metabolism.
Keywords
The separation of biochemical reactions into organelles with specific function is a central principle of cellular organization. However, many cell biological and metabolic pathways use activities that are spread over several organelles, and a controlled integration of compartmentalized activities as well as the regulated interplay of metabolic pathways across organelles is essential for life (Bohnert, 2020).
While most membrane bound organelles such as the endoplasmic reticulum (ER), the Golgi apparatus, and the endocytic compartments are connected by vesicular membrane traffic, not all organelles possess the machinery to generate and receive vesicular transport carriers. Instead these organelles are linked by non-vesicular transport pathways and physical organelle contact sites (Figure 1(a)) (Holthuis and Levine, 2005; Barbosa et al., 2015; Murley and Nunnari, 2016; Gatta and Levine, 2017).
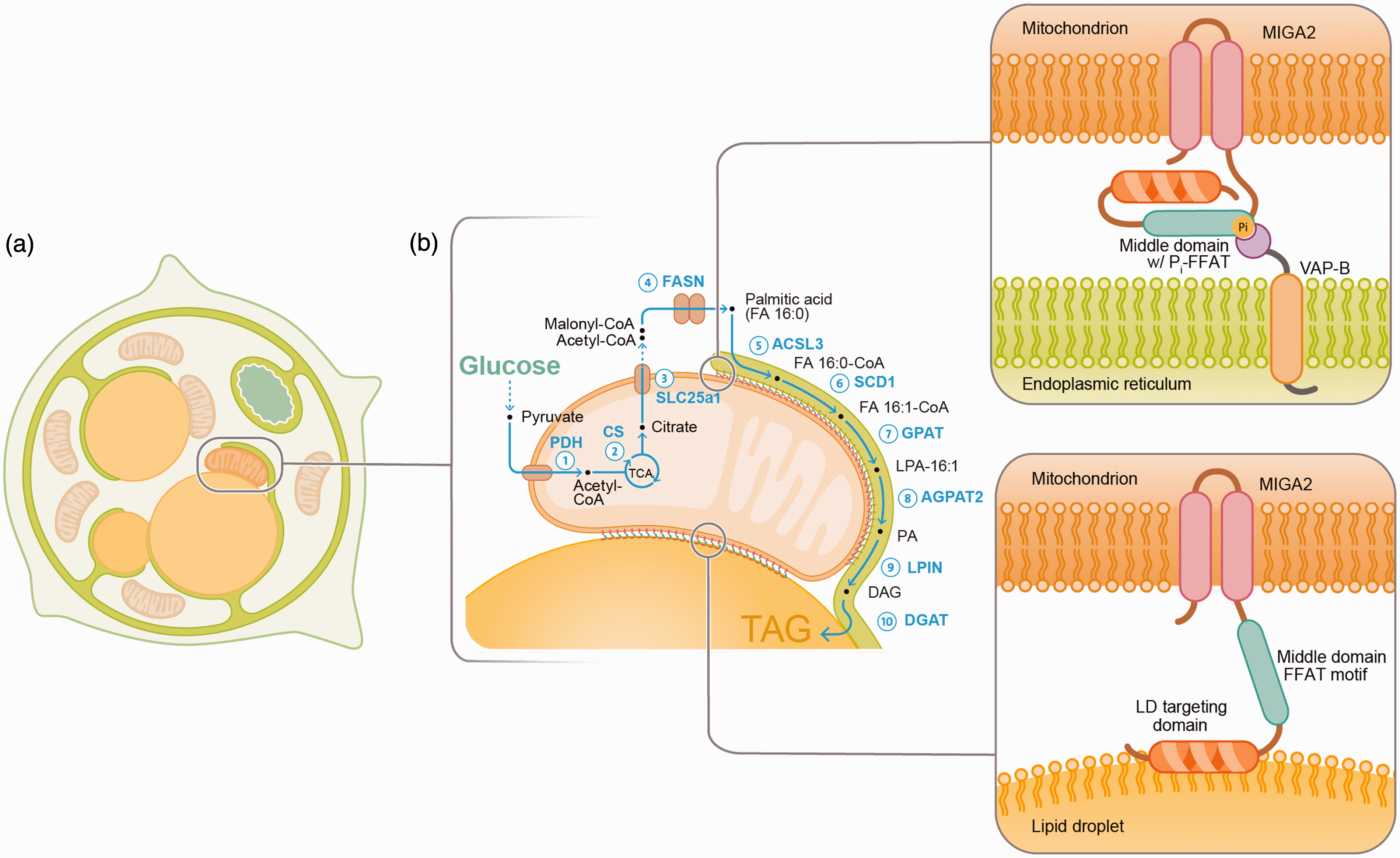
Schematic depiction of organelle contact-sites between the ER (green) mitochondria (brown), and lipid droplets (LDs, yellow) and their role in de novo lipogenesis in differentiating white adipocytes (adopted from Freyre et al., 2019). (a) Adipocyte differentiation is controlled by a transcriptional program that induces structural and biochemical changes to enlarge LDs for lipid storage. The LD is connected with most organelles during differentiation and unexpectedly interfaces with mitochondria. (b) Apart from storing dietary derived fatty acids in LDs, adipocytes convert non-lipid precursors such as glucose into fatty acids, a process called de novo lipogenesis. MIGA2 links mitochondria to the ER by binding VAP proteins through a Phospho-FFAT motif (yellow) in the middle domain (magenta). LD binding is mediated by a C-terminal amphipathic segment in MIGA2, shown in in orange. MIGA2 promotes de novo lipogenesis and the synthesis of triacylglycerols (TAGs). The different steps of the de novo lipogenesis and TAG synthesis pathway are indicated in blue and the pathway intermediates are shown in black. Glucose is converted into pyruvate which is transported into mitochondria, and decarboxylated by the pyruvate dehydrogenase (PDH, step 1) to produce acetyl-CoA which is condensed with oxaloacetate into citrate by the citrate synthase (CS, step 2). Citrate is exported into the cytoplasm by the mitochondrial citrate exporter (SLC25a1, step 3). In the cytoplasm citrate is cleaved into oxaloacetate and acetyl-CoA, which is then carboxylated into malonyl-CoA the major substrate for the production of palmitic acid (FA16:0) by the fatty acid synthase (FASN, step 4). In the ER the acyl-CoA synthetase-3 (ACSL3) activates FA16:0 to FA16:0-CoA (step 5), which can be desaturated by the stearyl-CoA desaturase (SCD1) to produce FA16:1-CoA (step 6). Two FA-CoAs are then esterified to a glycerol first by the glycerol-3-phosphate-acyltransferase-3 (GPAT3) which produces lysophosphatidic acid (LPA, step 7) and second by the 1-acyl-sn-glycerol-3-phosphate acyltransferase-2 (AGPAT2, step 8) which synthesizes phosphatidic acid (PA). Lipin (LPIN) dephosphorylates PA to produce diacylglycerol (DAG, step 9), the direct precursor of triacylglycerol (TAG) which is produced by the activity of the diacylglycerol-O-acyltransferases (DGAT1 and 2, step 10).
Organelle contact sites carry out the exchange of lipids, signalling molecules or metabolic intermediates, and are therefore observed between most, if not all organelles in the cell, even if they are linked by vesicular traffic (Henne et al., 2015; Wu et al., 2018; Henne, 2019; Antonny et al., 2018).
The molecular basis of organelle associations and elucidating the mechanisms of organelle communication at contact sites have given rise to a vibrant research field, which is currently laying out a new molecular framework of organelle contact site biology (Grippa et al., 2015; Prinz, 2014; Henne et al., 2015; Murley and Nunnari, 2016; Schrepfer and Scorrano, 2016; Barbosa et al., 2015; Eisenberg-Bord et al., 2016; Saheki and Camilli, 2017; Bohnert, 2019, 2020; Cao and Mak, 2020; Harper et al., 2020; Wang et al., 2018). Among the wealth of novel insights, a central conceptual advance is that many more organelle tethering complexes exist than initially anticipated (Kornmann et al., 2009; Giordano et al., 2013; Kornmann, 2013; Dong et al., 2016; Kumar et al., 2018). The fact that multiple contact sites are observed even between the same organelles, and that these contacts are formed by different protein-complexes, indicate that each contact site may have specific functions (Wu et al., 2018). While organelle tethering is obviously an essential activity shared by all organelle contact site complexes, the new insights suggest that merely establishing spatial proximity is not a specific function but a more general requirement to form organelle interfaces. The rapidly increasing number of factors linking cellular compartments, as well as a new understanding of the regulatory mechanisms that control the spatial organization and function of contact sites is opening up a new perspective on organelle biology, and many more important discoveries are expected in this burgeoning field.
New Insights Into Mechanisms That Link Mitochondria With the ER and LDs
A recent study by Freyre et al. (Freyre et al., 2019) has identified the molecular basis of an organelle interface that is formed between three organelles including mitochondria, lipid droplets (LDs) as well as the ER in differentiating white adipocytes (Figure 1(a)). Using mass spectrometry proteomics after isolation of LDs from differentiating adipocytes, combined with expression localization screening of a number of candidate proteins, Freyre et al. uncovered the outer mitochondrial membrane protein mitoguardin 2 (MIGA2) (Zhang et al., 2016) as a factor involved in linking mitochondria to LDs (Figure 1(b)) (Freyre et al., 2019). The structural features of MIGA2 with N-terminal mitochondrial membrane anchors and several amphipathic segments at the C-terminus suggested that MIGA2 might directly mediate the formation of the mitochondria - LD interface (Figure 1(b)). Extensive structure function analyses with a set of truncation mutants and domain-swap constructs revealed a C-terminal amphipathic segment necessary and sufficient for LD interaction. The deletion of a single amphipathic segment between residues 489 and 512 or the introduction of charged aspartates in the hydrophobic face of this segment prevented LD binding (Figure 1(b)) (Freyre et al., 2019). Strikingly, moving a portion of MIGA2 containing this amphipathic segment to the C-term of the ER membrane protein calnexin led to efficient lipid droplet targeting. Despite being sufficient in targeting membrane proteins to the LD, the C-terminal portion of MIGA2 alone, without any membrane anchor was not able to localize to the LD surface, but remained largely cytoplasmic.
A Functional Role for Organelle Contact Sites in Metabolic Control
A functional analysis by CRISPR-Cas9 knockout and overexpression experiments in differentiating adipocytes resulted in the surprising finding that docking of mitochondria onto LDs does not lead to the expected consumption of the LDs. Instead, MIGA2 promoted the synthesis of triacylglycerols (TAGs), which are the storage lipids deposited in LDs of adipocytes. The simplest explanation for these data was that adipocyte mitochondria are predominantly anabolic and do not oxidize fatty acids. This hypothesis was corroborated by results in the literature showing that block of fatty acid import into mitochondria by pharmacological inhibition of the carnitine-palmitoyl transferase does not dramatically interfere with adipocyte function (Gao et al., 2011). The biosynthetic role of MIGA2 was extensively tested by radioisotope tracing experiments and quantitative mass spectrometry based lipidomics showing that MIGA2 enhances de novo lipogenesis by increasing the supply of non-lipid derived precursors for the fatty acid synthase leading to a drastic increase in TAGs (Figure 1(b)).
In brown adipocytes Benador et. al. have recently shown that LD binding can lead to the formation of a specific sub-population of mitochondria, characterized by a specific metabolic signature and termed them “peridroplet mitochondria” (Benador et al., 2018). Consistent with the model by Freyre et al., peridroplet mitochondria do not specialize in oxidizing fatty acids, but are involved in the synthesis of TAGs. This result is perhaps even more surprising than in white adipocytes, because brown adipocytes are known to efficiently consume fatty acids mobilized from TAGs for thermogenesis. Direct physical contact between mitochondria and LDs was so far considered critical for efficient oxidation of TAGs. The careful metabolic analysis by Benador et al. revealed, however, that oxidation reactions in mitochondria that co-purified with LDs are not uncoupled from ATP synthesis. The higher oxidative capacity, and enhanced flux through the tricarboxylic acid (TCA) cycle of LD-bound mitochondria rather led to higher levels of ATP production. Unexpectedly, the substrates supplying energy for ATP synthesis were pyruvate, malate, and succinate instead of free fatty acids. According to the authors, augmented synthesis of ATP improved the ATP dependent activation of dietary fatty acids (FAs) by attaching coenzyme A (CoA). A high level of FA-CoAs was in turn necessary to increase the esterification with diacylglycerols (DAGs) in order to produce TAGs, confirming an anabolic function of mitochondria that interact with LDs (Benador et al., 2019).
Given that the higher flux through the TCA cycle also led to an increase in citrate production as shown by Benador et al., it is conceivable that next to increasing ATP production, binding of mitochondria to the LD surface might additionally stimulate the synthesis of FAs as suggested in the model by Freyre at al. After export from mitochondria, citrate is cleaved into oxaloacetate and acetyl-CoA, and subsequently carboxylated to form malonyl-CoA, which is then used as the substrate for de novo synthesis of FAs by the fatty acid synthase (Figure 1(b); Lunt and Heiden, 2011; Freyre et al., 2019; Wallace and Metallo, 2020). Consistent with this interpretation Benador et al. report that cells with an increased amount of LD-bound mitochondria have higher levels of NADH or NADPH which is a critical co-factor in the FA-elongation reactions by the fatty acid synthase. A simultaneous increase of NADPH, citrate, and enhanced ATP synthesis could have synergistic effects on both the synthesis of TAGs from de novo generated FAs as shown by Freyre at al. (Figure 1(b)) and from exogenous fatty acids as demonstrated by Benador et al.
Despite these overlapping aspects, the molecular basis of mitochondrial LD-binding appears to be different between white and brown adipocytes. Strikingly, the transcriptome analysis by Freyre et al. showed that perilipin 5 (PLIN5), which is involved in linking mitochondria and LDs in brown adipocytes, is not detectable in white adipocytes. The data by Freyre et al. further do not provide evidence for distinct populations of mitochondria as seen in brown adipocytes. In fact, MIGA2 was found on both the cytoplasmic and LD-bound mitochondria. Whether MIGA2 is present in the LD bound mitochondria population of brown adipocytes, and whether MIGA2 in turn increases ATP production in white adipocytes and simultaneously stimulates TAG synthesis with de novo synthesized, as well as dietary FAs remains to be answered.
FFAT-Motif Dependent VAP Binding Forms Contact Between the ER and Mitochondria
Apart from the obvious difference in physiological function of brown and white adipocytes the mechanistic differences observed in LD-mitochondria interfaces of these two cell types might further be explained by the ability of MIGA2 to additionally bind the ER (Figure 1(b)) (Hurtley, 2019). Freyre et al. discovered an amino acid sequence within the middle domain of MIGA2, which is critical for the interaction with the ER. The segment belongs to the evolutionarily conserved family of peptide sequences called “two phenylalanines in an acidic tract” or FFAT motifs (Loewen et al., 2003; Loewen and Levine, 2005; Murphy and Levine, 2016), which are known to interact with VAMP associated proteins (VAP), a family of tail anchored ER membrane proteins. Using ectopic expression in COS7 cells and a stably integrated trans-gene for inducible overexpression in pre-adipocytes, MIGA2 was shown to bind endogenously expressed as well as over-expressed VAP-A and VAP-B. The interaction between the two proteins required both a FFAT motif in MIGA2 and a functional FFAT peptide binding domain in VAPs (Figure 1(b), upper zoom).
Regulation of FFAT Motifs by Phosphorylation
A question that was left open by Freyre et al. pertained to the origin of several MIGA2 species visible in western blotting experiments. This aspect was recently clarified in work by Xu and Wang et al. focusing on mitoguardin (Miga) in fly neurons (Zhang et al., 2016). Xu and Wang et al. revealed that the FFAT motif in fly Miga is activated by phosphorylation. A first indication for regulation of FFAT motifs by phosphorylation was already obtained in earlier work by Mikitova et al. (Mikitova and Levine, 2012). Indeed, some of the phosphoserines in Miga are found directly within the FFAT motif itself and their phospho-status proved critical for the interaction with fly Vap33, an ortholog of the mammalian VAPs.
Importantly, Di Mattia, Martinet, and Ikhlef et al. have recently shown that the FFAT motif in MIGA2 belongs to an entirely new subclass of non-conventional FFAT motifs called Phospho-FFAT motifs (Di Mattia et al., 2020). Bioinformatic analyses predicted more than 100 candidates in the human proteome possibly containing Phospho-FFAT motifs, with MIGA2 topping the list (Di Mattia et al., 2020). The extensive biochemical and structural validation of the interaction between Phospho-FFAT motifs and the Major Sperm Protein (MSP) domains in VAP and MOSPD2 proteins suggest a completely new regulatory function of FFAT motifs by phosphorylation. Since the activation of the FFAT motif in MIGA2 by phosphorylation is critical for the interaction with MSP domains, these findings provide evidence for an exciting novel mechanism that is likely used to dynamically regulate the formation of organelle interfaces.
The study by Xu and Wang et al. further suggested that phosphorylation of Miga in fly retina might occur as part of a starvation response possibly involving casein kinase I and calmodulin/Ca2+ dependent protein kinase II. Given that the activation of FFAT motifs is likely reversible by an as yet unknown phosphatase, this mode of regulation might dynamically control the formation and release of organelle contact sites perhaps in response to metabolic requirements within cells.
Consistent with these data the hormonal cocktail that is used to initiate differentiation of adipocytes in tissue culture induces a strong starvation-like signal by inhibiting cAMP/cGMP cyclic phosphodiesterases leading to activation of protein kinase A (PKA). PKA might be an upstream factor in the signalling cascade activating the Phospho-FFAT motifs in MIGA2. Interestingly, the PKA anchoring protein AKAP220 was among the first proteins shown to have phosphorylated FFAT motifs (Mikitova and Levine, 2012), and binds to the ER via interaction with VAPs. It might be the case that phosphorylation of client FFAT motifs can only occur when both the relevant kinases and substrates such as MIGA2 are recruited close to the ER.
Since it is important that the FFAT motifs are sterically accessible to the activating kinases, it is tempting to speculate that binding of the C-terminus of MIGA2 to LDs has a function in controlling the exposure of FFAT motifs in the middle domain (Figure 1(b), zoom). Conversely, binding to VAPs might stabilize MIGA2 in a particular conformation and either positively influence or restrict exposure of the LD targeting segment. The postulated open and closed conformations that are indicated in Figure 1(b) might be intermediates in the dynamic life cycle of the MIGA2 contact site.
These possible concepts of regulation pose multiple questions, such as when exactly ER binding of MIGA2 occurs during adipocyte differentiation and how it supports anabolic metabolism during TAG synthesis. Should the interaction with VAPs on the ER rather restrict access to the LDs, for instance during starvation, it would be expected that binding to the ER might negatively regulate de novo lipogenesis possibly by moving MIGA2 away from the LD surface and attenuate TAG production temporarily. Future experiments with the large catalogue of available MIGA2 mutants may shed light on these issues.
Possible Functions of MIGA2 at Organelle Contact Sites
The exact function of MIGA2-mediated organelle interactions, or even more generally speaking the cell biological role of contact sites between mitochondria and LDs or the ER is not completely understood in mechanistic terms. While previous work from several laboratories focusing on the formation of contact sites between mitochondria and LDs in cells undergoing starvation has suggested a role for mitochondria bound to LDs in oxidizing lipids (Herms et al., 2015; Rambold et al., 2015; Nguyen et al., 2017), which would be most consistent with a function in lipid transport at these interfaces, it is not yet clear whether MIGA2 or PLIN5 play a role in this context. Given that a number of lipid transfer proteins contain classical (Loewen et al., 2003; Kumagai et al., 2014) or Phospho-FFAT motifs (Di Mattia et al., 2020) it is an interesting possibility that MIGA2 might participate in lipid transport between mitochondria and the ER or the LDs. For instance, MIGA2 might be directly necessary for an alternative mitochondrial fatty acid import pathway or might transport fatty acids instead to mitochondrial machinery or factors on the ER for fatty acid activation by CoA synthetases.
A last possible function for MIGA2 at organelle contact sites might be related to scaffolding. Bringing several metabolic factors into close spatial proximity, either across the organelle interface or within the same organelle might be beneficial for efficient substrate flux, or regulation of pathway activity. Regulation of contact site components by clustering into functional platforms has recently been shown for a contact site between VAP proteins in the ER and members of the Kv2 family of voltage gated potassium channels at the neuronal plasma membrane (Kirmiz et al., 2018). Importantly, clustering was dependent on the activation of the Phospho-FFAT motif in the C-terminus of Kv2.1 which led to binding of VAPs in the ER and seemed to downregulate ion conductivity of Kv2.1 (Johnson et al., 2018). Instead, the clustered channels were suggested to take over functions in membrane traffic, possibly acting as a docking sites for vesicles. Interestingly, the localization to clusters was rapidly reversed when the neuronal activity was stimulated leading to the rapid dispersion of the channels within the plasma membrane which in turn increased ion conductivity. Similar structural functions, perhaps in organizing components of metabolic pathways are possible for MIGA2 dependent membrane domains within mitochondria and through VAP clusters at the ER.
How the mitoguardins relate to other factors forming contact between the ER, mitochondria and lipid droplets such as members of the VPS13 family (Bean et al., 2018; Kumar et al., 2018), RMDN3 (PTPIP51) (Vos et al., 2012; Stoica et al., 2014), or ORP5 and 8 (Galmes et al., 2016; Du et al., 2020), and whether these different contact sites synergize with one another to integrate multiple metabolic pathways across organelles as seen for various organelle interfaces in budding yeast (Elbaz-Alon et al., 2014; Ungermann, 2015; John Peter et al., 2017) will have to be worked out. Interestingly, according to Di Mattia, Martinet, and Ikhlef et al. VPS13C and RMDN3 are also candidates that might contain Phospho-FFAT motifs, which indicates that metabolic phosphoregulation of contact sites between mitochondria, the ER, and lipid droplets might be a more general mechanism important in controlling lipid metabolism at organelle interfaces.
Another interesting avenue of investigation is how the ER, mitochondria, and LDs communicate with the lysosomal compartments and autophagosomes. Recent evidence points at a function of the ER protein PDZD8 in bringing late endosomes and lysosomes close to these organelles, a function that might be of particular importance in the context of metabolic homeostasis during nutrient stress (Hirabayashi et al., 2017; Guillén-Samander et al., 2019; Elbaz-Alon et al., 2020; Shirane et al., 2020). Given that starvation-induced autophagy, also depends on physical contact sites, in this case between autophagosomes, LDs and mitochondria (Herms et al., 2015; Rambold et al., 2015; Nguyen et al., 2017), the evidence for a more general role of organelle interfaces in regulating metabolism and cellular differentiation is steadily increasing, and we will soon need to extensively update our text book models of mechanisms controlling organelle function. Most excitingly, metabolism appears to be controlled by the spatial organization of pathways across different organelles, and the concerted interplay between cellular compartments appears to be regulated by signalling processes possibly relaying metabolic information to contact site complexes.
Footnotes
Acknowledgements
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by University of Oxford