Addressing Compound Reactivity and Aggregation Assay Interferences: Case Studies of Biochemical High-Throughput Screening Campaigns Benefiting from the National Institutes of Health Assay Guidance Manual Guidelines
Free accessResearch articleFirst published online December, 2021
Addressing Compound Reactivity and Aggregation Assay Interferences: Case Studies of Biochemical High-Throughput Screening Campaigns Benefiting from the National Institutes of Health Assay Guidance Manual Guidelines
Compound-dependent assay interferences represent a continued burden in drug and chemical probe discovery. The open-source National Institutes of Health/National Center for Advancing Translational Sciences (NIH/NCATS) Assay Guidance Manual (AGM) established an “Assay Artifacts and Interferences” section to address different sources of artifacts and interferences in biological assays. In addition to the frequent introduction of new chapters in this important topic area, older chapters are periodically updated by experts from academia, industry, and government to include new technologies and practices. Section chapters describe many best practices for mitigating and identifying compound-dependent assay interferences. Using two previously reported biochemical high-throughput screening campaigns for small-molecule inhibitors of the epigenetic targets Rtt109 and NSD2, the authors review best practices and direct readers to high-yield resources in the AGM and elsewhere for the mitigation and identification of compound-dependent reactivity and aggregation assay interferences.
High-throughput screening (HTS) is a key fixture in many modern drug and chemical probe discovery campaigns. However, even the most robust and well-executed HTS assays inevitably identify interference compounds among the apparently active screening hits.1 Left unchecked, compound-dependent assay interferences waste substantial scientific resources, lead to flawed scientific conclusions, and are propagated in the scientific literature.2
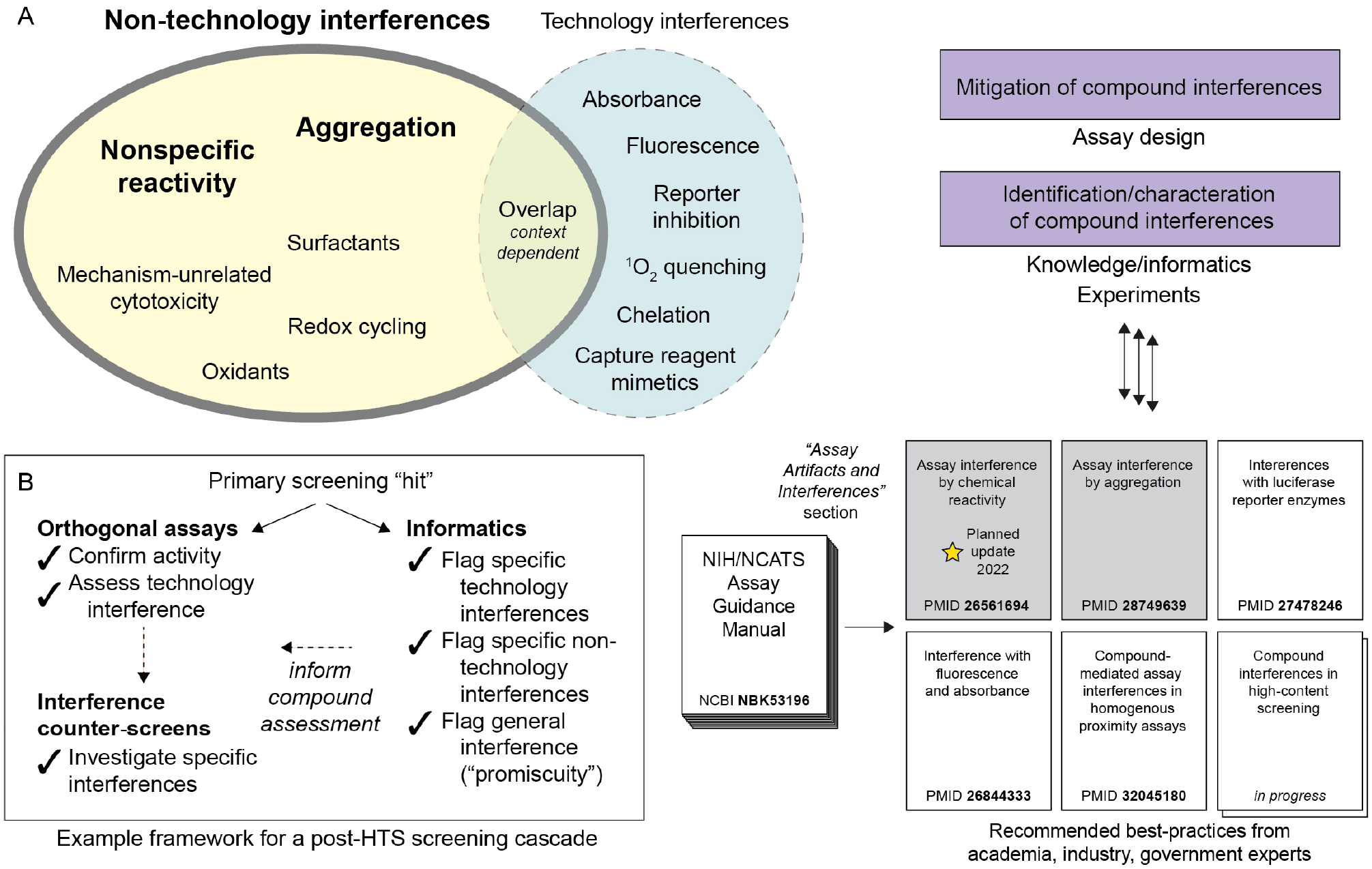
Compound-dependent assay interferences can be divided into two broad but overlapping categories: technology-related and non-technology-related interferences (Fig. 1A). Compounds can fail to modulate the intended biological target but interfere with the core assay technology to produce apparent bioactivity by a variety of mechanisms (artifacts), including optical interferences (e.g., absorbance, fluorescence, quenching, light scattering), singlet oxygen quenching (e.g., AlphaScreen), capture reagent disruption (e.g., streptavidin-biotin, Ni-His, GSH-GST tags), modulation of a reporter enzyme (e.g., luciferase), and others.3–5 By contrast, interference compounds can also genuinely modulate the target bioactivity of interest but through undesirable and intractable mechanisms of action (MOAs) that are not amenable to improvements in potency and selectivity toward a specific target, such as nonspecific chemical reactivity, colloidal aggregation, and mechanism-unrelated cytotoxicity.6,7 In certain cases, compounds can interfere with both the biology of interest and the assay technology, often through nonspecific reactivity or chelation. Furthermore, compound stock solutions can contain impurities, including degradation products, metals, and remnants of reactive reagents used in the compound synthesis that interfere with assays or cause nonspecific activity.8–11 Compound interferences, including nonspecific reactivity and aggregation, can adversely affect both biochemical (cell-free) and cellular (including phenotypic and high-content assays) assays.12
Summary of compound-dependent assay interferences. (A) Compound-dependent assay interferences can be broadly divided into non-technology-related and technology-related categories, although overlap can exist. These interferences are best mitigated through a combination of assay design strategies and a robust post-HTS cascade consisting of both experimental and knowledge-based approaches. To assist scientists with this important task, the open-source AGM maintains an expanding and frequently updated section on assay artifacts and interferences. (B) An example framework for addressing compound interferences among hits from a biochemical HTS. Orthogonal assays are helpful for confirming activity and de-risking technology-related interferences, although compounds with non-technology-related interference mechanisms might still appear active even with orthogonal assay technologies. Counterscreens for common non-technology-related interferences like nonspecific reactivity and aggregation are therefore recommended. Informatics tools, such as substructure filters and promiscuity scoring, are helpful approaches to support compound prioritization decisions.
In order to counter this significant threat to scientific productivity and integrity, the National Center for Advancing Translational Sciences (NCATS) at the U.S. National Institutes of Health (NIH) created the “Assay Artifacts and Interferences” section within the Assay Guidance Manual (AGM) (Fig. 1, right).13 The goals of this section are to summarize the fundamental mechanisms of various assay interferences, suggest strategies for mitigating compound-dependent interferences during the assay design phase, and provide guidance for designing a “testing funnel” of assays to identify and de-risk compound-dependent interferences from apparently active compounds arising from these bioassays.
In this Perspective, we utilize two previously published HTS campaigns as model case studies to walk through some best practices for identifying nonspecific reactivity and aggregation interferences in biochemical (cell-free) assays. These practices are detailed in the AGM, and readers will be directed to specific high-yield resources in the AGM and elsewhere. For each case study, readers will primarily learn about (1) experimental approaches to triage interference compounds regardless of the mechanism (e.g., orthogonal assays), (2) methods to investigate non-technology-related compound interferences (e.g., nonspecific thiol reactivity, aggregation), and (3) supporting informatics approaches (e.g., promiscuity analyses, substructure filters) (Fig. 1B).
Case Study 1: Nonspecific Reactive Compounds
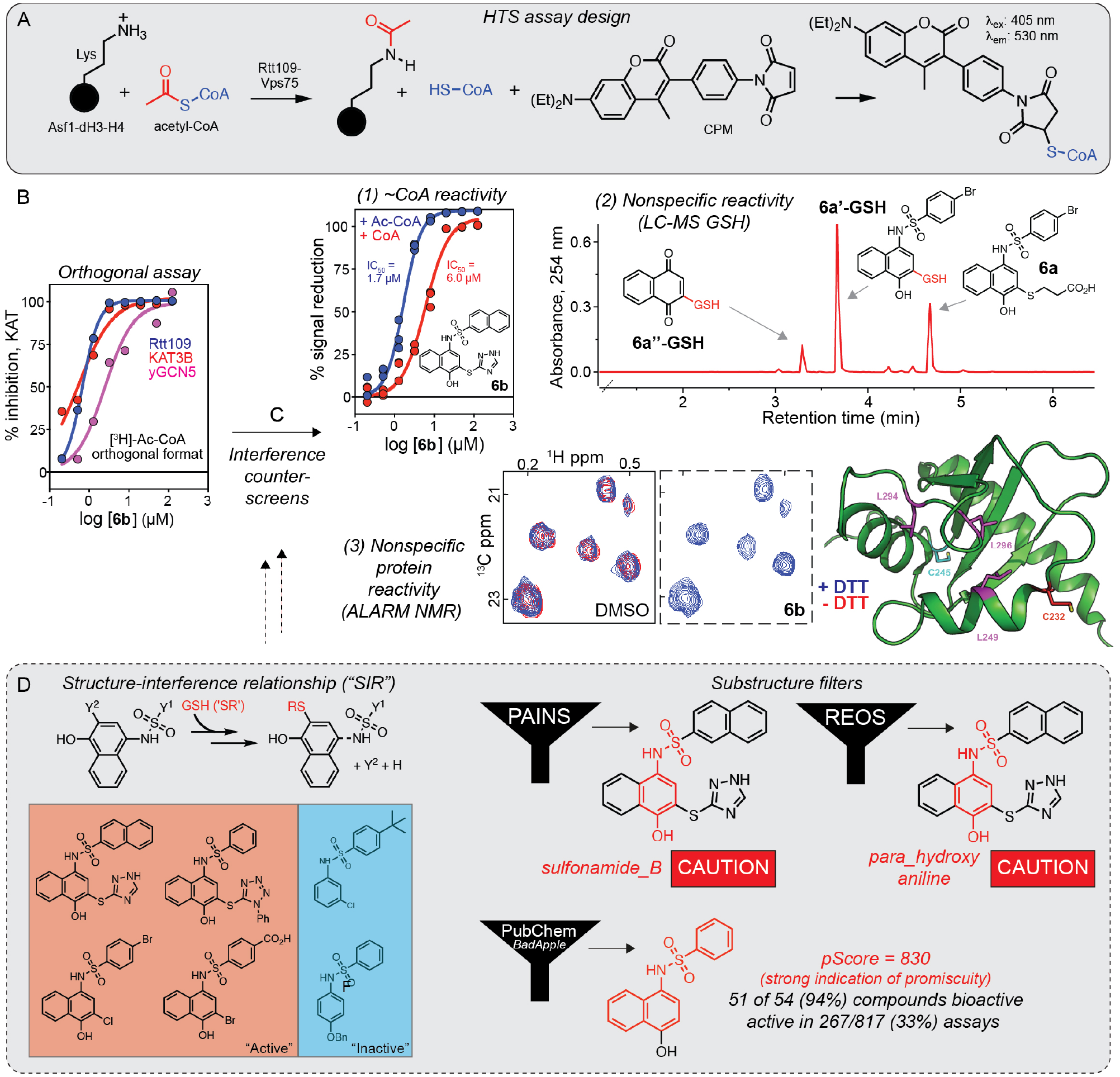
A cell-free fluorometric assay was developed to identify small-molecule inhibitors of the Saccharomyces cerevisiae Rtt109 lysine acetyltransferase (KAT) as potential antifungal agents (Fig. 2A).14 In this assay, the Rtt109-Vps75 KAT complex catalyzes the transfer of an acetyl moiety from acetyl–coenzyme A (Ac-CoA) to the Asf1-dH3-H4 histone–chaperone substrate to produce acetylated histones and CoA. The Rtt109 activity can then be indirectly quantified by measuring the resulting CoA using a thiol-reactive probe, 7-diethylamino-3-(4-maleimidophenyl)-4-methylcoumarin (CPM), which rapidly reacts with the free thiolate on CoA to produce a highly fluorescent CPM-CoA adduct. The assay was sufficiently robust (Z′ = ~0.7) and could identify reported KAT inhibitors such as garcinol. A screen of 225,000 small molecules identified active compounds, such as 6b, with a particular para-hydroxy-arylsulfonamide chemotype 6, as inducing reproducible, low micromolar inhibition of Rtt109. A [3H]-Ac-CoA filter-binding orthogonal assay confirmed this low micromolar activity, demonstrating that the observed primary HTS activity was due, at least in part, to Rtt109 inhibition (Fig. 2B). Because non-technology-related interferences can cause on-target activity independent of assay technology, the AGM recommends performing at least one counterscreen for the most common nonspecific interferences, such as nonspecific thiol reactivity.7
Case study of nonspecific reactive compound identification from a high-throughput screen. (A) Schematic of the primary HTS assay for Rtt109-catalyzed histone acetylation. (B) Activity of the representative para-hydroxy-arylsulfonamide 6b can be confirmed by an orthogonal [3H]-Ac-CoA filter-binding assay. Note that 6b is not selective versus the related targets KAT3B and yeast GCN5. (C) Compounds active by an orthogonal assay should be assessed for non-technology-related interferences. The compound 6b and related analogs were flagged for nonspecific reactivity using (1) a counterscreen of the primary HTS that replaced the Ac-CoA substrate (blue) with the CoA product (red), (2) an LC-MS counterscreen for compound instability and compound-thiol adducts under assay-like conditions (n.b., analog 6a shown), and (3) an ALARM NMR counterscreen for nonspecific protein reactivity. (D) Informatics supported the decision to triage chemotype 6: substructure filter results (PAINS, REOS) and historical screening performance (PubChem) were consistent with a nonspecific hit, while the apparent SAR was best explained by reactivity-based SIRs. Portions of this figure were adapted from a previous publication.15
A counterscreen based on the primary HTS assay was performed that substituted the Ac-CoA substrate with the CoA product, which showed 6b had similar activity in the absence of an acetyl source (Fig. 2C, 1). These data indicated that 6b could artifactually reduce the assay readout signal by reacting with the CoA, fluorescence quenching, or a combination of both. Notably, a counterscreen that spiked compounds into premade CPM-CoA demonstrated that most compounds with this sulfonamide chemotype did not generate light-based interferences.15 In cases where the purified enzymatic product is not easily obtained, one can allow the enzyme assay to proceed to near completion in the absence of compound, followed by addition of the compound after most of the substrate is consumed and immediate detection of the assay signal. This counterscreen should flag compounds that interfere with the detection step. To further confirm that chemotype 6 was interfering with the primary HTS readout by CoA scavenging, 6b and analogs were subjected to a thiol probe counterscreen by incubating compounds with glutathione (GSH) or CoA under assay-like conditions and then analyzing aliquots by liquid chromatography–mass spectrometry (LC-MS). This showed that multiple para-hydroxy-arylsulfonamides formed detectable adducts with GSH (Fig. 2C, 2, analog 6a shown) as well as CoA. In the AGM, it is noted that certain compounds can degrade in situ to form reactive substances.7 By monitoring parent compound stability under assay-like conditions, many compounds with chemotype 6 were also found to be unstable, and they were proposed to undergo degradation to form a reactive quinone or benzoquinone imine that could nonspecifically react with thiols (Fig. 2D).15
While counterscreens with small-molecule thiol probes are relatively straightforward, compounds can react at different rates with small-molecule and proteinaceous thiols due to microenvironment effects.16 Therefore, the AGM also recommends testing active compounds for nonspecific protein reactivity.7 As expected for a nonspecific reactive chemotype, compounds such as 6a formed detectable adducts with Rtt109 and/or Vps75 when analyzed by protein MS of trypsin-digested peptides.15 The sulfonamide chemotype 6 was also tested by ALARM NMR (a La assay to detect reactive molecules by nuclear magnetic resonance), an industry-developed counterscreen for nonspecific compound–protein thiol reactivity.17 Unlike other thiol probe counterscreens, ALARM NMR utilizes the human La antigen as a protein probe that contains two cysteines (C232 and C245) that produce pathognomonic chemical shift perturbations in nearby leucine residues (L249, L294, L296) by 2D heteronuclear multiple quantum coherence (HMQC) NMR. As expected, compounds such as 6b significantly perturbed the conformation of the hLa reporter in the absence of the thiol-scavenging reagent DTT, further incriminating this chemotype as nonspecific (Fig. 2C, 3). For performing these high-yield thiol probe assays, we refer readers to the detailed original protocols and the AGM chapter on reactivity for additional tips.7,17,18
Consistent with nonspecific activity, 6b showed similar inhibition of structurally unrelated KATs, yeast GCN5 and KAT3B (Fig. 2B). The combination of positive results from different interference counterscreens indicated that the HTS hits bearing this sulfonamide chemotype are nonspecifically reactive interference compounds.
It is recommended in the AGM to pair experimental evidence with so-called knowledge-based approaches incorporating cheminformatics, structure–activity relationship (SAR) analyses, and fundamental medicinal chemistry principles. It is our general philosophy to use cheminformatics as supportive tools for compound triage decisions (flagging for liabilities), as the overzealous use of filters and calculated descriptors may inadvertently discard an optimizable compound series.
The Rtt109 HTS contained multiple members of the sulfonamide chemotype 6, allowing for the construction of an apparent SAR (Fig. 2D, left). Compounds lacking the para-hydroxy group or naphthalene moiety were markedly less active, whereas potent activity was maintained with a variety of aryl substituents adjacent to the sulfone and/or stable leaving groups in the meta-position on the naphthol moiety. Without knowledge of the underlying nonspecific reactivity, one might erroneously conclude that such SAR is promising, when in fact it represents what are often called structure–interference relationships (SIRs), in that the relationship between apparent activity and a chemical structure is best explained by compound-dependent interference.
Cheminformatics analyses added further confidence in triaging this chemotype. The AGM chapter on reactivity interference recommends subjecting screening hits to a variety of substructure filters such as pan assay interference compounds (PAINS), rapid elimination of swill (REOS), and Lilly medicinal chemistry alerts.19,20 Performing these calculations can be done with open-source tools such as FAF-Drugs4.21 In this case, the sulfonamide chemotype was flagged by all three sets of filters (Fig. 2D, right). Triage by substructure alerts alone can be an efficient albeit blunt tool, as there are notable limitations to filters such as PAINS. Specifically, the generalizability of PAINS filters has been questioned due to their development from a single data source and assay technology (AlphaScreen), and concerns with wider applicability and overzealously removing potentially tractable compounds.22–24 Therefore, we recommend that compounds flagged by such filters be subjected to close scrutiny, using the structural alerts as guidance for implementing appropriate follow-up interference counterscreens. An analysis of historical screening performance using the open-source BadApple tool also flagged 6 as a probable promiscuous chemotype (Fig. 2D, bottom right).25 Along with mining screening and activity data, the AGM recommends a search of the scientific and patent literature for key compounds. In this case, a structure-based query using SciFinder Scholar readily identified this sulfonamide chemotype as prototypical redox cyclers and among reported inhibitors of KEAP1-NRF2, BRAF V600E, and MCL-1.26–29
Based on the results summarized above, the Rtt109 hits containing the para-hydroxy arylsulfonamide chemotype were triaged as nonspecific reactive compounds. Importantly, this triage was based on multiple lines of both experimental and knowledge-based evidence. Key learning points from this case study include the importance of performing counterscreens for nontechnology interferences even if activity is confirmed by an orthogonal assay technology, the importance of assessing thiol and target selectivity in the context of reactive compounds, and that apparent SAR can be explained by chemotype nonspecific reactivity. It is worth noting here that although compound reactivity has been successfully leveraged for targeted covalent inhibitors,30,31 utilizing this mechanism to design selective small molecules is not trivial or appropriate for all targets or biological contexts.
Case Study 2: Nonspecific Colloidal Aggregators
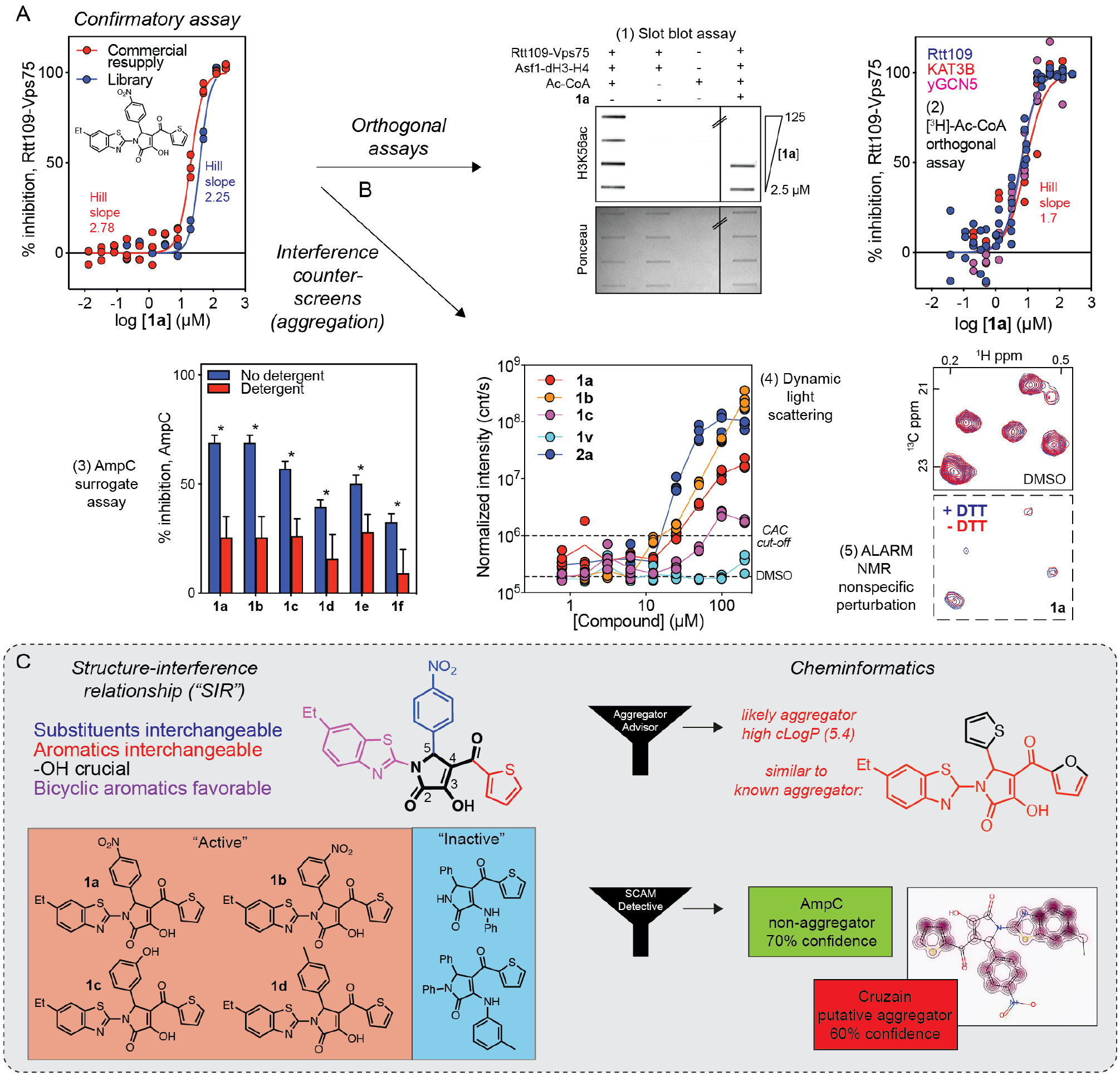
In the aforementioned Rtt109 HTS (case study 1, Z′ ~ 0.7), an unrelated 4-aroyl-1,5-disubstituted-3-hydroxy-2H-pyrrol-2-one chemotype 1 initially showed intriguing low micromolar activity.32 In accordance with recommended practices, similar activity of the key analog 1a was confirmed using a fresh powder sample obtained from the vendor (Fig. 3A). As the fluorometric assay indirectly measures Rtt109-catalyzed histone acetylation via quantifying the CoA by-product, the same reaction aliquots were analyzed by a slot blot and a [3H]-Ac-CoA filter-binding assay to further confirm 1a-dependent decreases in histone acetylation (Fig. 3B, 1 and 2). Notably, 1a showed similar activity versus two other KATs, human KAT3B and yeast GCN5, suggesting this compound might not be sufficiently specific (Fig. 3B, 2).
Case study of a compound aggregator identified from a high-throughput screen. (A) The activity of the representative 4-aroyl-1,5-disubstituted-3-hydroxy-2H-pyrrol-2-one 1a was confirmed in the primary Rtt109 assay using a fresh powder sample obtained from the vendor. (B) The activity of 1a was confirmed by two orthogonal assays: (1) a slot blot assay for H3K56ac in reaction aliquots from the primary HTS and (2) the aforementioned [3H]-Ac-CoA filter-binding assay. (C) Despite being active in orthogonal assays, 1a was suspected of acting by a non-technology-related interference. Compound 1a was active in counterscreens for colloidal aggregation, including (1) an AmpC surrogate assay to establish detergent-dependent inhibition and poor selectivity, (2) DLS for biophysical demonstration of colloid formation, and (3) ALARM NMR to demonstrate nonspecific protein perturbation independent of DTT. (D) Informatics supported the decision to triage chemotype 1: the apparent SAR was best explained by aggregation-based SIRs, while cheminformatics tools (Aggregator Advisor, SCAM Detective) flagged 1a as a potential aggregator. Portions of this figure were adapted from a previous publication.32
In the AGM chapter on aggregation, it is recommended to analyze concentration–response curves (CRCs) for apparent cooperativity. This is because colloidal aggregates tend to exhibit steep CRCs upon reaching the critical aggregation concentration (CAC).33 Notably, in all three assay formats (CPM, slot blot, [3H]-Ac-CoA), the CRCs for 1a were remarkable for steep Hill slopes (i.e., > 1.5), suggestive of compound aggregation.
Given this potential for colloidal aggregation, a series of counterscreens focused on compound aggregation were performed. First, 1a and chemical analogs were tested for detergent-sensitive inhibition of the unrelated enzyme AmpC β-lactamase. As expected for colloidal aggregators, the inhibition of AmpC by 1a was significantly attenuated by the inclusion of the nonionic detergent Triton X-100 (Fig. 3B, 3). For instructions on how to perform this high-yield surrogate assay, we refer readers to a detailed original protocol and the AGM chapter on aggregation for additional tips.6,34 Along with AmpC, we and others have had comparable successes identifying likely aggregators by assaying for detergent-sensitive malate dehydrogenase (MDH) inhibition.35 The results of a second counterscreen by dynamic light scattering (DLS) indicated that 1a–1c formed detectable colloids at low micromolar concentrations. Interestingly, the analog 2a, which was inactive against Rtt109 and AmpC, also formed detectable colloids by DLS. Another inactive analog, 1v, did not form detectable colloids (Fig. 3B, 4). While unexpected, the DLS data of 2a may be explainable: (1) given the sensitivity of aggregation to experimental conditions, 2a might not have formed aggregates during the Rtt109 or AmpC assays, and (2) it is possible that the 2a colloidal aggregates, while detectable by DLS, may not functionally perturb Rtt109 or AmpC. Finally, a third counterscreen for aggregation, ALARM NMR, showed that 1a perturbed the hLa conformation independent of DTT, which is suggestive of nonspecific protein binding that is consistent with colloidal aggregation (Fig. 3B, 5). These data highlight the importance of multiple counterscreens for aggregation, including both functional and biophysical assays.
The lack of other promising lead compounds from the Rtt109 HTS allowed for a focused exploration of chemotype 1 using “SAR by commerce” (Fig. 3C, left). The inhibition of Rtt109 by the 2H-pyrrol-2-one chemotype tolerated a variety of aryl substituents at the C5 position, while the thiophene and furan substitutes at the C4 position were essentially interchangeable. However, the hydroxy group at the C3 position and fused aromatics from the N1 position were otherwise essential for Rtt109 activity. Interestingly, these structural features correlated with detergent-sensitive AmpC inhibition, indicating that this apparent SAR is more appropriately viewed as SIR, in that the relationship between apparent activity and chemical structure is best explained by the propensity to form colloidal aggregates.
In addition to experimental approaches, the AGM chapter on aggregation interference also recommends subjecting screening hits to a variety of cheminformatics methods, which added further confidence in triaging this chemotype (Fig. 3C, right). Compound 1a and close analogs were not flagged by PAINS, REOS, or Lilly filters. However, 1a was flagged as a likely aggregator by two open-source aggregation filters, Aggregator Advisor and SCAM Detective (Fig. 3C, right).36,37
In summary, compounds bearing this 2H-pyrrol-2-one chemotype 1 were triaged as nonspecific colloidal aggregators. As with the reactive sulfonamide chemotype 6, this triage was based on multiple lines of both experimental and knowledge-based evidence. Important lessons from this case study are that aggregators can still inhibit a biochemical target even in the presence of a nonionic detergent, that there are occasional discrepancies between DLS and functional aggregation counterscreens, and that apparent SAR can be explained by the propensity of a chemotype to form colloidal aggregates.
Case Study 3: Potent Yet Nonspecific Bioactivity
A cell-free bioluminescence assay was developed to identify small-molecule inhibitors of the histone lysine methyltransferase nuclear receptor-binding SET domain protein 2 (NSD2) as potential anticancer agents (Fig. 4A),38 In this highly robust assay (Z′ = 0.92), NSD2 catalyzes the methylation of nucleosome substrates using S-adenosyl-l-methionine (SAM) to produce mono- and dimethylated histones and S-adenosyl-l-homocysteine (SAH). The NSD2 activity is indirectly quantified with the MTase-Glo technology, which enzymatically couples the SAH reaction product to ATP production, and the resulting ATP is converted to a bioluminescent signal by a luciferase reagent.39
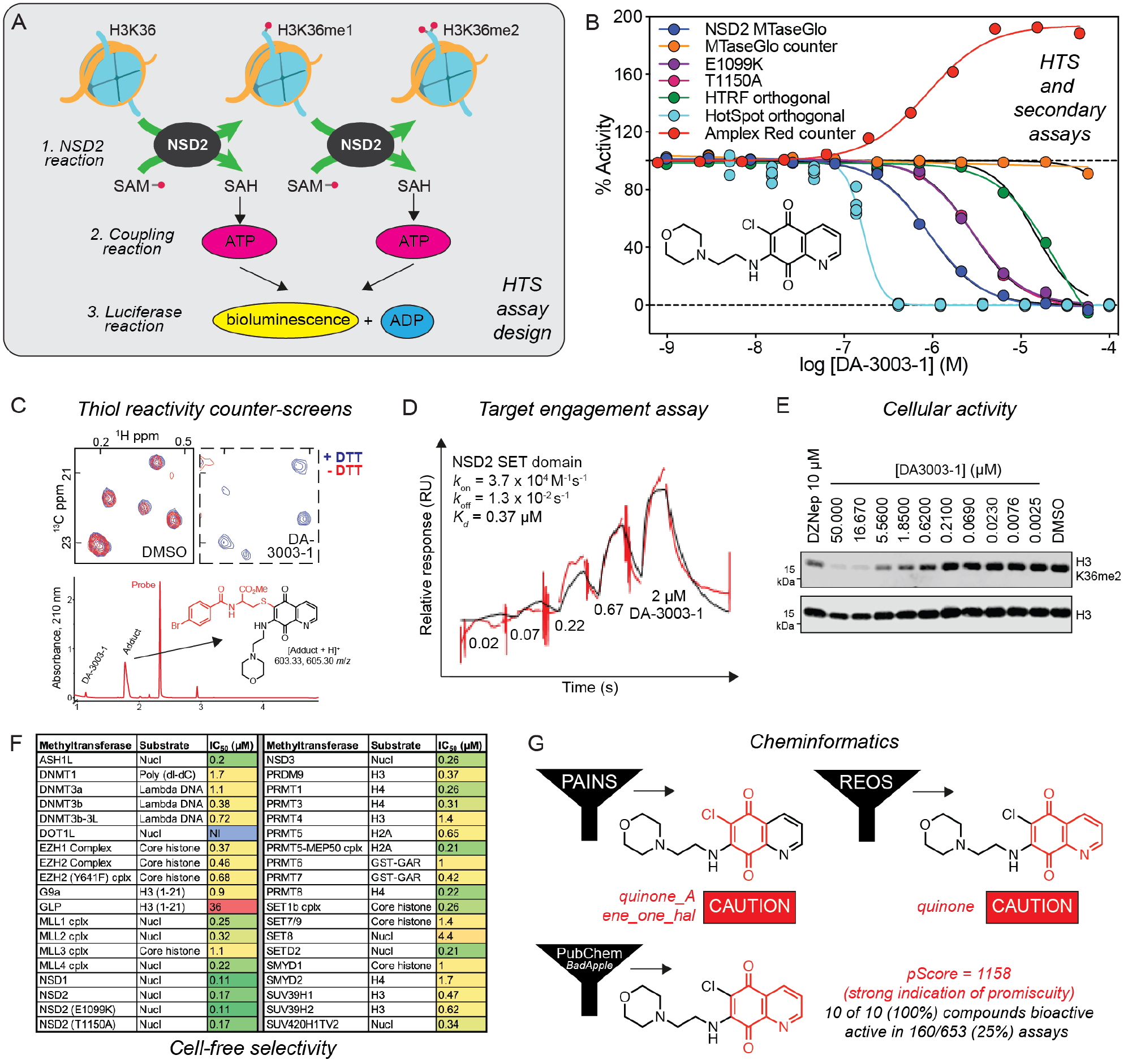
Putting it all together: a case study identifying potent but nonspecific bioactivity from an epigenetic HTS campaign. (A) A schematic of the primary HTS assay for NSD2-catalyzed histone methylation. (B) NSD2 inhibitory activity of DA-3003-1 in the MTase-Glo primary (dark blue), orthogonal HotSpot (light blue), and HTRF (green) assays, and counterscreens with the NSD2 variants E1099K (purple) and T1050A (magenta) using the MTase-Glo assay. Counterscreens were also performed for MTase-Glo interference (orange) and redox activity (Amplex Red, red). (C) Additionally, the ALARM NMR (top) and LC-MS (bottom) counterscreens were performed to identify nonspecific thiol reactivity. (D) A raw SPR sensorgram demonstrating direct binding of DA-3001-1 to NSD2. (E) Nonspecific modulation of an NSD2-related biomarker H3K36me2 by DA-3001-1 in U-2 OS cells from a Western blot. (F) Results of biochemical selectivity profiling of DA-3001-1 against 38 epigenetic targets using the [3H]-SAM-based HotSpot assay. (G) Cheminformatics analyses by substructure filters and historical screening performance. Portions of this figure were adapted from a previous publication.38
During the course of a pilot quantitative HTS (qHTS) with 16,000 compounds enriched with pharmacologically active small molecules,40 the compound DA-3003-1 (also known as NSC-663284) showed promising low micromolar potency (Fig. 4B, dark blue). The AGM recommends that scientists perform technology-related counterscreens with active compounds to characterize any technology-related compound interferences. Indeed, the primary MTase-Glo assay is potentially susceptible to compounds that modulate luciferases.5 However, DA-3003-1 showed negligible interference in an MTase-Glo interference counterscreen (Fig. 4B, orange). Inhibition of NSD2 by DA-3003-1 was further supported by both the [3H]-SAM-based HotSpot and the homogeneous time-resolved fluorescence (HTRF) orthogonal assays (Fig. 4B, light blue and green, respectively). Furthermore, DA-3003-1 also inhibited the activities of the NSD2 E1099K and T1150A variants in MTase-Glo counterscreens (Fig. 4B, purple and magenta, respectively).
In addition to a technology-related counterscreen and multiple orthogonal assays, DA-3003-1 was further subjected to several counterscreens for nonspecific reactivity. In an Amplex Red counterscreen for redox activity, DA-3003-1 produced hydrogen peroxide that mirrored its activity in the primary MTase-Glo assay (Fig. 4B, red). Interestingly, DA-3003-1 was ALARM NMR-positive in both the presence and absence of the thiol-scavenging reagent DTT, which would be consistent with nonspecific reactivity in DTT-free conditions and redox cycling when DTT is present (Fig. 4C, top). An additional LC-MS counterscreen for nonproteinaceous thiol reactivity detected the expected addition–elimination adduct between DA-3003-1 and a cysteine derivative optimized for LC-MS detection (Fig. 4C, bottom).41
At this point, it would have been reasonable to triage DA-3003-1 based on these data alone. As one of the goals of this project was to establish a screening pipeline for NSD2 inhibitor discovery, selected active compounds from the primary HTS were tested in several follow-up assays to better characterize the performance of the NSD2 screening cascade.
In target-based discovery, it is critical to demonstrate direct target engagement. This is often evaluated by surface plasmon resonance (SPR), isothermal titration calorimetry (ITC), or differential scanning fluorimetry (DSF). Interestingly, SPR indicated that DA-3003-1 binds the NSD2 SET domain with a strong affinity that is comparable to its biochemical potency (Kd = 370 nM) (Fig. 4D). Altogether, these data suggest that DA-3003-1 binds the NSD2 SET domain directly and inhibits its methyltransferase activity. However, it is critical to realize that target engagement does not necessarily indicate tractability, as apparent target engagement may be due to potentially undesirable MOAs such as nonspecific covalent modification or breakdown of the parent compound.
In U-2 OS cells, DA-3003-1 showed a concentration-dependent reduction in cellular H3K36me2 (IC50 = 545 nM), a biomarker of NSD2 activity (Fig. 4E). While this finding might appear promising, caution should be exercised when interpreting the results, because cellular events associated with cytotoxicity can be overinterpreted as mechanism-based compound-dependent activity. Indeed, DA-3003-1 demonstrated a concentration-dependent cytotoxicity in U-2 OS cells (CC50 = 270 nM) with a potency comparable to that of the biomarker modulation. Under such conditions, reductions in cellular H3K36me2 could result from the modulation of pathways other than NSD2. When profiled against 38 methyltransferases using the [3H]-SAM-based HotSpot biochemical assay, DA-3003-1 primarily showed nanomolar and some low micromolar potencies against all but two of the epigenetic targets (Fig. 4F). This combination of similar potencies and a lack of selectivity strongly supports a nonspecific MOA.
As with the previous case studies, analyzing hits from a screen such as DA-3003-1 by cheminformatics can complement experimentally based triage methods. When subjected to several substructure filters, DA-3003-1 was flagged by the PAINS, REOS, and Lilly alerts due to its quinone moiety, while analysis of historical screening performance in PubChem using BadApple demonstrated that DA-3003-1 was grossly nonspecific (Fig. 4G). Searching SciFinder Scholar and PubMed readily identified DA-3003-1 as a reported inhibitor of MKP-1, Cdc25, and SET8 (among others), along with being a prototypical redox cycler.26,42–45 In retrospect with this particular case and the abundance of available activity data, there would have been a compelling argument for triaging DA-3003-1 based on cheminformatics alone.
In summary, DA-3003-1 should be triaged as a potent, yet nonspecific reactive and redox-active compound based on multiple lines of both experimental and knowledge-based evidence. Important lessons from this particular case study are that nonspecific reactive compounds can display activity by SPR and cellular assays such as Western blots.
Discussion
Using two previously reported HTS campaigns as model case studies, we show how best practices such as those codified in the AGM can be used to effectively identify nonspecific reactive and aggregating compounds from an HTS. Our case studies provide several examples in which small molecules can demonstrate misleading results by appearing to have desirable biological activity. Careful design of the primary, secondary, and counterscreens for an HTS campaign will substantially reduce the number of nuisance compounds selected for further analysis. However, these case studies illustrate how nonspecific compounds with intractable MOAs can demonstrate reproducible biological activity in a range of orthogonal assays, apparent SAR, direct target engagement, and even apparent activity in cell-based assays. This underscores the substantial efforts that are required during a post-HTS triage to identify and confirm any compounds with genuine activity. Such rigorous characterizations are critical to avoid the misplacement of resources, both in-house and by the wider research community.
Post-HTS triage requires a careful balance of competing factors including the relative weight of experimental versus informatics-based evidence, the optimal order and composition of secondary assays, and even intuition.46 In practice, it can be resource-prohibitive and even excessive to rigorously investigate every screening hit for every interference. Based on our experiences with the Rtt109 and NSD2 HTS campaigns, high-yielding secondary assays were orthogonal confirmations of activity, followed by a combination of counterscreens for aggregation and nonspecific reactivity. In retrospect, while testing for KAT selectivity was helpful, there was a view that pan-KAT inhibitors might still be optimizable in the absence of additional off-targets. In this respect, interference counterscreens utilizing completely unrelated targets (AmpC, ALARM NMR) provided additional confidence that the observed activities were truly nonspecific. In these case studies, informatics were most helpful to increase our index of suspicion for these interference chemotypes, and later for providing additional confidence in our triage decisions.
The NIH/NCATS AGM “Assay Artifacts and Interferences” section is frequently updated to reflect advances in best practices. As such, the AGM provides a wealth of information to assist investigators in assay design and the selection of a testing funnel of assays to mitigate compound-dependent artifacts and other interferences. While the aforementioned case studies focused on cell-free assays, many of the same experimental and informatics principles for assessing compound reactivity and colloidal aggregation apply to more complex cellular assays.12 Future content for the AGM “Assay Artifacts and Interferences” section will likely address (1) compound interferences in cellular assays, (2) targeted covalent modulators and covalent fragments, and (3) compound interferences in biophysical assays.
Footnotes
Acknowledgements
J.L.D. gratefully acknowledges Henry O’Donnell and Dr. Brian Shoichet for performing DLS experiments. The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: B.K.W. (Ono Pharma Foundation; NIH High-End Instrumentation Program, S10-OD026839; NIDDK, U01-DK123717); J.L.D. (intramural program of NCATS and NIH); N.P.C. (federal funds from the National Cancer Institute, National Institutes of Health, under contract HHSN261200800001I).
ORCID iDs
Nathan P. Coussens
Douglas S. Auld
Jonathan R. Thielman
Jayme L. Dahlin
References
1.
ThorneN.AuldD. S.IngleseJ.Apparent Activity in High-Throughput Screening: Origins of Compound-Dependent Assay Interference. Curr. Opin. Chem. Biol. 2010, 14, 315–324.
2.
NelsonK.DahlinJ.BissonJ., et al. The Essential Medicinal Chemistry of Curcumin. J. Med. Chem. 2016, 60, 1620–1637.
3.
SimeonovA.DavisM. I.Interference with Fluorescence and Absorbance. In Assay Guidance ManualSittampalamG.CoussensN. Eds. Eli Lilly and the National Center for Advancing Translational Science: Bethesda, MD, 2015.
4.
CoussensN. P.AuldD.RobyP., et al. Compound-Mediated Assay Interferences in Homogenous Proximity Assays. In Assay Guidance ManualSittampalamG.CoussensN. Eds. Eli Lilly and the National Center for Advancing Translational Science: Bethesda, MD, 2020.
5.
AuldD. S.IngleseJ.Interferences with Luciferase Reporter Enzymes. In Assay Guidance ManualSittampalamG.CoussensN. Eds. Eli Lilly and the National Center for Advancing Translational Science: Bethesda, MD, 2016.
6.
AuldD. S.IngleseJ.DahlinJ. L.Assay Interference by Aggregation. In Assay Guidance ManualSittampalamG.CoussensN. Eds. Eli Lilly and the National Center for Advancing Translational Science: Bethesda, MD, 2017.
7.
DahlinJ. L.BaellJ. B.WaltersM. A.Assay Interference by Chemical Reactivity. In Assay Guidance ManualSittampalamG.CoussensN. Eds. Eli Lilly and the National Center for Advancing Translational Science: Bethesda, MD, 2015.
8.
HermannJ. C.ChenY.WartchowC., et al. Metal Impurities Cause False Positives in High-Throughput Screening Campaigns. ACS Med. Chem. Lett. 2013, 4, 197–200.
9.
MorrealeF.TestaA.ChauguleV., et al. Mind the Metal: A Fragment Library-Derived Zinc Impurity Binds the E2 Ubiquitin-Conjugating Enzyme Ube2T and Induces Structural Rearrangements. J. Med. Chem. 2017, 60, 8183–8191.
10.
WipfP.ArnoldD.CarterK., et al. A Case Study from the Chemistry Core of the Pittsburgh Molecular Library Screening Center: The Polo-Like Kinase Polo-Box Domain (Plk1-PBD). Curr. Topics Med. Chem. 2009, 9, 1194–1205.
11.
TarziaG.AntoniettiF.DurantiA., et al. Identification of a Bioactive Impurity in a Commercial Sample of 6-Methyl-2-p-Tolylaminobenzo[d][1,3]oxazin-4-one (URB754). Ann. Chim. 2007, 97, 887–894.
12.
DahlinJ. L.AuldD. S.RothenaignerI., et al. Nuisance Compounds in Cellular Assays. Cell Chem. Biol. 2021, 28, 356–370.
13.
CoussensN. P.SittampalamG. S.GuhaR., et al. Assay Guidance Manual: Quantitative Biology and Pharmacology in Preclinical Drug Discovery. Clin. Transl. Sci. 2018, 11, 461–470.
14.
DahlinJ. L.SinvilleR.SolbergJ., et al. A Cell-Free Fluorometric High-Throughput Screen for Inhibitors of Rtt109-Catalyzed Histone Acetylation. PLoS One2013, 8, e78877.
15.
DahlinJ. L.NissinkJ. W. M.StrasserJ. M., et al. PAINS in the Assay: Chemical Mechanisms of Assay Interference and Promiscuous Enzymatic Inhibition Observed during a Sulfhydryl-Scavenging HTS. J. Med. Chem. 2015, 58, 2091–2113.
16.
WilsonJ. M.WuD.Motiu-DeGroodR., et al. A Spectrophotometric Method for Studying the Rates of Reaction of Disulfides with Protein Thiol Groups Applied to Bovine Serum Albumin. J. Am. Chem. Soc. 1980, 102, 359–363.
17.
HuthJ. R.MendozaR.OlejniczakE. T., et al. ALARM NMR: A Rapid and Robust Experimental Method to Detect Reactive False Positives in Biochemical Screens. J. Am. Chem. Soc. 2005, 127, 217–224.
18.
DahlinJ.SinghG.CuellarM., et al. ALARM NMR for HTS Triage and Chemical Probe Validation. Curr. Protoc. Chem. Biol. 2018, 10, 91–117.
19.
BaellJ. B.HollowayG. A.New Substructure Filters for Removal of Pan Assay Interference Compounds (PAINS) from Screening Libraries and for Their Exclusion in Bioassays. J. Med. Chem. 2010, 53, 2719–2740.
20.
WaltersW. P.NamchukM.Designing Screens: How to Make Your Hits a Hit. Nat. Rev. Drug Discov. 2003, 2, 259–266.
21.
LagorceD.SperandioO.BaellJ., et al. FAF-Drugs3: A Web Server for Compound Property Calculation and Chemical Library Design. Nucleic Acids Res. 2015, 43, W200–W207.
22.
ChakravortyS. J.ChanJ.GreenwoodM. N., et al. Nuisance Compounds, PAINS Filters, and Dark Chemical Matter in the GSK HTS Collection. SLAS Discov. 2018, 23, 532–545.
23.
VidlerL. R.WatsonI. A.MargolisB. J., et al. Investigating the Behavior of Published PAINS Alerts Using a Pharmaceutical Company Data Set. ACS Med. Chem. Lett. 2018, 9, 792–796.
24.
BaellJ.NissinkJ.Seven Year Itch: Pan-Assay Interference Compounds (PAINS) in 2017—Utility and Limitations. ACS Chem. Biol. 2018, 13, 36–44.
25.
YangJ. J.UrsuO.LipinskiC. A., et al. BadApple: Promiscuity Patterns from Noisy Evidence. J. Cheminform. 2016, 8, 29.
26.
SoaresK. M.BlackmonN.ShunT. Y., et al. Profiling the NIH Small Molecule Repository for Compounds That Generate H2O2 by Redox Cycling in Reducing Environments. Assay Drug Dev. Technol. 2010, 8, 152–174.
27.
ZhuangC.NarayanapillaiS.ZhangW., et al. Rapid Identification of Keap1-Nrf2 Small-Molecule Inhibitors through Structure-Based Virtual Screening and Hit-Based Substructure Search. J. Med. Chem. 2014, 57, 1121–1126.
28.
QinJ.XieP.VentocillaC., et al. Identification of a Novel Family of BRAFV600E Inhibitors. J. Med. Chem. 2012, 55, 5220–5230.
29.
AbulwerdiF.LiaoC.LiuM., et al. A Novel Small-Molecule Inhibitor of mcl-1 Blocks Pancreatic Cancer Growth In Vitro and In Vivo. Mol. Cancer Ther. 2014, 13, 565–575.
30.
SinghJ.PetterR.BaillieT., et al. The Resurgence of Covalent Drugs. Nat. Rev. Drug Discov. 2011, 10, 307–317.
31.
StrelowJ.A Perspective on the Kinetics of Covalent and Irreversible Inhibition. SLAS Discov. 2017, 22, 3–20.
32.
DahlinJ. L.NissinkJ. W. M.FrancisS., et al. Post-HTS Case Report and Structural Alert: Promiscuous 4-Aroyl-1,5-Disubstituted-3-Hydroxy-2H-pyrrol-2-one Actives Verified by ALARM NMR. Bioorg. Med. Chem. Lett. 2015, 25, 4740–4752.
33.
ShoichetB. K.Interpreting Steep Dose-Response Curves in Early Inhibitor Discovery. J. Med. Chem. 2006, 49, 7274–7277.
34.
FengB. Y.ShoichetB. K.A Detergent-Based Assay for the Detection of Promiscuous Inhibitors. Nat. Protoc. 2006, 1, 550–553.
35.
DuanD.DoakA.NedyalkovaL., et al. Colloidal Aggregation and the In Vitro Activity of Traditional Chinese Medicines. ACS Chem. Biol. 2015, 10, 978–988.
36.
IrwinJ.DuanD.TorosyanH., et al. An Aggregation Advisor for Ligand Discovery. J. Med. Chem. 2015, 58, 7076–7087.
37.
AlvesV. M.CapuzziS. J.BragaR., et al. SCAM Detective: Accurate Predictor of Small, Colloidally-Aggregating Molecules. J. Chem. Inf. Model. 2020, 60, 4056–4063.
38.
CoussensN. P.KalesS. C.HendersonM. J., et al. High-Throughput Screening with Nucleosome Substrate Identifies Small-Molecule Inhibitors of the Human Histone Lysine Methyltransferase NSD2. J. Biol. Chem. 2018, 293, 13750–13765.
39.
HsiaoK.ZegzoutiH.GoueliS. A.Methyltransferase-Glo: A Universal, Bioluminescent and Homogenous Assay for Monitoring All Classes of Methyltransferases. Epigenomics2016, 8, 321–339.
40.
IngleseJ.AuldD.JadhavA., et al. Quantitative High-Throughput Screening: A Titration-Based Approach That Efficiently Identifies Biological Activities in Large Chemical Libraries. Proc. Nat. Acad. Sci. U.S.A. 2006, 103, 11473–11478.
41.
ReimerD.HughesC. C.Thiol-Based Probe for Electrophilic Natural Products Reveals That Most of the Ammosamides Are Artifacts. J. Nat. Prod. 2017, 80, 126–133.
42.
VogtA.McDonaldP. R.TamewitzA., et al. A Cell-Active Inhibitor of Mitogen-Activated Protein Kinase Phosphatases Restores Paclitaxel-Induced Apoptosis in Dexamethasone-Protected Cancer Cells. Mol. Cancer Ther. 2008, 7, 330–340.
43.
LazoJ. S.AslanD. C.SouthwickE. C., et al. Discovery and Biological Evaluation of a New Family of Potent Inhibitors of the Dual Specificity Protein Phosphatase Cdc25. J. Med. Chem. 2001, 44, 4042–4049.
44.
JohnstonP. A.SoaresK. M.ShindeS. N., et al. Development of a 384-Well Colorimetric Assay to Quantify Hydrogen Peroxide Generated by the Redox Cycling of Compounds in the Presence of Reducing Agents. ASSAY Drug Dev. Technol. 2008, 6, 505–518.
45.
BlumG.IbáñezG.RaoX., et al. Small-Molecule Inhibitors of SETD8 with Cellular Activity. ACS Chem. Biol. 2014, 9, 2471–2478.
46.
DahlinJ.WaltersM.The Essential Roles of Chemistry in High-Throughput Screening Triage. Future Med. Chem. 2014, 6, 1265–1290.