
Abstract
Carbapenem-resistant Enterobacteriaceae (CRE) are an emerging antimicrobial resistance threat for which few if any therapeutic options remain. Identification of new agents that either inhibit CRE or restore activity of existing antimicrobials is highly desirable. Therefore, a high-throughput screen of 182,427 commercially available compounds was used to identify small molecules which either enhanced activity of meropenem against a carbapenem-resistant Klebsiella pneumoniae ST258 screening strain and/or directly inhibited its growth. The primary screening methodology was a whole-cell screen/counterscreen combination assay that tested for reduction of microbial growth in the presence or absence of meropenem, respectively. Screening hits demonstrating eukaryotic cell toxicity based on an orthogonal screening effort or identified as pan-assay interference compounds (PAINS) by computational methods were triaged. Primary screening hits were then clustered and ranked according to favorable physicochemical properties. Among remaining hits, we found 10 compounds that enhanced activity of carbapenems against a subset of CRE. Direct antimicrobials that passed toxicity and PAINS filters were not, however, identified in this relatively large screening effort. It was previously shown that the same screening strategy was productive for identifying candidates for further development when screening known bioactive libraries inclusive of natural products. Our findings therefore further highlight liabilities of commercially available small-molecule screening libraries in the Gram-negative antimicrobial space. In particular, there was especially low yield in identifying compelling activity against a representative, highly multidrug-resistant, carbapenemase-producing K. pneumoniae strain.
Keywords
Introduction
Enterobacteriaceae are a common cause of bacterial bloodstream, urinary tract, and surgical site infections. Concerningly, these organisms also are commonly associated with resistance to clinically useful first- and second-line antimicrobials, including penicillins, cephalosporins, fluoroquinolones, and aminoglycosides. 1 Carbapenems are the major last line of defense against multidrug-resistant Gram-negative pathogens. Unfortunately, carbapenem-resistant Enterobacteriaceae (CRE) emerged rapidly during the past two decades. 2 They have now been detected worldwide, 3 with particularly high prevalence in Asia 4 and Southern Europe. 5 CRE are also isolated with increasing frequency in the United States. 6 Few if any treatments remain, and those few often have dose-limiting toxicity. 7 Recently, truly pan-drug-resistant CRE have appeared, 8 along with highly pathogenic hypermucoviscous strains that cause metastatic multi-organ infection in otherwise healthy adults,9,10 highlighting the pressing need for new antimicrobials with activity against these pathogens.
Development of new antimicrobials active against multidrug-resistant Gram-negative pathogens has proven difficult due to the relative impermeability of the Gram-negative cell membrane 11 and ubiquitous expression of efflux pumps. 12 Carbapenems overcome both of these challenges and may retain detectable in vitro 13 and in vivo 14 activity even in strains expressing enzymes that degrade carbapenems (carbapenemases). Therefore, we hypothesized that this partial activity could be potentiated by small molecules through a variety of mechanisms to restore carbapenem efficacy against otherwise resistant CRE.
We therefore chose to use our previously validated screening/counterscreening approach to evaluate activity of a large collection of small molecules for their ability to either directly inhibit or potentiate activity of a representative carbapenem (meropenem) against a CRE screening strain. 15 To rapidly triage compounds with nonspecific activity, we used data from an orthogonal screening effort to eliminate those with eukaryotic cytotoxicity. 16 Finally, we identified a series of compounds with optimal physicochemical properties, tested their spectrum of activity against representative CRE strains using commercially available compounds, and confirmed activity on resynthesis. Based on our observations, we believe the screening strategy will prove an efficient method for identifying direct and indirect antimicrobials, although only in libraries optimized for the Gram-negative antimicrobial space.
Materials and Methods
Primary Screening
Our primary screening strain was Klebsiella pneumoniae BIDMC12A, a CRE strain of sequence type 258 (ST258), the most common sequence type of K. pneumoniae CRE strains circulating in the United States; it expresses the KPC3 carbapenemase, and blaSHV11, blaSHV134, and blaTEM1 β-lactamases. 17 The screen was performed as a screening/counterscreening experiment, as described in our previous work in which only known bioactive compounds were examined. 15 Briefly, prior to screening, 30 µl of cation-adjusted Mueller Hinton broth (CAMHB; BD Diagnostics, Sparks, MD) containing 20 µg ml−1 meropenem (ArkPharm, Libertyville, IL) (screen) or no antibiotic (counterscreen) was added to clear, untreated polystyrene 384-well plates (Greiner Bio-One, Monroe, NC) using a MultiDrop Combi liquid handler (Thermo Fisher Scientific, Waltham, MA). Compounds were added using a pin-transfer robot calibrated to deliver 300 nL to each well and screened in duplicate in separate screening plates.
We screened commercially available libraries available at the Institute of Chemistry and Cell Biology (Harvard Medical School, Boston, MA), listed in Supplemental Material 2, which consist of small molecules without previously characterized activity. Compound concentrations varied by library. For libraries with concentrations expressed in µg mL−1, screening concentrations were 2.5, 10, 25, or 75 µg mL−1. For libraries with concentrations expressed as molarity, screening concentrations were 0.5, 5, 16.5, 44, or 50 µM. Immediately after compound transfer, 30 µl of K. pneumoniae BIDMC12A [1×106 colony-forming units (CFU) ml−1] in CAMHB was added, bringing the final concentration of cells to approximately 5×10 5 CFU ml−1 per Clinical and Laboratory Standards Institute (CLSI) guidelines 18 and meropenem (when applicable) to 10 µg mL−1 in a final assay volume of 60 µL.
Plates were incubated for 48 h at 37 °C in 100% humidity. Bacterial growth was quantified by optical density at 600 nm (OD600) using an EnVision multimode plate reader (PerkinElmer, Waltham, MA). For each plate, Z’ was calculated based on positive (5 µg mL−1 colistin) and negative controls (CAMHB alone). 19 Graphical representations of screening results were created using a custom Python script using the matplotlib library, 20 with point density calculated using the kernel density function as implemented in the scipy library. 21
Hit Identification and Confirmation
For each well, z-scores were calculated based on average and standard deviation of all experimental wells from the same assay plate. Direct antimicrobial hits were defined as strong (z < −6), moderate (−3 > z > −6), or weak (−1.5 > z > −3) based on the least significant z-score between replicates. Compounds were defined as potential adjunctives when the z-score for the screen was more than threefold that of the counterscreen. Based on previous work, 15 we selected hits with >50% inhibition in the screen as candidates for follow-up testing.
Hits with eukaryotic cell cytotoxicity were identified based on results from a separate orthogonal high-throughput screening (HTS) effort using the same compound libraries. 16 Briefly, the cytotoxicity assay consisted of application of compounds to J774A.1 macrophages incubated in the presence of 125 nM SYTOX Green, a membrane-impermeant nucleic acid–binding dye. In this assay, cytotoxicity results in increased eukaryotic cell membrane permeability and associated increase in SYTOX Green fluorescence, which is measured relative to controls. The assay was described previously as part of a combined screen for intracellular bacterial growth and eukaryotic cell death. 16 Cytotoxic compounds were defined conservatively as those with cytotoxicity z-scores >1.5.
Hits were cherrypicked for confirmatory testing from library plates using a Tecan EVO75 liquid handler (Tecan, Morrisville, NC). We then used an HP D300 digital dispenser (HP Inc., Palo Alto, CA) to add 300 nL of compound to CAMHB or CAMHB containing 10 µg mL−1 meropenem to replicate conditions of the screen and counterscreen. K. pneumoniae BIDMC12A was added to a concentration of 5×105 CFU mL−1 with a final assay volume of 60 µL. Plates were incubated at 37 °C in 100% humidity for 48 h, and growth was quantitated as described in the primary screen. Adjunctive activity was considered confirmed if it resulted in >25% growth inhibition in the presence of meropenem, but <25% growth inhibition in CAMHB alone, while direct activity was considered confirmed if inhibition were >25% in the absence of meropenem.
Secondary Analysis Using Commercially Available Compounds
Select compounds were ordered as powder from ChemDiv (San Diego, CA), ChemBridge (San Diego, CA), Enamine (Monmouth Junction, NJ), or Asinex (Winston-Salem, NC). Compounds were dissolved in 100% DMSO (Sigma-Aldrich, St. Louis, MO) to a concentration of 5 mg mL−1 and stored at −80 °C. For each compound, we performed two-dimensional synergy assays in combination with meropenem using a previously validated protocol.22,23 Briefly, we used an HP D300 digital dispenser to prepare combinatorial twofold orthogonal dilution series of meropenem and compounds of interest. Minimal inhibitory concentrations (MICs) were defined as the lowest concentration of antimicrobial resulting in complete growth inhibition (OD600 < 0.08), as previously validated by our laboratory. 24
In these experiments, we tested only meropenem potentiators with no detectable MIC value on their own. Therefore, synergy was assessed solely based on the greatest fold reduction of the meropenem MIC in the presence of compound, that is, the MIC of meropenem in the presence of compound divided by the MIC of meropenem alone, which is expressed as the fractional inhibitory concentration ratio (FIC). FIC values ≤0.5, consistently observable in biological replicates, were considered to indicate synergy.23,25
In-House Synthesis of Confirmed Hits
Selected compounds were resynthesized in-house for follow-up activity confirmation experiments. Materials, instrumentation used, and experimental details can be found in Supplemental Material 1 (Materials and Instrumentation) and Supplemental Material 6 (Synthesis).
Cheminformatics
Pan-assay interference compounds (PAINS) 26 filtering was performed through an available PAINS filter 27 (Eli Lilly, Cambridge, MA). Next, using Scaffold Hunter, hit compounds were arranged into clusters based on Tanimoto distance measurements of fingerprints generated for each molecule. 28 Physicochemical properties of compounds were predicted with Microsoft Excel (Microsoft, Redmond, WA) using add-ins from ChemDraw (PerkinElmer) and JChem for Office (ChemAxon, Cambridge MA). Compounds were scored through two multiparameter optimization (MPO) tools using these predicted properties.
First, compounds were ranked by a previously reported MPO algorithm for calculating optimal physicochemical properties of drug molecules with good bioavailability. 29 Furthermore, an “in-house” MPO algorithm was designed to predict the ability of a compound to penetrate a bacterial cell and avoid efflux, both characteristics of effective Gram-negative antimicrobial compounds. For this reason, we refer to our “in-house” MPO algorithm as PEMPO (permeation and efflux multiparameter optimization).
PEMPO scoring focused on assessing optimal ranges (shown in parentheses) for targeted physicochemical properties of Gram-negative antimicrobials, including the isoelectric point (6.1–8.7), the total polar surface area (100–200 Å 2 ), the number of hydrogen bond donors (HBDs; 2–6), the number of hydrogen bond acceptors (HBAs; 6–11), the partition coefficient clogP (≤3), and the distribution coefficient clogD7.4 (≤0.2). Optimal ranges were defined by analysis of average physicochemical properties of 100 known Gram-negative active antimicrobials from a study by Moser et al. 30 Compounds were then scored based on how similar each physicochemical property related to the optimal value (Suppl. Material 3c). Thus, a high scoring compound suggested a high probability for bacterial cell permeation and a low probability for efflux.
Two antibacterial classes were excluded from development of the PEMPO model: macrocycles (such as macrolides or cyclic peptides such as colistin) and aminoglycosides. Both compound classes exhibit a significantly higher molecular weight than most “druglike” compounds found within screening libraries and as a result would disproportionately influence the scoring of compounds based on extreme characteristics compared with other classes. Aminoglycosides contain on average 30 HBDs and HBAs, whereas the other six classes of antibacterials (penicillins, cephems, carbapenems, sulfa drugs, fluoroquinolones, and tetracyclines) contain on average 13 HBDs and HBAs. 30 Macrolides display many more lipophilic residues, contributing to a higher average cLogD7.4 of 2.6, whereas an average cLogD7.4 of −2.77 is observed among the other six antimicrobial classes. 30 Molecular weight was not used to calculate PEMPO scores because molecular weight of known Gram-negative compounds can vary widely based on compound class. The formula for calculating the PEMPO score is described in Supplemental Material 3a.
As physicochemical property calculators vary between platforms, we evaluated PEMPO scores calculated from properties generated by Pipeline Pilot (Accelrys, San Diego, CA) and ACD/Labs (Toronto, ON, Canada) compared to scores generated from Chemdraw and ChemAxon. Known Gram-negative antimicrobials had an average PEMPO score of 4.97 and 5.08 out of 6.0, respective to property prediction platform. Results listed by compound can be found in Supplemental Material 3b. We observed an average increased PEMPO score of +0.11 using the latter compared to the former property generation tools, but considered this difference to be negligible.
Spectrum of Activity Testing
Follow-up activity spectrum studies were performed for selected compounds. We tested commercially available or resynthesized compounds in combination with meropenem as described above, using 30 de-identified CRE isolates collected at our institution, including Escherichia coli (n = 8), K. pneumoniae (n = 20), Serratia marcescens (n = 1), and Enterobacter cloacae (n = 1). The genome sequences of all strains are available. 17
Construction of Carbapenemase-Expressing E. coli Strains
KPC2, KPC3, and NDM1 carbapenemases were PCR amplified with Q5 DNA polymerase (New England Biolabs, Ipswich, MA; annealing temperature = 60 °C) using primers listed in Supplemental Material 4. PCR products were introduced into the pUC19 vector using the NEBuilder HiFi DNA Assembly Kit (New England Biolabs) according to the manufacturer’s instructions. Vectors alone or vectors containing carbapenemases were transformed into electrocompetent DH5α (New England Biolabs); tolC mutant strain, JW5503-1 (E. coli Genetic Resources Stock Center, Yale University, New Haven, CT); or lptD mutant strain, RFM795 (E. coli Genetic Resources Stock Center), and selected with 100 µg mL−1 ampicillin (Thermo Fisher Scientific). Cloning fidelity was confirmed by DNA sequencing (Genewiz, Boston, MA). Carbapenemase expression was confirmed phenotypically by determination of meropenem MICs. 24 Select resynthesized compounds were tested for synergy with meropenem in all constructed strains, as outlined in the Secondary Analysis Using Commercially Synthesized Compounds section.
Results
Primary Screening
Gram-negative bacteria are intrinsically resistant to a variety of antibiotics owing to the relative impermeability of the cell envelope. In addition, multidrug-resistant organisms have an extensive system of efflux pumps with broad and unpredictable specificities, which also prevent molecules from reaching the cytoplasm. 12 Therefore, we chose to perform a whole-cell bacterial growth inhibition screen so that screening hits would have already passed these two significant hurdles. Furthermore, our screening strain was a representative ST258, multidrug-resistant clinical K. pneumoniae isolate, representative of the most common CRE strains circulating in the United States, and resistant to a variety of antimicrobial agents, including penicillins, cephalosporins, carbapenems, fluoroquinolones, nitrofurantoin, trimethoprim/sulfamethoxazole, tobramycin, and amikacin. 15 Thereby, a high bar for activity was set, which is appropriate for identifying efficacy against an emerging multidrug-resistant pathogen target.
Our HTS assay was designed as a screen/counterscreen. Screening wells contained meropenem at a subinhibitory concentration of 10 µg mL−1 and the counterscreen contained no antimicrobial. Therefore, compounds that potentiated meropenem would demonstrate activity in the screen (growth inhibition in the presence of meropenem) but not in the counterscreen (without meropenem). Direct antimicrobials would exhibit inhibitory effects independent of meropenem and therefore demonstrate activity in both the screen and counterscreen. In total, we screened 182,427 compounds without previously characterized biological activity in duplicate using this two-tiered assay. An overview of the screening effort and secondary analysis is summarized in Figure 1 . During the screening effort, we found good reproducibility between replicates for both the screen and counterscreen experiments ( Fig. 2 ). Average cumulative Z’ was 0.61 for the screen and 0.67 for the counterscreen based on positive and negative screening wells from screening plates.
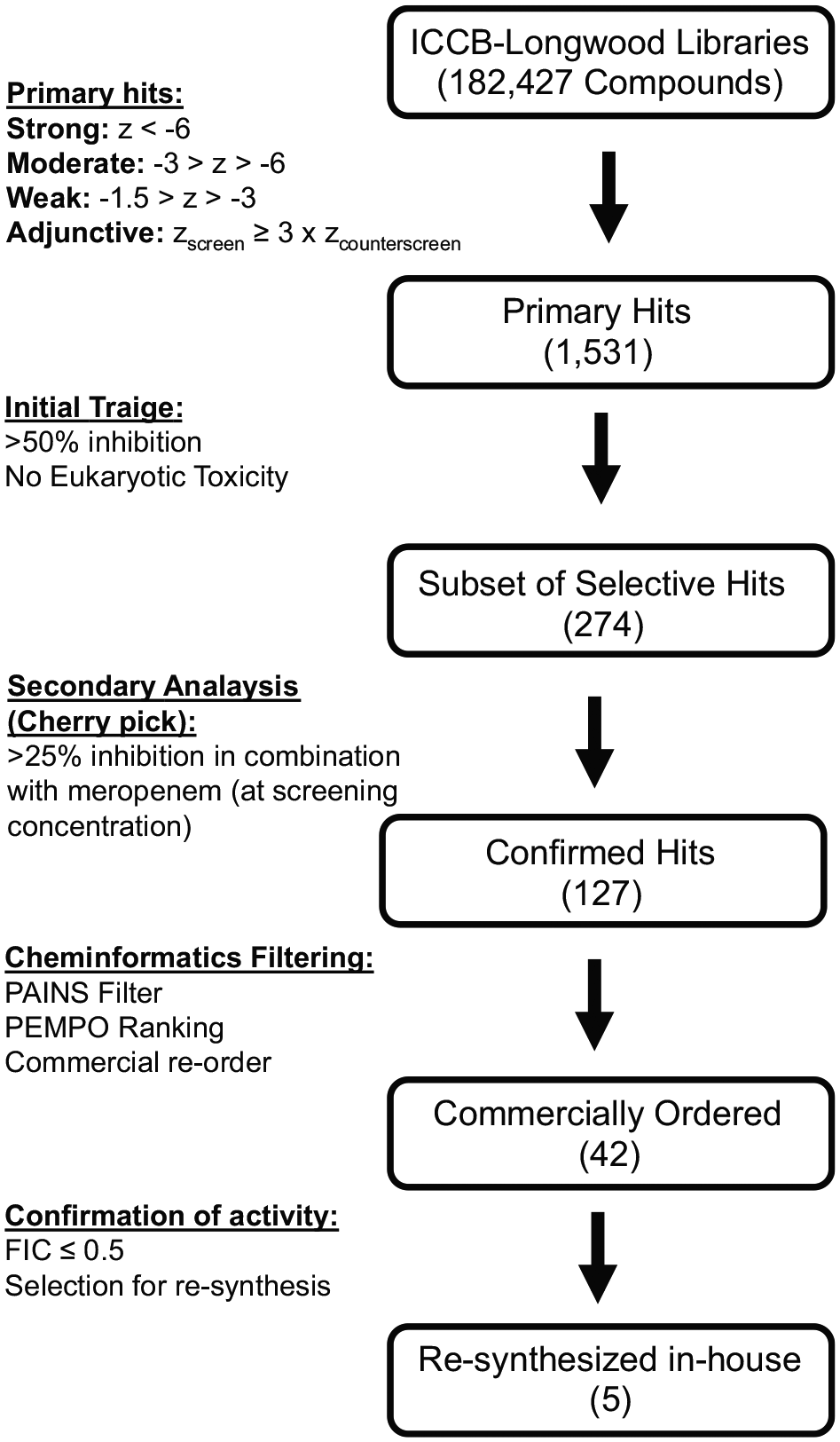
Overview of high-throughput screening hit analysis.
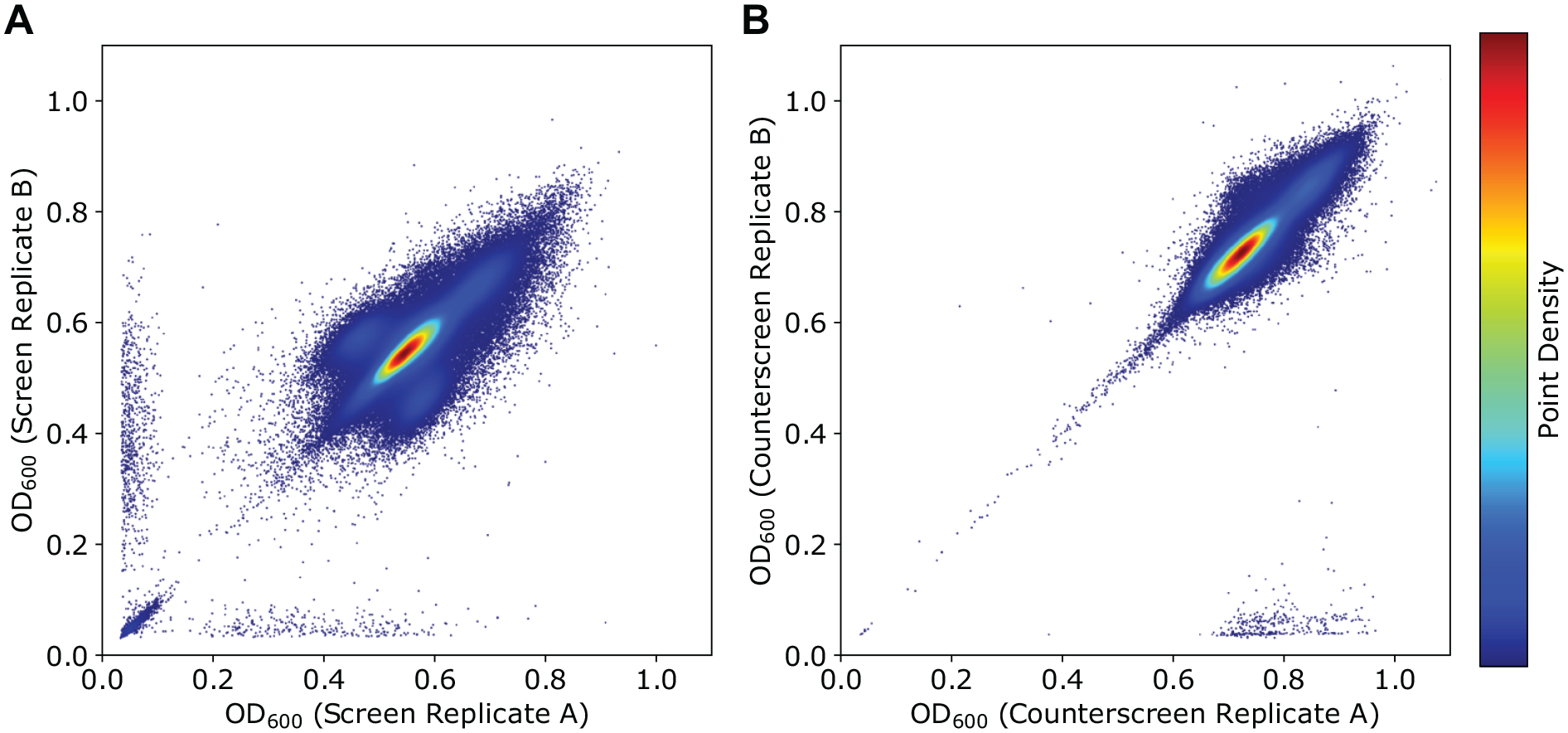
Correlation between high-throughput screen and counterscreen. Assays were performed in duplicate. Screening wells contained 10 µg mL−1 of meropenem; counterscreening wells did not contain antibiotic. Readout was the OD600 of microwells after a 48 h incubation. The values for each pair of duplicate measurements were plotted on x- and y-axes for the screen (
Hit Identification
We initially identified 1531 (0.84% of total compounds screened) total adjunctive and direct antimicrobial screening hits. Of the adjunctive hits, 605 (0.332%), 599 (0.328%), and 43 (0.02%) were weak, medium, and strong, respectively. Of the direct antimicrobial hits, 205 (0.11%), 71 (0.04%), and 8 (0.004%) were weak, medium, and strong, respectively. The z-score distribution for the screen/counterscreen is graphically summarized in Figure 3 .
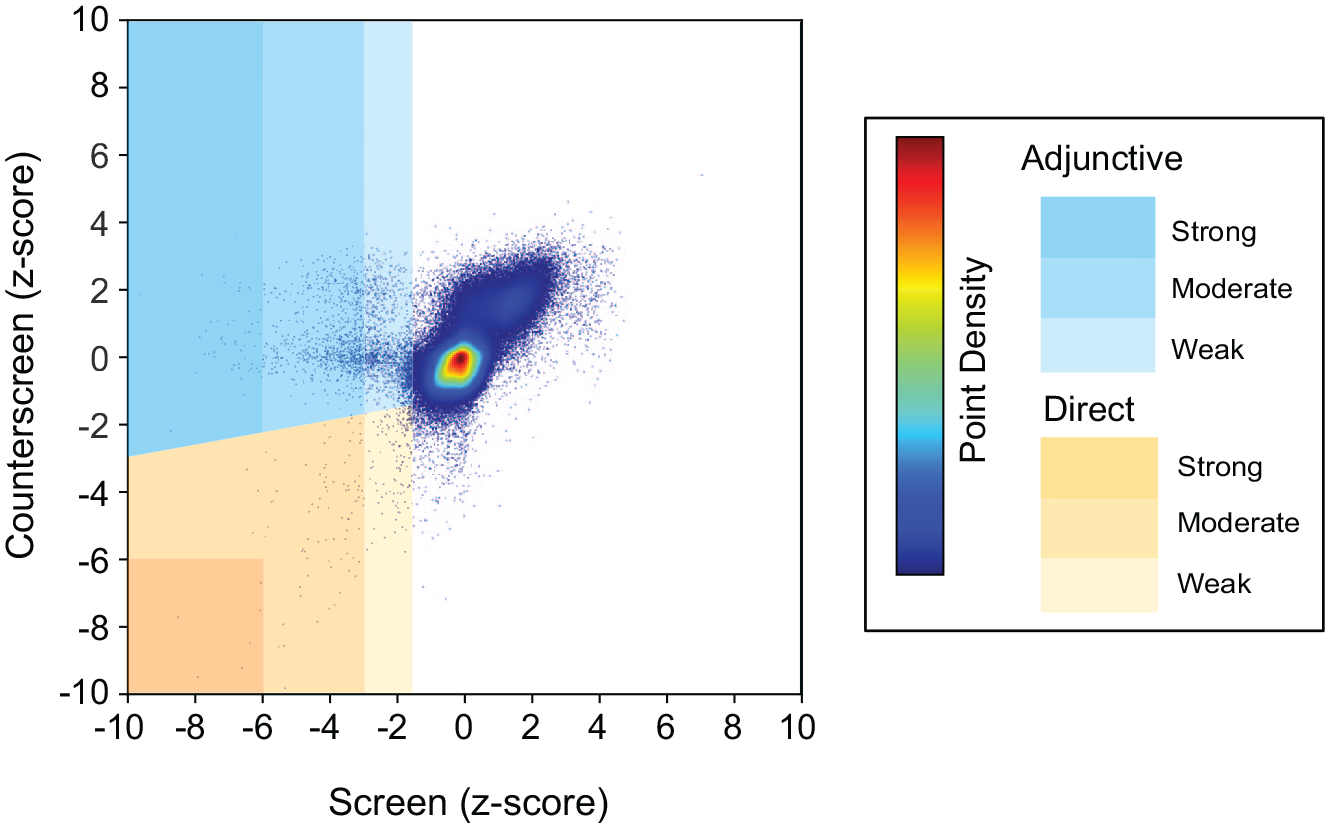
Plot of least significant z-scores for duplicate compound testing in the screen and counterscreen. Z-criteria hit ranking (strong, medium, and weak) is represented in shades of yellow (direct antimicrobials) or blue (adjunctive antimicrobials). Higher relative data point density is represented by warmer colors, as indicated in the legend.
We previously established that our screening assay yielded a high false-positive rate based on the z-score criterion alone and that those hits demonstrating <50% inhibition (compared to control wells) were unlikely to confirm in secondary analysis. 15 Accordingly, we applied a potency requirement of >50% inhibition for at least one of the duplicate measurements. After applying this filter, 439 (72.6%) weak, 598 (99%) medium, and all strong adjunctive hits and 20 (9.8%) weak, 52 (73.2%) moderate, and all strong direct hits were retained.
In addition, we filtered out compounds that showed cytotoxicity to eukaryotic cells, a marker for nonspecific activity, or a target shared by both prokaryotes and eukaryotes, which as a consequence would not be druggable. Eukaryotic cytotoxicity data were from a previously described screening assay using the same compound libraries. 16 After applying this filter, 252 (57.4%) weak, 375 (62.7%) moderate, and 31 (72.1%) strong adjunctive hits were retained; 9 (45%) weak, 31 (59.6%) moderate, and no strong direct antimicrobial hits were retained.
Hit Confirmation
We selected 274 filtered adjunctive and direct antimicrobial hits based on primary screening potency for confirmatory testing using cherry picks from commercial library plates in a manner identical to that of the primary screening assay. Here, we set a less stringent 25% inhibition cutoff in recognition that hits may not recapitulate activity exactly on secondary analysis. In total, 127 (44.2%) adjunctive hits and no direct antimicrobial hits confirmed on retesting.
Cheminformatics Triage
Cheminformatics filtering was then performed to remove nonspecific PAINS with features of covalent modifiers (for example, electrophiles such as aldehydes, ketones, or boronic acids) or metal binders (for example, hydroxamic acids or phosphonates). Even though many antibacterial drugs contain such reactive structural features (approximately 58% of known antibiotics from our testing set fail the PAINS filter), PAINS are considered to be problematic for hit-to-lead optimization and drug development.26,31 Therefore, of the 127 confirmed adjunctive hits, 20 compounds were identified as PAINS and excluded from further analysis. The remaining 107 were clustered based on common substructure, which resulted in identification of 15 clusters and 17 singletons. Further prioritization within clusters was performed based on a compound activity profile and scoring of physicochemical properties characteristic of known antibacterials, using a cheminformatic pipeline called PEMPO that was described in the Materials and Methods section. Select singletons were removed after visual inspection because of limited synthetic tractability or the presence of unfavorable functional groups known to possibly pose bioavailability limitations, narrowing our future analysis to 42 compounds representing 15 clusters and 6 singletons. PEMPO and MPO scoring for these compounds is shown in Supplemental Material 3c. Representative structures of top-scoring clusters and singletons are shown in Figure 4 .
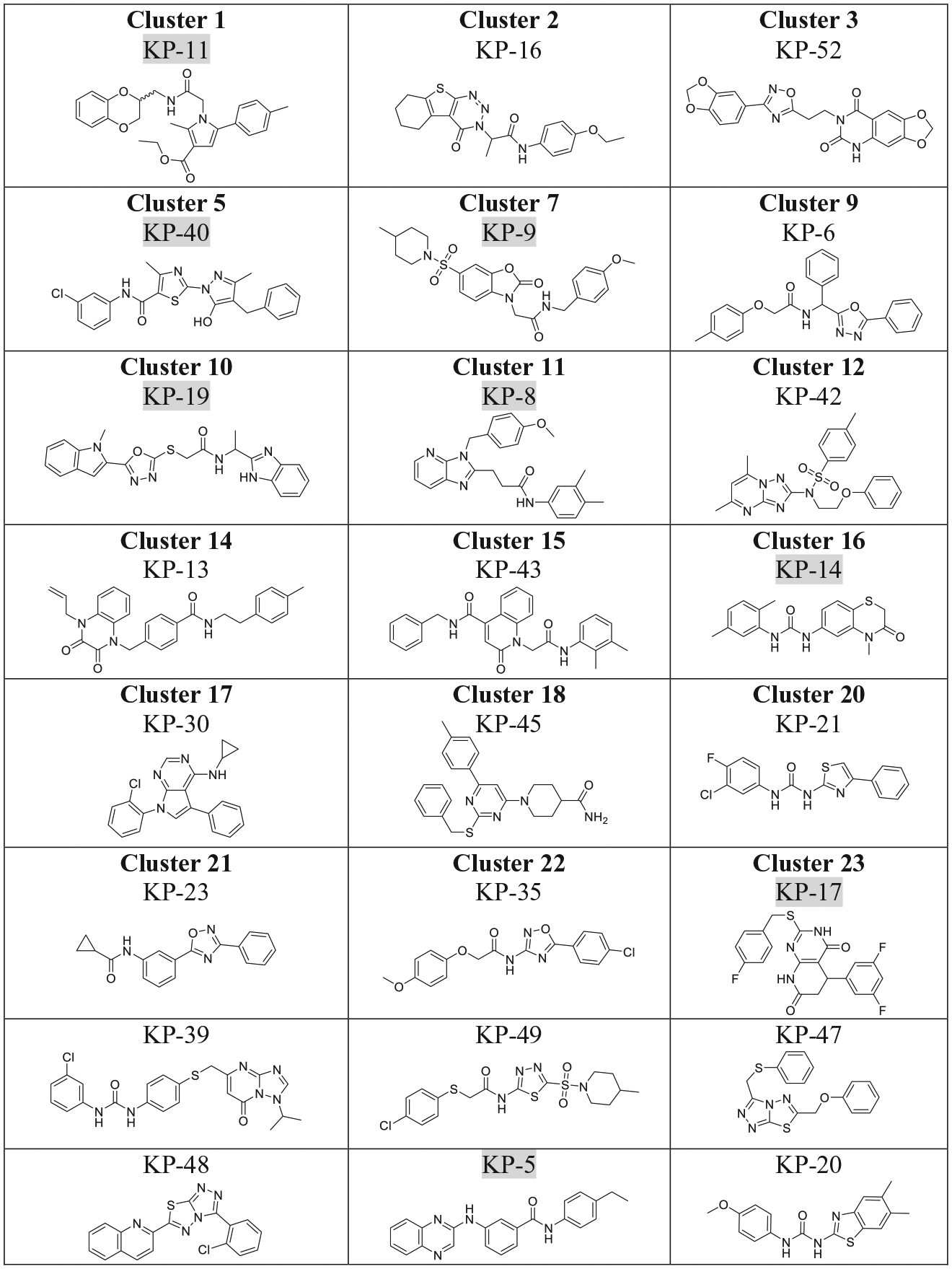
Representative structures of clusters and selected singletons identified by filtering and permeation and efflux multiparameter optimization (PEMPO) analysis. For clusters, representative structures shown are the highest PEMPO scoring compounds within each cluster. Compounds highlighted in gray demonstrated a synergistic adjunctive activity against representative carbapenem-resistant Enterobacteriaceae (CRE) strains after repurchase from commercial suppliers.
Secondary Analysis Using Commercially Synthesized Compounds
We ordered these 42 compounds from commercial suppliers and performed synergy assays in combination with meropenem using our primary screening strain. In total, 23.8% (n = 10) had evidence of synergistic activity with meropenem (FIC ≤ 0.5).
All compounds with an FIC ≤ 0.5 were tested for activity spectrum against a panel of CRE strains consisting of E. coli and K. pneumoniae containing either KPC2 or KPC3 carbapenemases. All selected compounds had activity against ≥50% of strains tested ( Table 1 ).
Spectrum of Activity of Commercially Synthesized and Resynthesized Compounds.
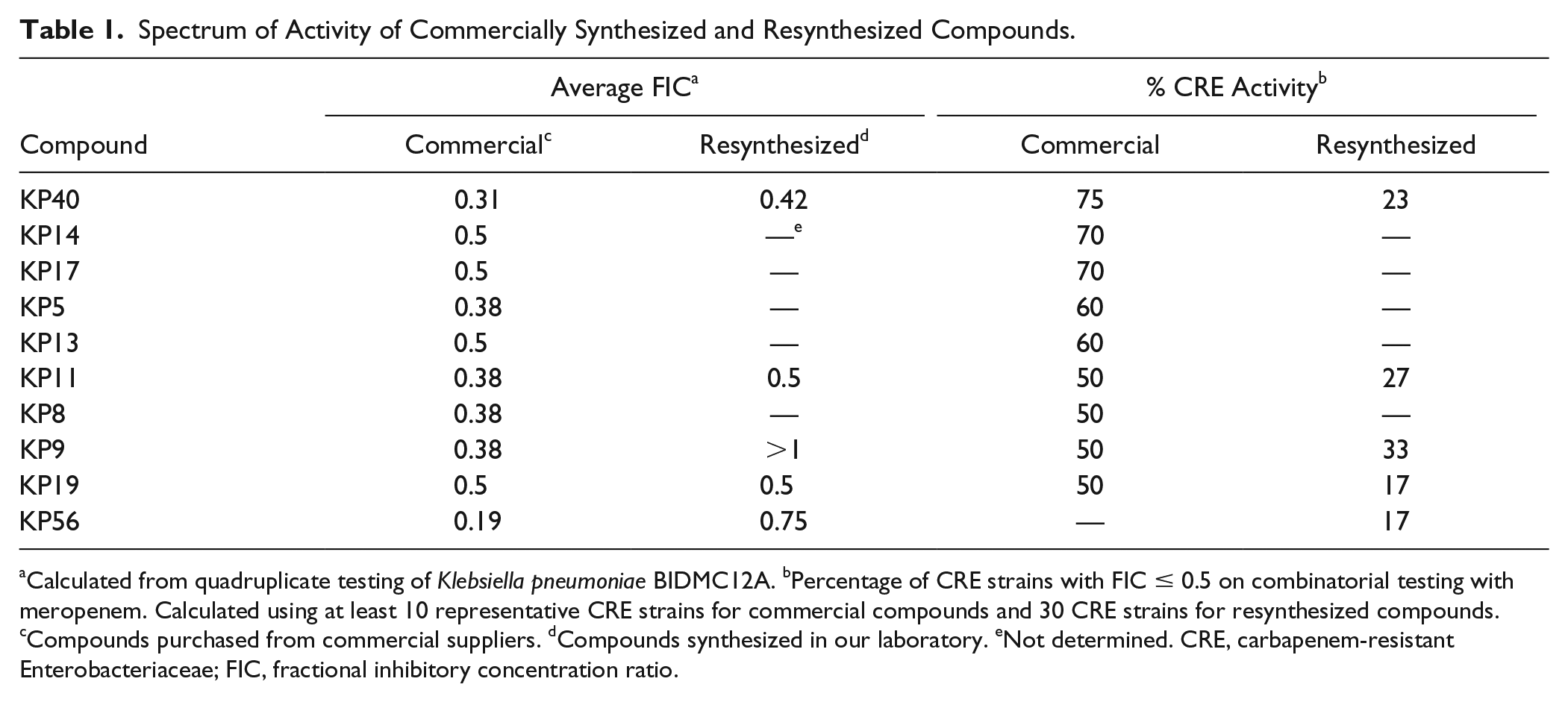
Calculated from quadruplicate testing of Klebsiella pneumoniae BIDMC12A. bPercentage of CRE strains with FIC ≤ 0.5 on combinatorial testing with meropenem. Calculated using at least 10 representative CRE strains for commercial compounds and 30 CRE strains for resynthesized compounds. cCompounds purchased from commercial suppliers. dCompounds synthesized in our laboratory. eNot determined. CRE, carbapenem-resistant Enterobacteriaceae; FIC, fractional inhibitory concentration ratio.
Analysis of Resynthesized Compounds
To this point, we had been using compounds available in limited quantities from the commercial suppliers of our screening libraries. We added an additional layer of confirmation by resynthesizing hit compounds and confirming structural identity and purity by liquid chromatography with diode array detection, mass spectrometry, and nuclear magnetic resonance (NMR) (see Supplemental Materials 6 and 7 for details). We used potency (based on FIC) and spectrum of activity as primary criteria, and predicted physicochemical property data as secondary criteria to select KP40 and KP11 for resynthesis. Other compounds chosen for resynthesis were KP9 and KP19, which displayed excellent PEMPO scores (4.4 and 4.8, respectively), along with KP56, which demonstrated good activity in primary screening. Following synthesis, these compounds were tested again using our screening strain to confirm activity using our standard synergy assay.
Of the resynthesized compounds, KP11, KP40, and KP19 demonstrated synergy with meropenem against our screening strain ( Table 1 ). Two compounds, however—KP9 and KP56—did not. We then tested resynthesized compounds against our 30-strain CRE panel. Resynthesized compounds showed synergy against 17 to 33% of CRE strains. KP9 and KP56, while not showing synergy against the screening strain, demonstrated synergy against a subset of clinical CRE strains.
Synergy Testing in a Non-CRE Background
Confirmed adjunctive hits might interfere with carbapenemase activity or alternatively affect the physiology of specific bacterial strains to enhance potency of meropenem by other mechanisms. To distinguish phenotypically between these possibilities, we first constructed isogenic E. coli strains expressing the serine carbapenemases (KPC2 or KPC3) or a metallo-carbapenemase (NDM1). The strain background used was DH5α, a laboratory-adapted E. coli K-12 strain with no intrinsic resistance to β-lactams, including meropenem. We found no synergy of compounds with meropenem, however, suggesting that effects might be strain specific, and as a result not effective against E. coli K-12.
We therefore considered whether the lack of activity in the E. coli K-12 background might relate to either lack of permeation and/or efflux. To distinguish between these two possibilities, we introduced our KPC2-, KPC3-, and NDM1-containing plasmids into E. coli strains with defects in the outer-membrane permeability barrier (lptD) or efflux activity (tolC). The lptD mutant expresses a truncated form of LptD, a protein critical in transporting lipopolysaccharide (LPS) to the outer membrane. 32 The resulting deficit in LPS in the outer membrane leads to increased permeability. The tolC mutant inactivates a critical shared component of several efflux pumps. 33 We did not observe synergy with meropenem in either strain. We did, however, observe direct antimicrobial activity of KP40 in the tolC mutant. MICs were independent of carbapenemase production and ranged from 16 to 128 µM during replicate testing, an unusual degree of biological variability not typical of established antimicrobials with specific mechanisms of action (Suppl. Material 5a–c).
Cheminformatic Characterization of HTS Libraries
The physicochemical properties of screening libraries used in this effort were characterized and compared to the chemical space occupied by 100 known Gram-negative antimicrobials. Molecular weight, polar surface area, cLogD7.4, and the summation of HBDs and HBAs of screening compounds and known antimicrobials were calculated. Plots of physicochemical properties versus molecular weight are shown in Figure 5 .
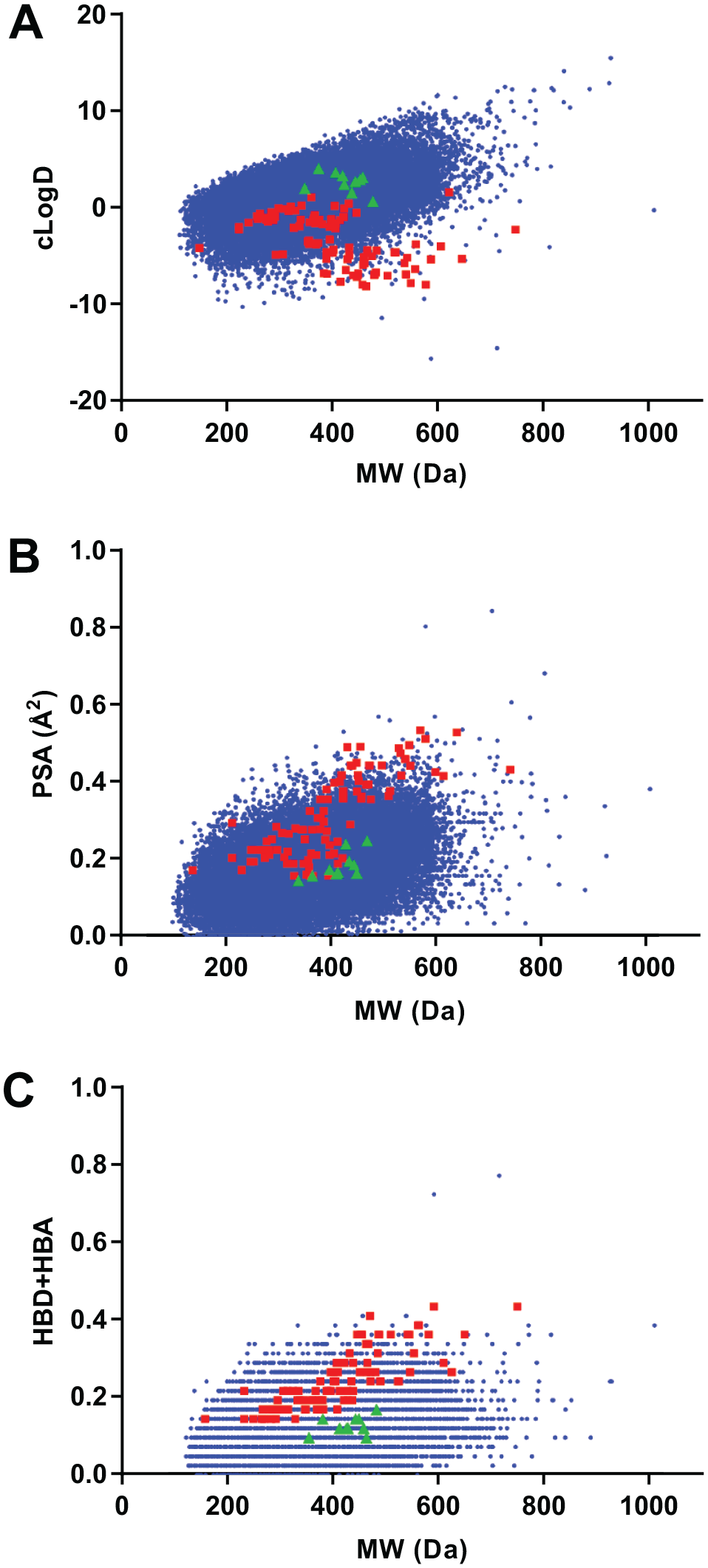
Cheminformatic analyses of screening libraries. (
From data plots, it is apparent that the screening library consists of compounds with a greater degree of lipophilic substituents. More specifically, partition coefficients (cLogD7.4) for library compounds demonstrate increasing lipophilicity with increasing molecular weight ( Fig. 5A ). In contrast, partition coefficients of 100 known Gram-negative active antimicrobials show the opposite trend. Similar but inverted trends for the screening libraries and known antimicrobials were observed in plots of polar surface area ( Fig. 5B ) as well as summations of HBDs and HBAs ( Fig. 5C ).
Discussion
A screen of commercially available small-molecule libraries was performed to identify carbapenem potentiators and direct antimicrobial inhibitors of a representative K. pneumoniae carbapenem-resistant clinical isolate. The goal was to identify hits that could be further improved by using medicinal chemistry approaches. Furthermore, the whole-cell screening approach was agnostic regarding the potential mechanism of action, which could be further delineated at a later time for promising scaffolds.
Hits underwent initial triage based on combined use of cheminformatics approaches and data from an orthogonal screen to eliminate eukaryotic cell toxic compounds. Based on this initially stringent but likely appropriate down-selection, only a few hits with potentiating activity and no hits with direct activity remained. Many, but not all, of these adjunctive hits retained activity on resynthesis. The lack of complete reproducibility on resynthesis is a well-known finding in commercial library screening efforts and may result from contaminants such as heavy metal catalysts, which may confer antimicrobial activity unrelated to the compound being studied. In addition, we observed reduction in activity spectrum of several of the compounds. Reasons for this are not immediately obvious, but may relate to borderline adjunctive activity, which did not reach a threshold for phenotypic detection with the resynthesized compound. It may also represent contributions of both compound and contaminants in the original commercial preparations that differed from resynthesized compounds.
We hypothesized that a subset of potentiators would represent hits that either directly or indirectly targeted carbapenemase activity. Tests in isogenic E. coli strains expressing several types of carbapenemases, however, failed to detect synergy with meropenem, suggesting effects were specific to only a subset of clinical strains being tested based on shared regulatory and/or biophysical characteristics, potentially a reflection of the diversity of the CRE strain set.
To address target access, we tested previously well-characterized E. coli K-12 strains with known defects in either their permeability barrier (lptD) or a major class of efflux pumps (tolC). Neither strain allowed observation of carbapenem potentiation in the E. coli K-12 background, suggesting that differences other than efflux or outer-membrane permeability barrier accounted for the observed activity spectrum. One compound, KP40, was noted to have direct, but highly variable, antimicrobial activity against the tolC mutant; this high biological variability suggests nonspecific interference with bacterial growth, that is, hitting multiple targets with total assay variability reflecting the sum of the variability of multiple events.
Our goal was to identify compounds that had already passed the high bar for activity against a multidrug-resistant pathogen. In that way, there would be a stringent biological triage with hopes of later improving initial activity using medicinal chemistry approaches. Our clinical screening strain is known to encode multiple antimicrobial resistance elements, 17 and it has a very high baseline carbapenem MIC (50 µg mL−1) that is 16- to 32-fold higher than in a laboratory E. coli strain expressing the same carbapenemase gene (data not shown). Therefore, the carbapenem resistance phenotype observed in this and other clinical isolates is likely complex and polygenic, with contributions from efflux pumps, altered porins, membrane and cell wall characteristics, and/or β-lactamases with low-level ability to hydrolyze carbapenems.
Unfortunately, compounds with compelling direct or adjunctive antimicrobial activity were not identified through these efforts. One potential explanation for this is that the biological triage was too stringent. For example, we did not follow up on compounds that demonstrated statistically significant cytotoxicity for J774A.1 macrophages observed in a separate screening effort. Therefore, it remains possible that some compounds identified as eukaryotic cell toxic may have had some degree of selectivity for bacteria that could have been improved during structure–activity relationship studies.
Another possibility is that the commercial screening libraries available did not contain sufficiently diverse compounds with physicochemical properties conducive to Gram-negative antimicrobial activity. For example, a prior screening effort of 500,000 compounds at GlaxoSmithKline against an efflux-competent strain of E. coli yielded no confirmed hits. 34 This finding was attributed to lack of chemical diversity. Although the chemical space occupied by the libraries examined was not reported, it is well-known that commercial and pharmaceutical libraries historically have been optimized for “druglike” molecules based on metrics such as Lipinski’s rule of five. 35
Antimicrobials in general and Gram-negative agents in particular, however, rarely satisfy these rules 30 . In analyses of the physicochemical properties of our screening libraries and representative hits, we observed trends suggesting that compounds with characteristics of Gram-negative antimicrobials were underrepresented. Gram-negative antimicrobials typically possess zwitterionic or polar moieties, which facilitate passage of compounds through water-filled transmembrane porins and entry into the periplasm. Our libraries and screening hits had, however, a paucity of such compounds. Instead, they were enriched for compounds with lipophilic substituents (high cLogD), decreased numbers of HBDs and HBAs, and decreased polar surface area, which face an increased enthalpic barrier for entry into the Gram-negative cell. Compounds with these attributes are generally less challenging to synthesize and purify, and therefore not unexpectedly are overrepresented in screening libraries.
An alternative screening approach using a screening strain with a lower barrier for activity may have been more productive in identifying lead candidates. For example, a screen of 150,000 small molecules using fully antimicrobial-susceptible E. coli and Pseudomonas aeruginosa strains identified several confirmed hits with weak activity. 36 To the best of our knowledge, however, further development of novel compounds from this screen has not been described. Additional perturbation, such as the use of tolC or lptD mutants, may further lower the bar for Gram-negative inhibitor detection 37 but later require additional chemistry efforts to address efflux and permeability effects that may or may not prove productive.
Taken together, our results support previous observations that Gram-negative antimicrobial lead candidates may be largely absent from commercially available screening libraries. Certainly, this appeared to be the case for compounds with intrinsic activity against a highly multidrug- and carbapenem-resistant K. pneumoniae clinical strain. Although we were able to find detectable activity for several compounds, the overall potency was low. Therefore, we provide further data that whole-cell screening efforts for Gram-negative antimicrobials should be conducted using libraries with diverse scaffolds and substituents outside the Lipinski space, including, for example, compounds with greater hydrophilicity.
Further supporting this view, our prior efforts using the same screening strategy to examine known bioactive libraries with higher diversity inclusive of natural products were highly productive. This led to identification of apramycin and several nucleoside analogues as lead direct antimicrobial candidates for development against highly drug-resistant CRE and (for the former) against multidrug-resistant Acinetobacter baumannii, for which apramycin is now recognized more generally as a candidate for preclinical development.38–40 In this earlier screening effort, we also identified potent meropenem adjunctive activity of triclosan, confirming the underlying ability of the screening strategy to detect both direct and adjunctive antimicrobials. 15 Therefore, the fundamental ability of the whole-cell, HTS assay to detect antimicrobials with activity against CRE—and by extension other, multidrug-resistant, Gram-negative pathogens of concern—offers promise as libraries with appropriate physicochemical properties become available.
Supplemental Material
Supplemental_Material_for_A_Whole_Cell_Screen_for_Adjunctive_and_Direct_Antimicrobials_Active_Against_Carbapenem-Resistant_Enterobacteriaceae_by_Smith_et_al – Supplemental material for A Whole-Cell Screen for Adjunctive and Direct Antimicrobials Active against Carbapenem-Resistant Enterobacteriaceae
Supplemental material, Supplemental_Material_for_A_Whole_Cell_Screen_for_Adjunctive_and_Direct_Antimicrobials_Active_Against_Carbapenem-Resistant_Enterobacteriaceae_by_Smith_et_al for A Whole-Cell Screen for Adjunctive and Direct Antimicrobials Active against Carbapenem-Resistant Enterobacteriaceae by Kenneth P. Smith, Matthew G. Dowgiallo, Lucius Chiaraviglio, Prakash Parvatkar, Chungsik Kim, Roman Manetsch and James E. Kirby in SLAS Discovery
Footnotes
Acknowledgements
We would like to thank Jennifer Smith, Jennifer Nale, David Wrobel, Stewart Rudnicki, Rachel Warden, and Richard Siu (ICCB-Longwood Screening Facility, Harvard Medical School, Boston, MA) for their assistance. The HP D300 digital dispenser was provided for our use by Tecan (Morrisville, NC). Tecan had no role in study design, data collection or interpretation, manuscript preparation, or decision to publish.
Supplemental material is available online with this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: K.P.S. and J.E.K. were supported by the National Institute of Allergy and Infectious Diseases of the National Institutes of Health under award numbers F32AI124590, and R21AI119114 and R33AI119114, respectively. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. The screening work was performed at ICCB-Longwood Screening Facility (Boston, MA).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.