
Abstract
Successful oral immunization depends on efficient delivery of antigens (Ags) to the mucosal immune induction site. Glycoprotein-2 (GP-2) is an integral membrane protein that is expressed specifically on M cells within follicle-associated epithelium (FAE) and serves as transcytotic receptor for luminal Ags. In this study, we selected peptide ligands against recombinant human GP-2 by screening a phage display library and evaluated their interaction with GP-2 in vitro and ex vivo. Selected peptides were conjugated to the C-terminal of enhanced green fluorescence protein (EGFP) and evaluated for their ability to induce an immune response in mice. One of our selected peptides, Gb-1, showed high binding affinity to GP-2 and, when fused to EGFP, significantly increased the uptake of EGFP by M cells compared to EGFP alone. After oral administration, the Gb1-EGFP fusion induced efficient mucosal and systemic immune responses in mice measured at the level of antigen-specific serum and fecal antibodies, cytokine secretion, and lymphocyte proliferation. Furthermore, the IgG subclasses and cytokine secretion showed that ligand Gb-1 induced a Th2-type immune response. Collectively, our findings suggest that the ligand we selected through phage library screening is capable of targeting Ags to GP-2 on M cells and can be used as an oral vaccine adjuvant.
Introduction
The mucosal surface of the gastrointestinal and respiratory tract is continuously exposed to microorganisms, and a large number of pathogens invade the body through the mucosal surface. 1 Gut-associated lymphoid tissue (GALT) serves as sentinel for the recognition of intestinal microbes and initiation of immune responses. 2 Theoretically, mucosal immunization is superior to systemic immunization since it elicits immune responses at systemic and mucosal levels. 3 Consequently, it seems logical to develop oral vaccines. Oral vaccination also offers additional advantages over systemic administration such as convenience of administration, low cost, and needle-free application. 4 However, there are challenges to the oral application of vaccines, and only a few oral vaccines are currently available. Most of the currently available vaccines are administered through the systemic route and induce an immune response predominantly in the systemic compartment.5,6 Difficulty in efficient delivery of antigens to the mucosal immune induction site is one of the main obstacles in the development of oral mucosal vaccines. 7 Consequently, devising new strategies to enhance the efficiency of antigen delivery to mucosal lymphoid tissue and to avoid possible oral tolerance induction is of great importance for the development of successful mucosal vaccines.
The luminal side of the GALT lymphoid follicles is covered by follicle-associated epithelium (FAE). M cells are specialized cells in the FAE that actively transport luminal antigens and macromolecules across the epithelial membrane, a process referred to as transcytosis. 8 The basal plasma membrane of M cells forms a pouch-like structure called the “M-cell pocket,” where dendritic cells (DCs) and lymphocytes migrate. Although M cells appear to be ideal targets for eliciting Ag-specific immune responses through oral vaccination, there are challenges to using these cells for Ag delivery. 9
Efficient uptake of antigens by M cells requires specific surface receptor molecules. Identification of M-cell–specific markers made it possible to target these cells for antigen (Ag) delivery to improve vaccine efficacy. Glycoprotein-2 (GP-2) is a glycosylphosphatidyl inositol anchored protein that is specifically expressed on M cells and serves as transcytotic receptor for intestinal Ags. Also, GP-2 was shown to be associated with specific uptake of FimH
Phage display library is a powerful tool for screening peptide ligands against proteins and other macromolecules both in vitro and in vivo and has been used in basic and applied research for studying molecular biology mechanisms involving protein-protein interactions. 11 In this study, we used a phage display library to screen short peptide ligands against the transcytosis receptor GP-2. The affinity of the selected ligands to bind to GP-2 and their ability in immune induction were assessed in mice. We selected three peptide ligands, hereafter called GP2-binding peptides (Gbs), of which one peptide, Gb-1, showed significant results in immune induction. Gb-1 fusion increased the uptake of Ag by M cells through GP-2 and elicited significantly high levels of serum IgG and mucosal IgA, as well as cytokines secreting cells in various lymphoid tissues, especially interleukin (IL)–4, IL-5, and IL-6, which are associated with isotype switching to secretory IgA. Our results suggest that peptide Gb-1 selected through biopanning has immune-stimulating ability and can be used as an adjuvant for mucosal vaccines.
Materials and Methods
Phage Library Biopanning
The 12-mer peptides phage display library of filamentous phage M13KE was biopanned according to the guideline provided by the vendor (New England BioLabs, Ipswich, MA). Briefly, 5 µg recombinant GP-2 (ProsPec Bio, Ness-Ziona, Israel) was adsorbed to a 96-well plate (Corning, Corning, NY) at 4 °C. After overnight incubation, wells were blocked with 5% bovine serum albumin (BSA) in H2O for 2 h at 4 °C. Approximately 1 × 1011 phages were added to the GP-2–coated wells after washing with TBST (.05% Tween 20) and incubated for 2 h at room temperature (RT). The loosely/nonspecifically bound phages were washed and the strongly bound phages were eluted with 100 µL of 0.2 M glycine-HCl (pH 2.2) and 1 mg/mL BSA. The rescued phages were amplified in Escherichia coli ER2738, titred on isopropyl β-D-1-thiogalactopyranoside (IPTG)/Xgal plates, and a known titer of the amplified phages was used for next round of screening. After four rounds of biopanning, DNA was extracted, quantitated on agarose gel by comparing with 0.5 µg of purified single-stranded M13mp18 DNA (NEB 4040), and sequenced. Amino acid sequences were deduced according to the vendor instructions using Chromas Lite software (Technelysium, Brisbane, Australia).
The resulting amino acid sequences were scanned using SAROTUP (http://immunet.cn/sarotup/index.html) to check whether they match any known target-unrelated peptide (TUP) motif or if any submitted peptide has also been selected by other groups with other targets or to confirm that the phage clones achieved in the biopanning results are without any propagation advantage and are true binders to the target. Insight II program (Molecular Simulations, Inc., San Diego, CA, USA) was used to align the amino acid sequences and find regions of structural conservation among the selected peptides.
Phage-Binding Enzyme-Linked Immunosorbent Assay
To identify high-affinity binding clones, 96-well enzyme-linked immunosorbent assay (ELISA) plates were coated with 150 µL GP-2 (5 µg/mL) in 0.1 M NaHCO3 (pH 9.6) at 4 °C overnight. Plates were blocked with 5% BSA in 0.1 M NaHCO3 (pH 9.6) at 4 °C for 2 h. Then, 100 µL phage clones (1 × 1010 pfu/well) was added and incubated at RT for 1 h. After washing, horseradish peroxidase (HRP)–conjugated anti-M13 antibody (1:5000 dilution) was added and incubated at RT, followed by TMB substrate incubation. Finally, the reaction was stopped using 2 M H2SO4, and plates were read at 450 nm using a SpectraMax M5 (Molecular Devices, Sunnyvale, CA) microplate reader. Phage clones giving target-to-background absorbance >4 were selected for further assays.
Peptides Synthesis
The deduced 12-mer peptides were synthesized by standard Fmoc method (China Peptides Co., Shanghai). The N-terminal of peptides was kept free while the C-terminal was amidated and conjugated to fluorescein isothiocyanate (FITC) using the GGGS linker.
Measurement of Peptide-GP2 Binding Affinity (K d Values)
To measure the binding strength of the selected peptide ligands to GP-2, saturation binding assays were performed as described. 12 Then, 150 µL (5 µg/mL) GP-2 in assay diluent (BioLegend, San Diego, CA) was adsorbed to 96-well plates at 4 °C overnight. The solution was discarded and wells were blocked with 5% BSA in water for 2 h at 4 °C. After washing, wells were subjected to incubation with different concentrations of FITC-conjugated peptides for 1 h, followed by incubation with HRP-conjugated anti-FITC antibody (1:5000 dilution). The reaction was visualized with 100 µL HRP substrate and OD450 was measured. Specific binding of each peptide to GP-2 was calculated by subtracting the OD value of each peptide binding to BSA-coated wells from the OD value of that peptide binding to GP-2. Assays were performed three times in duplicate. Binding affinity of each peptide was calculated by nonlinear regression and transformed to Scatchard plot using the GraphPad Prism 6 program (GraphPad Software, La Jolla, CA).
Next, we performed competition ELISA to investigate the epitope binding sites on GP-2 using the above method.
Production of Peptide-Conjugated Recombinant Ag
We used enhanced green fluorescence protein (EGFP) as a model antigen and fused selected peptides to the C-terminal of EGFP. The EGFP gene was amplified by PCR from pEGFP-C1 (Clontech, Mountain View, CA, USA) using primers F: 5′-GTGAGCAAGGGCGAGGAGCTG-3′ and R: 5′-CTTGTA CAGCTCGTCCATGCCG-3′. Fusion of DNA sequences of the selected peptides and insertion of the resulting construct into pET-28a was achieved by our newly introduced single primer–mediated circular PCR method 13 using primers as shown in Table 1 . Protein was expressed in BL21 (DE3) and purified on HisTrap FF columns (GE Healthcare Bio-Sciences, Uppsala, Sweden) according to the manufacturer’s instructions. Purified protein was confirmed by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and Western blot using anti-HisTag and anti-EGFP antibodies (Abcam, Shanghai, China).
Primers Used for Fusion of Peptides to Enhanced Green Fluorescent Protein and Plasmid Circularization.

Primers used for circularization of linear constructs.
Ex Vivo Ag Uptake Assay
Ex vivo Ag uptake of the selected ligands by M cells was assessed by gut loop assay as described previously. 14 A gut loop containing one or two Peyer’s patches (PPs) was prepared from overnight fasted mice and 0.2 mg of recombinant Ag-fused EGFPs or EGFP alone was injected into the ligated segment. After 1-h incubation at 37 °C, the ligated segment was excised, washed with phosphate-buffered saline (PBS) to remove the internal contents, and fixed with 4% paraformaldehyde (PFA) For whole-mount staining, PPs were stained with anti-GP2 (Abcam, Cambridge, UK) followed by allophycocyanin-conjugated anti-rabbit IgG, whole mounted with antifade medium. Frozen sections (10 µm) were prepared after freezing PPs in OCT (Takara, Shiga, Japan), blocked with 2.5% BSA and stained with anti-GP2, counterstained with 4′,6-diamidino-2-phenylindole (DAPI), and analyzed by confocal laser scanning microscopy (CLSM) (LSM 710; Carl Zeiss, Thornwood, NY, USA).
Mice Immunization and Measurement of Ag-Specific Immune Responses
Groups of five female BALB/c mice between 4 and 6 weeks of age were immunized with 100 µg of experimental antigen/PBS by oral gavage once every week for 6 weeks. 15 Five days after the last immunization, serum and fecal extracts were collected to monitor EGFP-specific systemic IgG and mucosal IgA as described by Hackett et al. 16 Since IgA in fecal extracts is rapidly degraded by the proteases from enteric bacteria, 1 mM phenylmethyl sulfonyl fluoride (PMFS) protease inhibitor was included in the extraction cocktail. 17 Ab titers were expressed as the reciprocal log2 of the highest sample dilutions that gave an OD450 of 0.08, which was the value of the PBS blank.
Lymphocytes from spleen (SPLs) and PPs (PPLs) were isolated, minced, and digested with 300 U/mL Collagenase D (Roche, Mannheim, Germany) for 30 min at 37 °C. The digested mixture was passed through a nylon mesh to remove undigested tissue and subjected to Percoll (Sigma-Aldrich, St. Louis, MO) density gradient centrifugation. Cells at the interface between 40% and 75% Percoll were collected as mononuclear cells and subjected to characterization for lymphocyte proliferation and cytokine secretion.
The level of cytokine secretion was determined by cytokine-specific ELISAs with culture supernatants from lymphocytes restimulated with the Ags for 24 h as described previously. Cytokine concentrations were calculated from the plotted standard curves of serial dilutions of the recombinant cytokine and expressed as mean ± standard error (SE) (ng/mL) of each group. To measure the number of antigen-specific cytokine (interferon [INF]–γ, interleukin [IL]–4, IL-5 and IL-6) secreting cells, enzyme-linked immunospot (ELISPOT) assays were performed with lymphocytes isolated from spleens and PPs as described previously. 15
Ag-Specific Lymphocyte Proliferation
Ag-specific lymphocyte stimulation was determined using Dojindo CCK-8 reagent (Dojindo Laboratories, Kumamoto, Japan). 18 Purified lymphocytes were plated in flat-bottom 96-well plates at a density of 5 × 105 cells per well and stimulated with recombinant EGFP (1 mg/mL) or PBS (negative control) at 37 °C in a 5% CO2 incubator. Plates were incubated for 72 h and pulsed with 10 µL CCK-8 reagent (Dojindo Laboratories) per well for another 4 h. Absorbance was measured at 450 nm, and the stimulation index (SI) was calculated as the ratio of the average OD450 value of wells containing antigen-stimulated cells to the average OD450 value of wells containing cells stimulated with PBS. All assays were performed in triplicate.
Statistical Analysis
The results are expressed as mean ± SE (SEM) using Microsoft Excel 2016 (Microsoft Corp., Redmond, WA), and at least three independent experiments were performed unless otherwise stated. An unpaired Student t test (two-tailed) was used to compare groups, and p values <0.05 were considered statistically significant.
Results
Biopanning of Phage Library
To select peptide ligands against mucosal transcytotic receptor GP-2, we screened a 12-mer peptide phage display library against recombinant GP-2 as described in Materials and Methods. Four rounds of biopanning were performed, and comparatively stringent conditions were applied in each round to ensure enrichment in favor of GP-2. Phage titer increased more than 4000-fold from the first round to the fourth round ( Table 2 ). The overall enrichment of the library suggests that enrichment for selectively binding phage was achieved. After four rounds of biopanning, 3.7 × 105 phages were recovered and 50 plaques were randomly picked for sequence determination. From 50 plaques, we obtained seven different amino acid sequences and named them GP-2 binding peptides (Gb-1 to Gb-7), of which three sequences appeared more frequently than the others and were selected for further characterization. SAROTUP scanning results showed that none of these sequences have been reported before for other targets or contain any known TUP motif or confer propagation advantages. Alignment analysis using Insight II showed that no conserved structural features exist among the selected peptides.
Summary of Phage Library Biopanning against Glycoprotein-2.
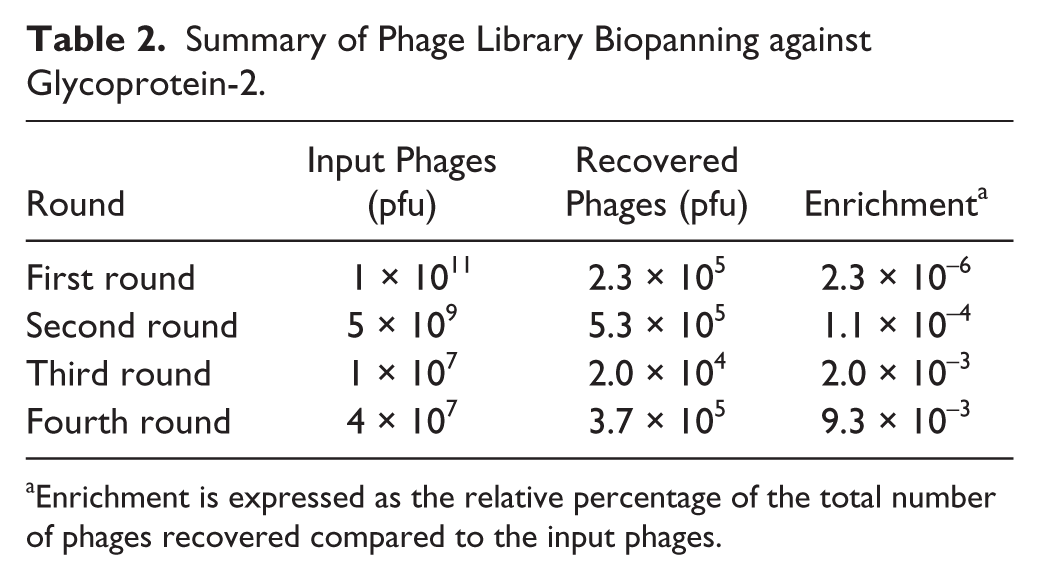
Enrichment is expressed as the relative percentage of the total number of phages recovered compared to the input phages.
Selected Peptides Bind to GP-2 with High Affinity
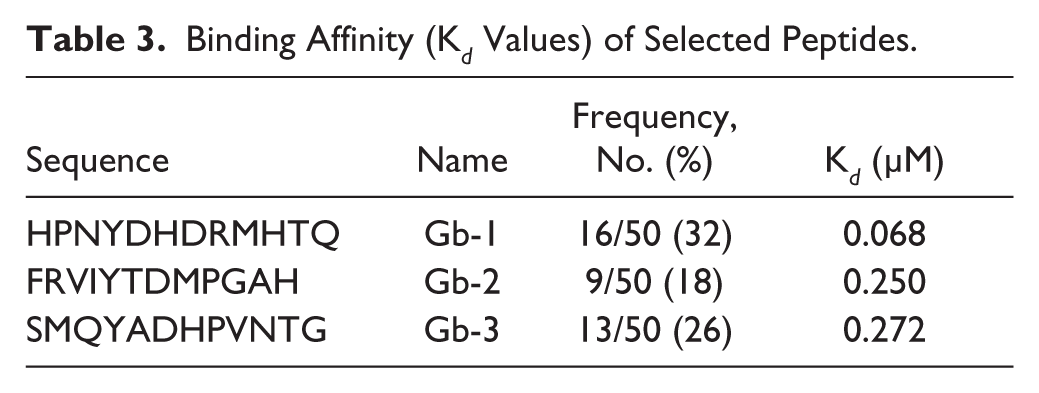
The ability of the selected peptides ligands to bind to GP-2 was measured in terms of dissociation constant (K d ) values of the synthetic peptides. K d values were determined by nonlinear regression analysis and Scatchard transformation using GraphPad Prism 6. The binding of peptides to GP-2 was specific and hyperbolic. All three peptides showed K d values in the nanomolar range: Gb-1 (Kd = 68 nM), Gb-2 (Kd = 250 nM), and Gb-3 (Kd = 272 nM) as shown in Table 3 . The binding pattern of these peptides to GP-2 measured by saturation binding assay was consistent with the binding pattern measured by phage-binding ELISA. Competition ELISA results showed that the selected peptides do not bind to any common or overlapping epitope on GP-2.
Binding Affinity (K d Values) of Selected Peptides.
Binding of Ligand-Fused EGFPs to GP-2 and Transcytosis to M Cells
To test the GP-2 targeting ability of selected peptides, we expressed peptide-fused EGFPs and analyzed their binding to M cells on PPs. We performed ligated gut loop assay and stained whole PPs and frozen sections with anti–GP-2 after incubation with either ligand-conjugated EGFPs or EGFP alone. Immunohistochemical staining of the whole-mount and frozen sections of PPs confirmed the binding of only Gb-1–conjugated EGFP to M cells that specifically express GP-2 ( Fig. 1A , B ). However, Gb-2 and Gb-3 conjugated to EGFP were not preferentially bound to M cells. To confirm the ligand-assisted internalization of EGFPs into PPs, we analyzed and compared the fluorescence intensity of EGFP fluorescence under the dome area of PPs in frozen sections 1 h after oral feeding. We found that ligand-fused EGFPs were internalized into the PPs successfully; in particular, peptide Gb-1 most efficiently transferred its fluorescence into PPs ( Fig. 1C ). However, unconjugated EGFP did not show such internalization. These results show that Gb-1 binds to GP-2 on M cells and can promote Ag transcytosis to PPs.
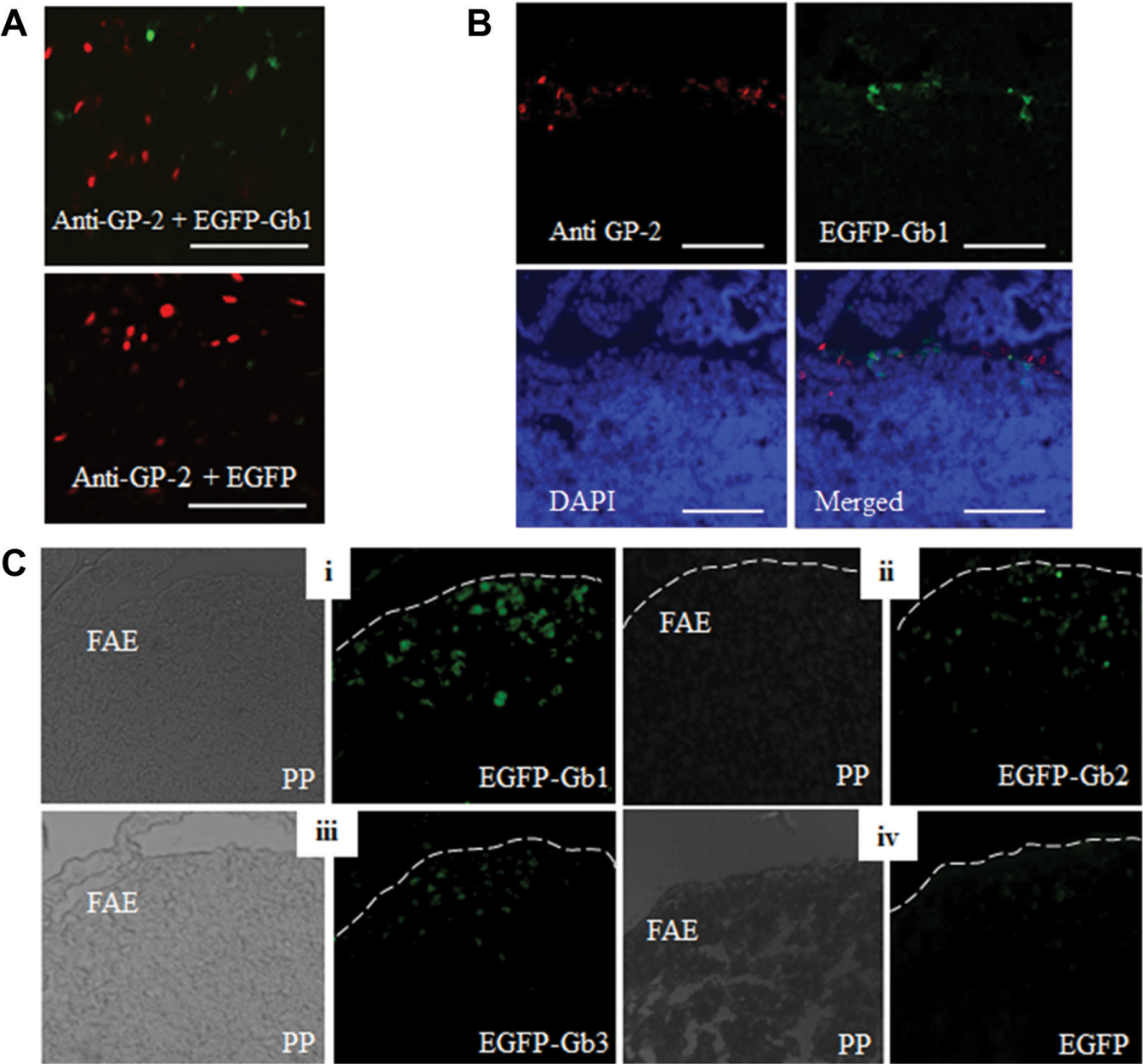
Immunohistochemical analysis of peptide–glycoprotein-2 (GP-2) interaction and internalization to Peyer’s patches (PPs). Interaction of selected peptides with GP-2 and internalization into PPs through M cells was evaluated by immunohistochemical staining of whole-mount specimens and cryosections of mouse PPs 20 min after administration of enhanced green fluorescent protein (EGFP) into ligated loops as described in the Materials and Methods. (
Gb-1 Enhances Ag-Specific Immune Response Induction
To test the ability of the selected peptides to induce immune response, we administered ligand-conjugated or only EGFP to mice orally and evaluated antibody response in systemic and mucosal compartments using ELISA. Immunization with the ligand-fused EGFPs increased the induction of EGFP-specific serum IgG compared to EGFP alone. In particular, Gb-1–conjugated EGFP enhanced the induction of EGFP-specific IgG more efficiently and increased the level of EGFP-specific serum IgG by >2-fold compared to other ligand-fused EGFPs ( Fig. 2A ). The enhancement in immune response was further analyzed at the IgG subclass level. As shown in the Figure 2C , D , a ~2-fold increase in the induction of EGFP-specific IgG1 was detected for Gb-1, but no significant increase in the level of Ag-specific IgG2a could be detected even with the Gb-1–fused EGFP.
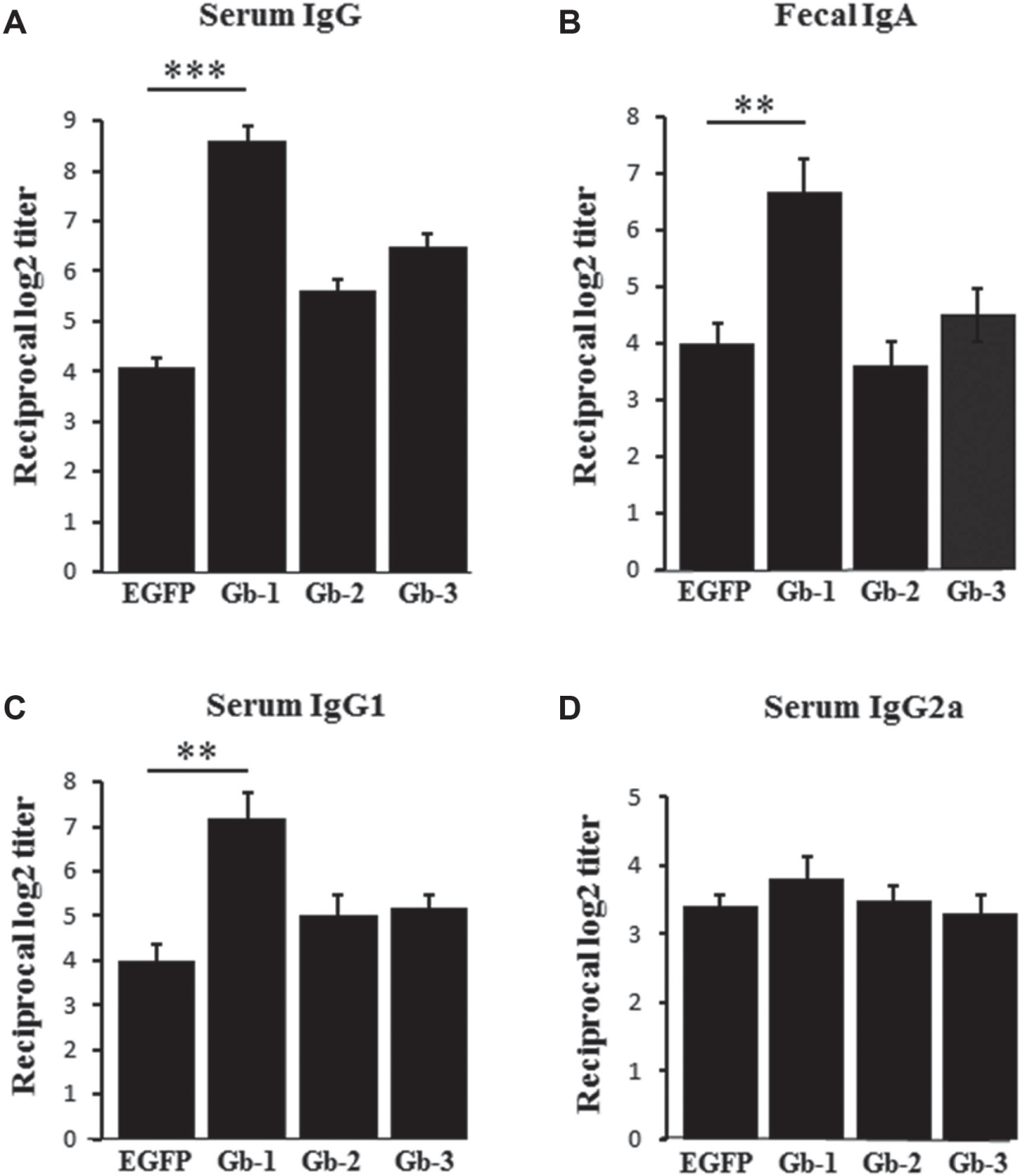
Enhanced green fluorescent protein (EGFP)–specific mucosal and systemic immune responses in orally immunized mice. Level of EGFP-specific immune response in mice orally immunized with the indicated antigens was evaluated 5 days after the last immunization by enzyme-linked immunosorbent assay (ELISA). (
At the mucosal level, mice immunized with Gb1-fused EGFP induced EGFP-specific fecal IgA level >1.5-fold stronger than those immunized with other ligand-fused EGFPs or with EGFP alone ( Fig. 2B ). IgA class switching is the process whereby B cells acquire the expression of IgA. IL-4, IL-5, and IL-6, and are known to induce IgA class switching. They not only enhanced the induction of Gb-1–specific fecal IgA but also increased the number of Gb-1–specific IgA-secreting cells in PPs compared with EGFP alone ( Fig. 3B ). In addition, oral administration of Gb1-fused EGFP also enhanced the priming of EGFP-specific lymphocytes ( Fig. 3C ).
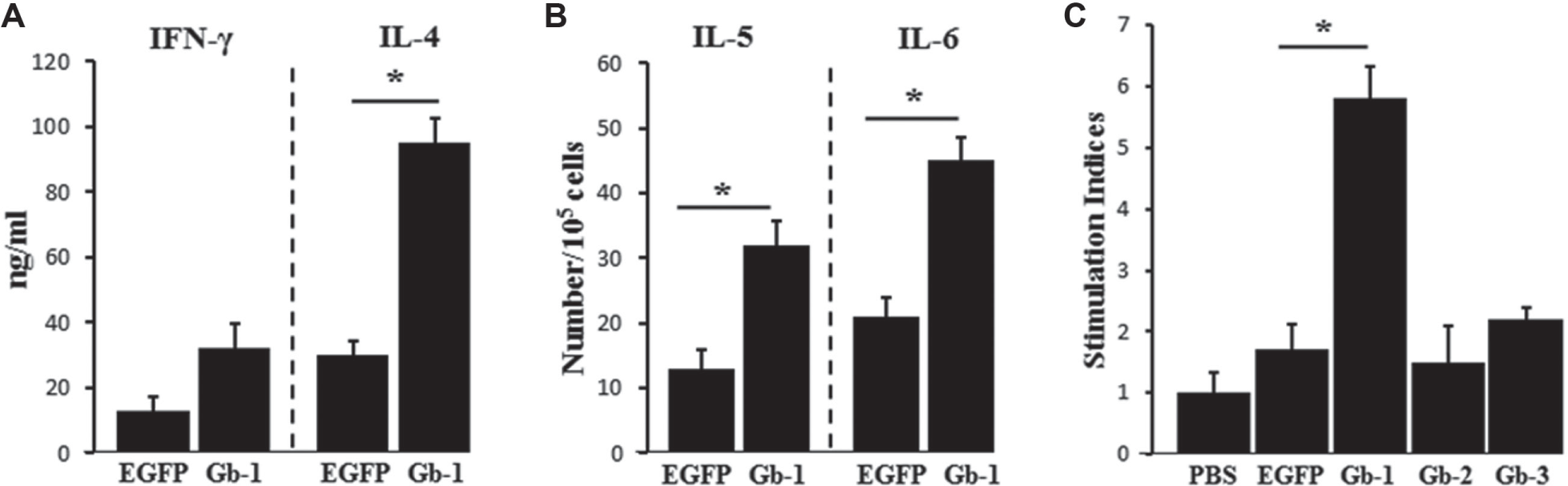
Levels of cytokines and cytokine-secreting cells and splenocyte proliferation in Gb-1–immunized mice. Levels of cytokines and cytokine-secreting cells were determined using enzyme-linked immunosorbent assay (ELISA) and enzyme-linked immunospot (ELISPOT), respectively, as described in the Materials and Methods, after in vitro stimulation of cells from spleen and Peyer’s patches (PPs). (
Interferon-γ (IFN-γ) is the representative cytokine for Th1-type response, and IL-4 and IL-6 represent the Th2-type response.19,20 We analyzed the pattern of cytokine secretion in splenocytes restimulated with EGFPs. IL-4 level was significantly raised in the Gb-1–immunized group compared to the EGFP group ( Fig. 3A ). Since IgG1 production and IL-4 secretion are indicators of Th2-type response, this implies that oral immunization with Gb-1 ligand induces predominantly Th2-type immunity against the subject antigen.
Discussion
Oral vaccination induces not only antibody response in the systemic compartment, but also the serum IgG response is strengthened by IgA production at the mucosal surface.19,21 This makes oral vaccination superior compared to systemic vaccination where only the IgG response is elicited. 22 Low cost and needle-free delivery are other attractive features of mucosal/oral vaccination, 4 so it is a feasible and economic vaccination strategy, especially in developing countries. However, low permeability of vaccines across the plasma membrane of intestinal epithelial cells is a major limitation to be overcome for the successful development of an oral vaccine. In the case of subunit vaccines, denaturation caused by the acidic environment of the stomach is also a barrier to oral vaccines. 23 Encapsulation of vaccines in nano- and microparticulate forms such as polylactide (PLA), polylactide-coglycolide (PLGA), or liposomes, and the use of enteric-coated capsules may offer some protection against acidic and enzymatic degradation24–29; however, these strategies have affected the delivery of vaccines across intestinal epithelial cells.
One approach to enhance drug and particulate delivery systems across the intestinal epithelial barrier is to target Ags to specific transcytotic receptor of the intestine. 23 M cells are specialized cells that internalize luminal Ags across the epithelial layer to the underlying immune induction sites without degradation and play an important role in initiating Ag-specific mucosal immune responses by inducing the production of secretory IgA.2,30 Therefore, the M-cell–mediated Ag sampling is believed to be an essential and critical step for eliciting a successful mucosal immune response.
In this regard, identifying mucosal vaccine adjuvants that exploit M cells would be an effective strategy to develop successful mucosal vaccines. 31 Research has focused on the characterization of M-cell–specific markers and their exploitation to enhance M-cell targeting efficiency of the antigen in oral vaccination. The identification of M-cell–specific markers made M cells possible to be targeted with delivery vehicles. GP-2 was shown to express specifically on M cells and acts as mucosal transcytosis receptor. 10
We used a phage display library to screen peptide ligands against GP-2 and validated their binding to GP-2 in vitro and in vivo by ELISA and immunohistochemistry (IHC). Selected peptides showed high binding affinity to recombinant GP-2 in vitro, especially peptide Gb-1. Competition ELISA results showed that the selected peptides do not bind to a common or overlapping epitope on GP-2. Immunohistochemical staining of whole-mount and frozen sections of PPs confirmed the binding of Gb-1 to M cells ( Fig. 1A , B ) and hence promoted the internalization of EGFP into PPs significantly compared to EGFP alone. However, we could not demonstrate this for the other two peptides profiled, suggesting that other factors also function. The selected peptides most likely interact with the target receptor in a multivalent fashion. Multivalent presentation of a ligand is a well-accepted approach to increase the biological potency of a ligand through enhanced ligand-receptor interaction and, in some cases, cellular internalization by surface receptor oligomerization. Peptide multimerization can also improve peptide stability and extend its half-life. 32 The conformation and/or dimerization of the peptide when presented as a conjugate to a protein may differ from the conformation or presentation of a free peptide in solution. Thus, conformation constraint or polymerization may be important for full retention of receptor binding activity. This is illustrated, in part, by peptides Gb-2 and Gb-3, which showed no binding to M-cell membranes when conjugated to EGFP peptide but demonstrated binding to GP-2 when presented as free peptides in solution.
Next, we evaluated the ability of selected ligands to stimulate the immune system. We immunized mice orally with a ligand-fused Ag and monitored immune induction in systemic and mucosal compartments. Gb-1 significantly (p < 0.001) enhanced antigen-specific serum IgG, particularly IgG1 in the systemic compartment as well as mucosal IgA, and the concentration of IgA-switching cytokines (IL-5 and IL-6) compared to EGFP alone or the other peptides. Similarly, elevated levels of IFN-γ and IL-4 were detected in supernatant from lymphocytes collected from the Gb-1–immunized group compared to control groups.
In summary, we used phage library biopanning to screen peptide ligands against mucosal transcytosis receptor GP-2 and validated an identified peptide’s role as a mucosal adjuvant. Results show that Gb-1 peptide has immune stimulatory ability and that it induces Th2 immune response. These findings suggest that peptide Gb-1 can be used as adjuvant for the development of mucosal vaccines.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Major National Projects for Infectious Diseases Research (2008ZX10002002, 2012ZX10002-006-002-003) and for New Drug Research & Development (2009ZX0913-710, 2011AA02A114), the National Natural Science Foundation of China (30571650, 31370927), and project of the Science and Technology Commission of Shanghai (13431900602).