
Abstract
Receptor tyrosine kinase–like orphan receptor (ROR) proteins are a conserved family of tyrosine kinase receptors that function in developmental processes including cell survival, differentiation, cell migration, cell communication, cell polarity, proliferation, metabolism, and angiogenesis. ROR1 has recently been shown to be expressed in various types of cancer cells but not normal cells. Pharmacokinetics and pharmacodynamics of single-chain Fragment variable (scFv) antibodies provide potential therapeutic advantages over whole antibody molecules. In the present study, scFvs against a specific peptide from the extracellular domain of ROR1 were selected using phage display technology. The selected scFvs were further characterized using polyclonal and monoclonal phage enzyme-linked immunosorbent assay (ELISA), soluble monoclonal ELISA, colony PCR, and sequencing. Antiproliferative and apoptotic effects of selected scFv antibodies were also evaluated in lymphoma and myeloma cancer cell lines using MTT and annexin V/PI assays. The results of ELISA indicated specific reactions of the isolated scFvs against the ROR1 peptide. Colony PCR confirmed the presence of full-length VH and Vκ inserts. The percentages of cell growth after 24 h of treatment of cells with individual scFv revealed that the scFv significantly inhibited the growth of the RPMI8226 and chronic lymphocytic leukemia (CLL) cells in comparison with the untreated cells (p < 0.05). Interestingly, 24-h treatment with specific scFv induced apoptosis cell death in the RPMI8226 and CLL cells. Taken together, our results demonstrate that targeting of ROR1 using peptide-specific scFv can be an effective immunotherapy strategy in hematological malignancies.
Introduction
Cancer-targeted therapy has emerged as a new promising treatment option for various cancers. 1 As part of the large effort in target discovery, receptor tyrosine kinase–like orphan receptor (ROR) proteins have recently been found to be expressed in various types of human cancers.1,2 ROR1 is a transmembrane receptor tyrosin kinase (RTK) of importance during the normal embryogenesis of the central nervous system, heart, lung, and skeletal systems but is not expressed in normal adult tissues. 3 However, ROR1 is found to be overexpressed in several cancer cells with a different origin as survival factor. 1 The ROR1 extracellular domain is composed of three conserved parts including an immunoglobulin-like motif, a cysteine-rich frizzled domain, a kringle domain followed by a transmembrane, and an intracellular tyrosine kinase domain. 4 It was named orphan receptors because its endogenous ligand had not yet been discovered. 5 Some reports suggested that ROR1 is a pseudokinase that is crucial for Met-driven tumorigenesis.6,7
Aberrant expression of ROR1 in tumor cells, although totally absent in healthy peripheral blood cells and almost other normal adult tissues, may provide a potential target for novel therapeutics including monoclonal antibodies (mAbs) and small-molecule inhibitors of tyrosine kinases.1,8 Promising experimental and preclinical findings have been reported in various cancer types including B-cell chronic lymphocytic leukemia, B-cell acute lymphocytic leukemia, mantle cell leukemia, pancreatic carcinoma, and lung and breast cancer. Moreover, recent findings revealed that ROR1 might be a potential oncofetal antigen for active immunotherapy.1,9
In the mid-1980s, chimerization and humanization provided state-of-the art techniques to avoid the side effect of human anti-mouse immune reaction (HAMA). However, HAMA reaction was still observed to a varying extent, and the cost- and time-consuming processes have also made these techniques more complicated. 10 Today, single-chain fragment variable (scFv) has been developed as a robust technology offering great potential for selecting specific, high-affinity fully human antibodies against various self- and non–self-target antigens. 11 A growing number of antibodies in clinical trials are derived from phage-display technology.12,13
In the present study, we employed phage display technology to isolate scFv antibodies against specific oligopeptides from the extracellular domain of ROR1 using a naïve human phage library (Tomlinson I + J). The specificity of selected scFvs was confirmed using polyclonal and monoclonal enzyme-linked immunosorbent assay (ELISA). Growth inhibitory and apoptotic effects of selected scFv antibodies on Raji and RPMI8226 cell lines and leukemic cells from chronic lymphocytic leukemia (CLL) patients were evaluated using MTT and annexin V/PI assays, respectively.
Materials and Methods
Materials and Samples
The study was approved by the local ethics committee. All patient samples were obtained with informed consent in keeping with the Helsinki declaration. Peripheral blood mononuclear cells were isolated by Ficoll Hypaque density-gradient centrifugation from heparinized blood.
The human single-fold scFv libraries I + J (Tomlinson I + J), Escherichia coli TG1Tr, were obtained from the MRC Laboratory of Molecular Biology and the MRC Centre for Protein Engineering (Cambridge, UK) for the phage propagation, and E. coli HB2151 was employed for expression of soluble scFvs. M13K07 helper phage was purchased from New England Biolabs Inc. (Ipswich, MA). Anti-c-Myc and anti-M13 horseradish peroxidase (HRP)–conjugated monoclonal antibodies were purchased from Roche (Berlin, Germany) and Amersham International Plc (Little Chalfont, UK), respectively. Peptide (H-VATNGKEVVSSTGVLFVKFGP-OH) was obtained from JPT (Berlin, Germany). Recombinant human ROR1 was obtained from Roche Molecular Biochemicals (Basel, Switzerland). The annexin V/PI apoptosis assay kit was purchased from BD Pharmingen (CA, USA).
Preparation of Phage Libraries for Biopanning
Briefly, 500 µL of the library stock (Tomlinson I + J) was incubated in 200 mL prewarmed 2xYT (16 g tryptone, 5 g NaCl, and 10 g yeast extract in 1 L) containing 1 % glucose and 120 µg/mL of ampicillin at 37 °C with shaking (250 rpm) until optical density (OD) 600 reached 0.4 to 0.5 (1–2 h). Then, about 5 × 1011 plaque-forming units of M13K07 helper phage (New England Biolabs Inc.) was added to each milliliter of cell suspension and incubated without shaking for 30 min at 37 °C and for another 30 min in a shaker. Subsequently, infected bacteria were collected (centrifuged at 4000 rpm at 4 °C for 10 min), resuspended in 100 mL 2xYT containing 50 µg/mL kanamycin (Sigma, St. Louis, MO), 120 µg/mL ampicillin, and 1% glucose and incubated overnight at 30 °C with shaking (200 rpm) and centrifuged at 4000 rpm at 4 °C for 20 min. Phages were precipitated from supernatant with polyethylene glycol (PEG)/NaCl (20% PEG 6000, 2.5 M NaCl) and collected by centrifugation at 14,000 rpm at 4 °C for 20 min. Phages were stored at −70 °C in 15% glycerol or at 4 °C for longer-term and short-term storage, respectively.
Panning Process
After library reamplification and helper phage preparation, isolation of specific phage antibody clones was carried out through a solid phase selection process. Briefly, maxisorp immunotubes (Nunc, Roskilde, Denmark) were coated with 50 µg/mL of peptide in phosphate-buffered saline (PBS; for positive screening) and 50 µg/mL of bovine serum albumin (BSA; for negative screening) for 16 h at 4 °C. The coated immunotubes were washed three times with PBS and blocked with 5% MPBS (5% skim milk powder in PBS) at 37 °C for 90 min. Subsequently, immunotubes were washed three times with PBS, and 1012 phages in 4 mL of 5% MPBS (input 1) were added to the BSA-coated immunotube and then coated and incubated at 37 °C for 60 min on a rotator (negative panning). Afterward, supernatants containing phages not bound to BSA were added to the immunotube coated with specific peptide and incubated at 37 °C for 30 min with shaking (50 rpm) and a further 30 min without shaking (positive panning). The immunotubes were washed 10 times with PBST 0.5 % (PBS containing 0.5% Tween 20), five times with PBS, and two times with distilled water to remove the unbound phages, and the binders were eluted from the immunotube by adding 2 mL of triethylamine (100 mM, pH 10.0). Following 10 min at room temperature (RT), 2 mL of Tris-HCl (0.5 M, pH 7.4) was added to neutralize the eluted phages solution (output 1). To propagate eluted phages, Escherichia coli TG1 cells (OD 600 = 0.5) were infected with eluted phages and M13K07 helper phage. Subsequently, propagated phages were collected from supernatant as described above and named input 2. The titer of outputs and inputs was also determined. Five rounds of panning were performed for isolation of ROR- specific scFv-phage clones.
Assessment of Panning Phage Outputs by Polyclonal Phage ELISA
The reactivity of phages obtained from each round of panning to ROR1 peptide was assessed by polyclonal phage ELISA. Briefly, ELISA plates (Maxisorp, Nunc) were coated with 50 µg/mL of ROR1 peptide (100 µL per well) and incubated overnight at 4 °C. The wells were washed two times with PBS and blocked with 5% MPBS (250 µL per well) for 90 min at 37 °C. Precipitated phages from each round of panning were diluted in 5% MPBS and added to the wells and incubated at 37 °C for 30 min with shake (50 rpm) and a further 30 min without shaking. Following a washing process, anti-M13 HRP-conjugated monoclonal antibodies (1:5000 dilution in 0.1% BSA-PBS) were added, and the wells were washed five times with PBS-0.05% Tween 20 and two times with PBS. TMB substrate solution was added, and the reaction was stopped by adding 1 M sulfuric acid (50 µL per well). ODs were measured at 450 nm by an ELISA reader (BP-800, Biohit, Helsinki, Finland).
Assessment of Isolated Clones by Monoclonal Phage ELISA
Reactivity of the single clone to ROR1 peptide was assessed by monoclonal phage ELISA. Briefly, E. coli TG1 cells (OD 600 = 0.5) were infected with phage outputs from the fourth and fifth rounds. Infected TG1 cells were spread on TYE agar plates (8 g NaCl, 5 g yeast extract, 10 g tryptone, and 15 g Bacto-Agar in 1 L) containing 1% glucose and 120 µg/mL ampicillin and incubated overnight at 37 °C. Fifty-five individual colonies were randomly picked from the plates and inoculated into 2xYT (containing 1% glucose and 120 µg/mL ampicillin) and incubated overnight at 37 °C with shaking (250 rpm). Five microliters of each culture from wells was used to infect a second plate containing 200 µL 2xYT with 1% glucose and 120 µg/mL ampicillin and incubated at 37 °C with shaking until OD 600 reached 0.4 to 0.5. M13K07 helper phage was added to each culture and incubated 30 min without shaking and 30 min with shaking; spun down at 2200 rpm at 4 °C; resuspended in 2xYT containing 50 µg/mL kanamycin, 120 µg/mL ampicillin, and 1% glucose; and incubated overnight at 30 °C with shaking. Subsequently, phages were precipitated from the supernatant and used in phage ELISA as described above.
Production of Soluble scFvs
To produce soluble scFvs, positive phage clones selected from monoclonal phage ELISA were employed to infect exponentially growing E. coli HB2151 (a nonsuppressor strain) and incubated at 37 °C with shaking (250 rpm) until OD 600 reached 0.9. The scFv gene expression was induced using isopropyl β-D-thiogalactoside (IPTG) at a final concentration of 1 mM for 18 h (250 rpm) at 30 °C with shaking. The cells were centrifuged at 3600 rpm for 10 min, and supernatant containing antibody fragments was collected and used in ELISA as described above, with the exception of anti-c-Myc-HRP to detect soluble scFv fragments.
Specificity Assay
Specificity of the soluble scFvs was assessed using three different antigens in ELISA. Briefly, ELISA plates were coated with 50 µg/mL of various antigens including insulin-like growth factor–I receptor (IGF-IR) and frizzled-receptor 7 (Fzd7). All samples were run in triplicate, and each assay was repeated three times in different days.
Colony PCR and DNA Sequencing
Reactive phage clones from the soluble scFv ELISA were amplified by PCR to investigate the presence of full-length VH and Vκ gene inserts (930 bp). PCR was performed using pHEN seq (CTA TGC GGC CCC ATT CA) and LMB3 (CAG GAA ACA GCT ATG AC) phagemid-specific primers with the following PCR program: initial denaturation at 95 °C for 5 min, then denaturing at 94 °C for 40 s, annealing at 55 °C for 40 s, extension at 72 °C for 40 s, for 40 cycles, and final extension at 72 °C for 5 min. The PCR products were examined by electrophoresis in a 1% agarose gel. PCR products were cut from gel, purified with the QIAquick gel extraction kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol, and sequenced by Bioneer Company (Daejeon, South Korea), and obtained sequences were then blasted in the IMGT database.
Electrophoresis and Western Blot Analysis
Western blot analysis was carried out to confirm the production of the soluble scFv protein. Expression of scFv antibody fragments was induced by adding 25 µL/well of 2xYT-A containing 9 mM IPTG and 0.9 M sucrose (at final concentrations of 1 mM and 100 mM, respectively), and the cultures were analyzed for protein expression through Western blotting. The bacterial periplasmic was subjected to electrophoresis in 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) as described previously. Subsequently, the gel was stained with Coomassie Blue or transferred on polyvinylidene fluoride membrane (Millipore, Billerica, MA) using a semidry transfer device (Amersham Biosciences, Freiburg, Germany). The membrane was blocked with the blocking solution and incubated with 1:2000 dilution of mouse anti-myc tag antibody (Santa Cruz Biotechnology, Santa Cruz, CA). The membrane was then incubated with HRP-conjugated anti-mouse IgG (1:5000 dilution; Santa Cruz Biotechnology), and the protein bands were visualized by enhanced chemiluminescence (Amersham Biosciences).
MTT Cytotoxicity Assay
Raji Burkitt’s lymphoma cells and RPMI8226 multiple myeloma cell lines were purchased from the National Cell Bank of Iran. Primary cancer cells from a newly diagnosed CLL patient were also included. The cells were cultured in RPMI-1640 medium (Gibco, New York, NY), supplemented with fetal bovine serum (10%), streptomycin (100 µg/mL), and penicillin (100 U/mL) at 37 °C in a humidified 5% CO2 incubator and seeded into a 96-well flat-bottomed plate (Nunc) at a density of 15 × 103/well for 24 h. Subsequently, the medium was replaced with fresh medium and treated with various concentrations of selected scFv (500, 1000, and 2000 scFvs/cell) and anti-BSA scFv (as negative controls) for 24 h, and the cell growth was determined using MTT assay. Briefly, the old media were replaced with fresh media containing 0.5 mg/mL of MTT (Sigma, Germany) solution, followed by a 4-h incubation at 37 °C. The supernatants were gently removed, and 150 µL DMSO (Merck, Darmstadt, Germany) was added to each well and incubated in a shaker at RT for 20 min. The colorimetric evaluation was performed using an ELISA reader (BP-800, Biohit) at 570 nm.
Apoptosis (Annexin-V) Assay
Apoptosis cell death was assessed using the annexin V-FITC apoptosis detection kit (BD Pharmingen, San Jose, CA) according to the manufacturer’s protocol. Briefly, CLL, Raji, and RPMI8226 cells were seeded into a culture plate well at 37 °C overnight. Medium was replaced with fresh medium containing selected scFv (2000 scFvs/cell) or anti-BSA scFv at 37 °C for 24 h. Cells were harvested and washed twice with cold PBS and stained with annexin V-FITC and PI in 1× binding buffer for 15 min at RT. Finally, the cells were analyzed by FACSCalibur (Becton Dickinson, Mountain View, CA). Each assay was repeated three times.
Statistical Analysis
All data were analyzed in SPSS-16 software using a one-way analysis of variance (ANOVA) test. A p value <0.05 was considered statistically significant.
Results
Isolation of ROR1-Specific scFvs
Human single-chain scFv libraries I + J (Tomlinson I + J) were amplified and used for isolation of scFvs against ROR1-specific peptide. The phage titers of five outputs and inputs were determined by colony-forming units (cfu) counting. The phage titer of input 1 was approximately 1012 cfu/mL. The results showed that the number of eluted phages (titer of outputs) was increased from 103 to 107 cfu/mL following five rounds of panning ( Fig. 1 ), indicating phage enrichment of output through the panning process.
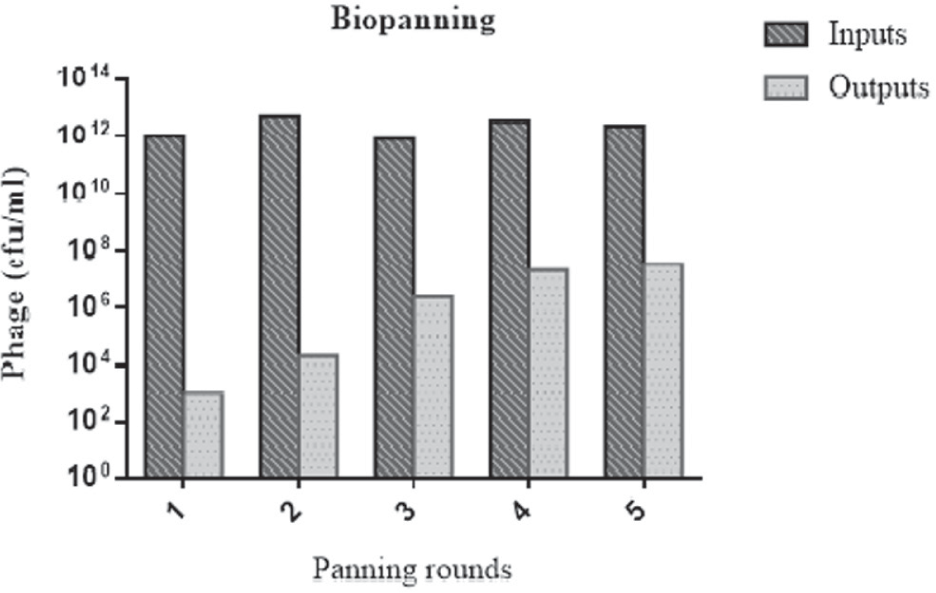
The titer of outputs and inputs from five rounds of panning. cfu = colony-forming unit.
Polyclonal Phage ELISA
As shown in Figure 2A , the OD values of wells coated with ROR1 peptide were higher than uncoated wells and increased round by round from 0.52 to 0.89, indicating the effectiveness of the panning process.
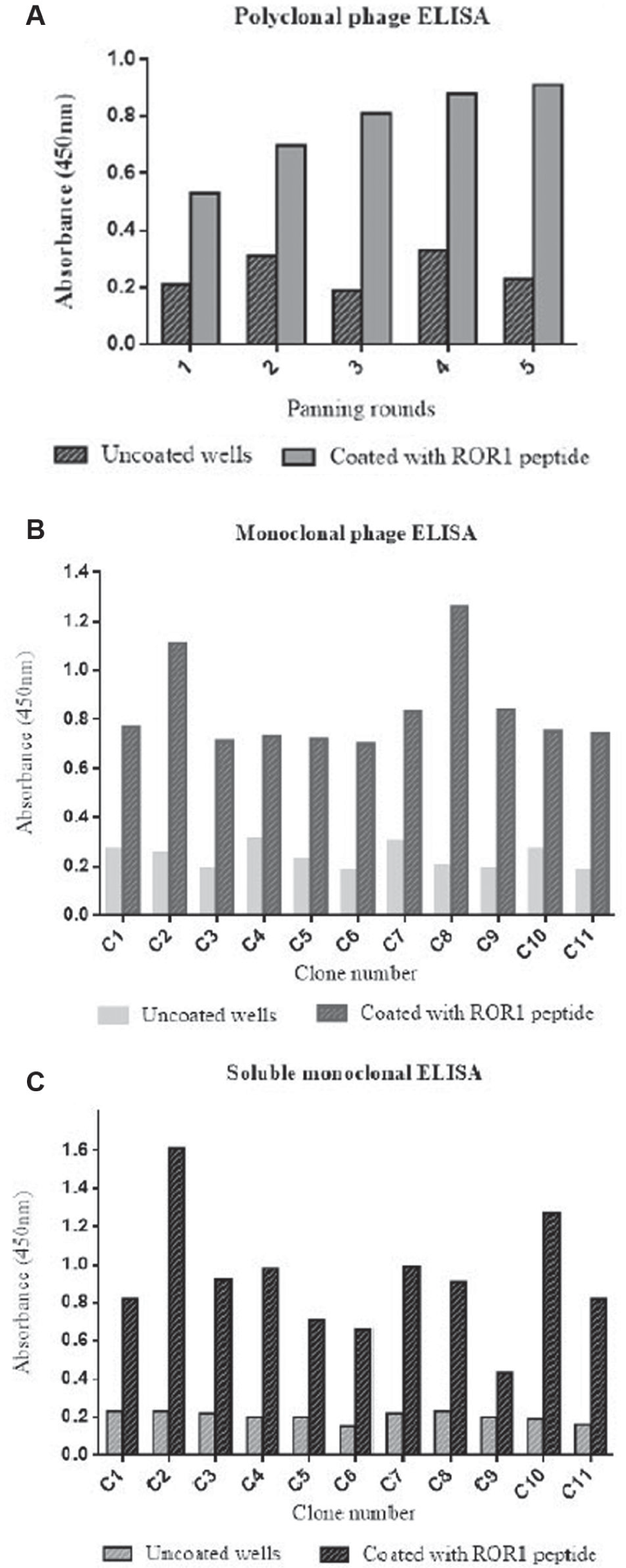
(
Assessment of Phage and Soluble scFv by Monoclonal Phage ELISA
Sixty-three individual clones from the fourth and fifth rounds of panning were randomly selected and screened by monoclonal phage ELISA; of these, 11 clones represented positive reactivity to ROR1 peptide ( Fig. 2B ).
Eleven positive clones from the monoclonal phage ELISA were employed to infect HB2151 (a nonsuppressor strain) cells, and expression of soluble scFv was induced by IPTG. Production of soluble scFv was evaluated using ELISA ( Fig. 2C ). Ten of 11 soluble scFvs represented positive reactivity as absorbance of ROR1 peptide/absorbance of uncoated wells >3.
Specificity Assay
The specificity of soluble monoclonal scFvs to irrelevant peptide IGF-IR, Fzd7, skimmed milk, and BSA was assessed by ELISA. As shown in Table 1 , no evident reactivity to irrelevant peptides was observed.
Specificity of soluble monoclonal scFvs by ELISA assay to a variety of antigens. a
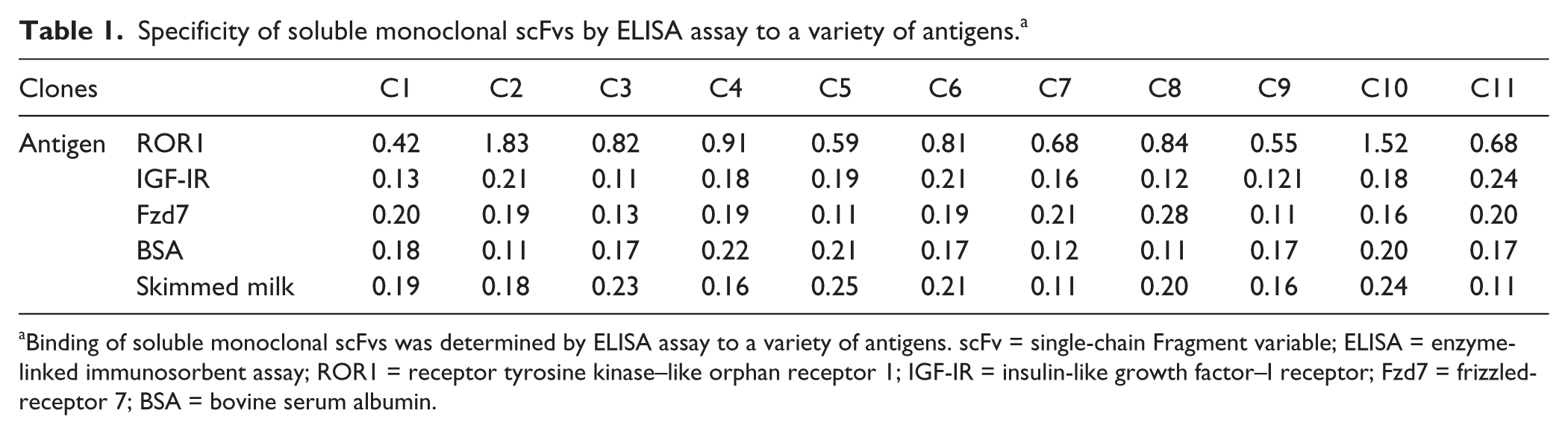
Binding of soluble monoclonal scFvs was determined by ELISA assay to a variety of antigens. scFv = single-chain Fragment variable; ELISA = enzyme-linked immunosorbent assay; ROR1 = receptor tyrosine kinase–like orphan receptor 1; IGF-IR = insulin-like growth factor–I receptor; Fzd7 = frizzled-receptor 7; BSA = bovine serum albumin.
Colony PCR and DNA Sequencing
Ten reactive clones from the soluble monoclonal ELISA were subjected to PCR amplification to investigate the presence of full-length VH and Vκ inserts (930 bp). A single band corresponding with the scFv gene was detected in 5 of 10 individual clones ( Fig. 3A ). All PCR products were extracted from gel and sequenced. Three different sequences were found, and alignment in the IMGT database revealed that the scFv nucleotide sequences derived from human VH and Vκ family.
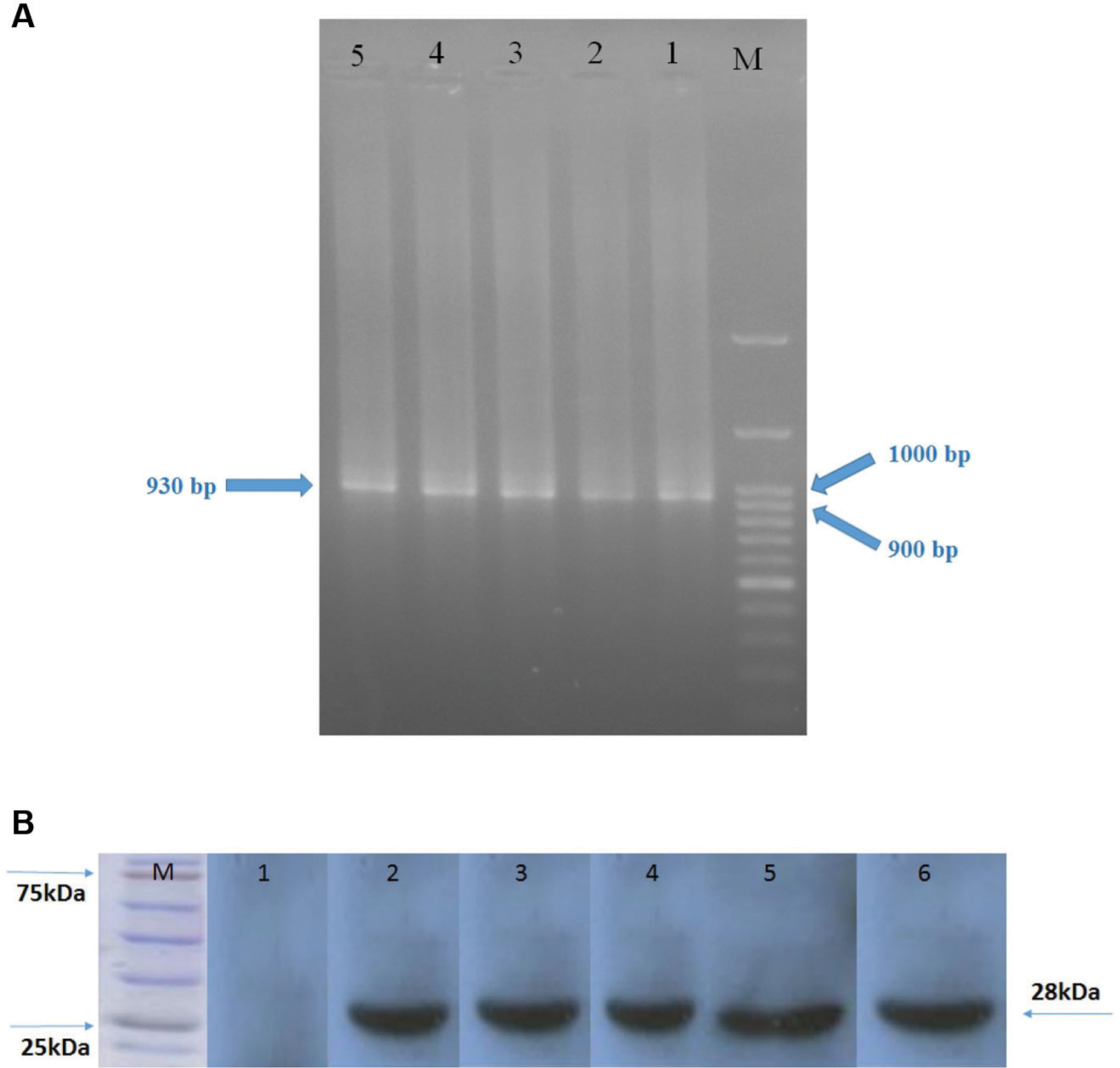
Electrophoresis of PCR products of the selected single-chain Fragment variable (scFv) genes. Lane M: DNA marker. Lanes 1–5: PCR products of the selected clones (
Electrophoresis and Western Blot Analysis
The crude periplasmic extract of HB2151 was resolved by SDS-PAGE in a reduced condition, and the protein bands were visualized by staining and Western blot analysis. The Western blot results confirmed the presence of scFv protein with a molecular weight of ~28 kDa ( Fig. 3B ).
Antiproliferative Effects of Anti-ROR1 scFv Antibody
The most reactive clone was further evaluated for growth-inhibitory properties. No significant antiproliferative effect was observed at concentrations of 500 and 1000 scFv/cell. However, treatment with 2000 scFvs/cell showed significant growth inhibition of RPMI8226 and CLL cells after 24 h (p < 0.05). In addition, no significant growth inhibition was observed in Raji Burkitt’s lymphoma cells treated by anti-ROR1 scFv (p > 0.05). The growth inhibition of Raji, RPMI8226, and CLL cells treated was 2000 scFvs/cell were 9.12%, 32.86%, and 38.94%, respectively ( Fig. 4 ). No significant growth inhibition was observed in cells treated with anti-BSA scFv (as negative control; p > 0.05).
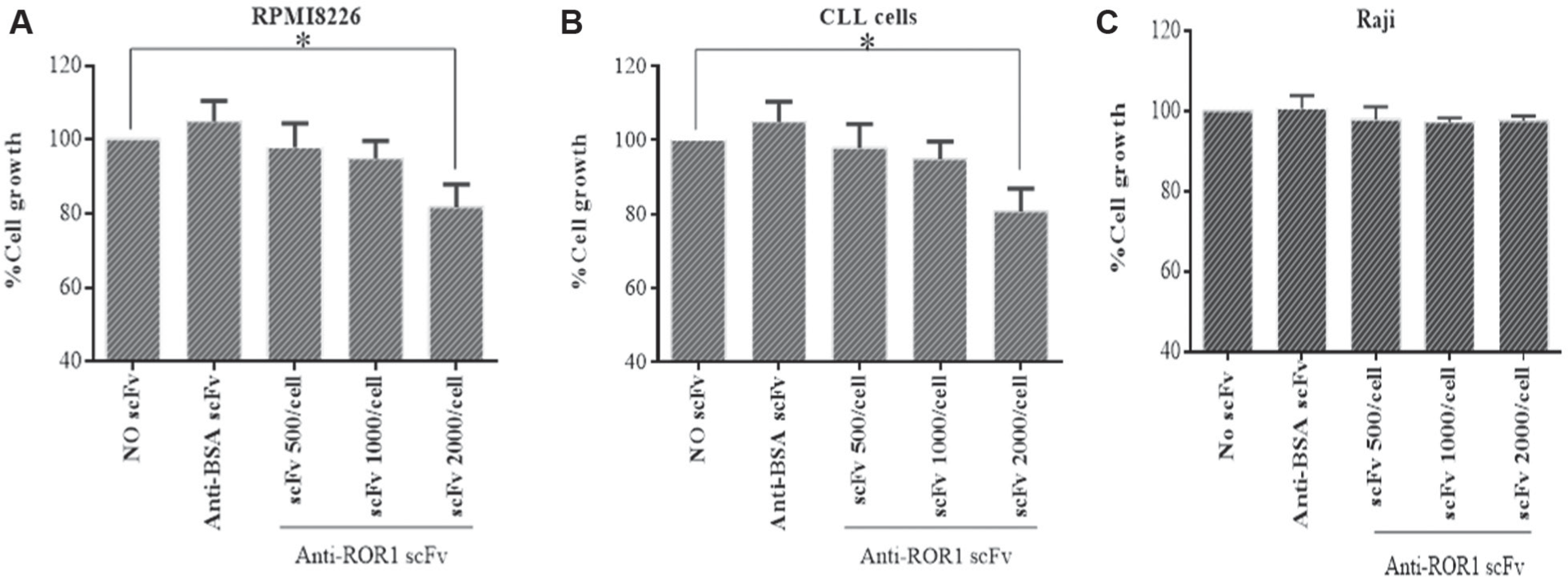
Antiproliferative effects of different concentrations of anti-ROR1 single-chain Fragment variables (scFvs) on RPMI8226, chronic lymphocytic leukemia, and Raji cells (
Apoptotic Effects of Anti-ROR1 scFv Antibody
Following 24 h, the majority of untreated cells (81.27% of RPMI8226, 78.48% of CLL cells, and 80.03% of Raji cells) were alive (annexin V−/PI−), and 37.10% of RPMI8226 cells, 36.41% of CLL cells, and 14.43% of Raji cells were at early apoptotic (annexin V+/PI−) and late apoptotic phase (annexin V+/PI+; Fig 5A–C ). However, scFv treatment (2000 scFvs/cell) resulted in apoptosis cell death in 48.13% of RPMI8226 cells, 50.07% of CLL cells, and 26.28% of Raji cells. In the cells treated with anti-BSA scFv (as negative control), apoptosis cell death was observed in 26.40% of RPMI8226 cells, 27.22% of CLL cells, and 23.67% of Raji cells ( Fig. 5D–F ).
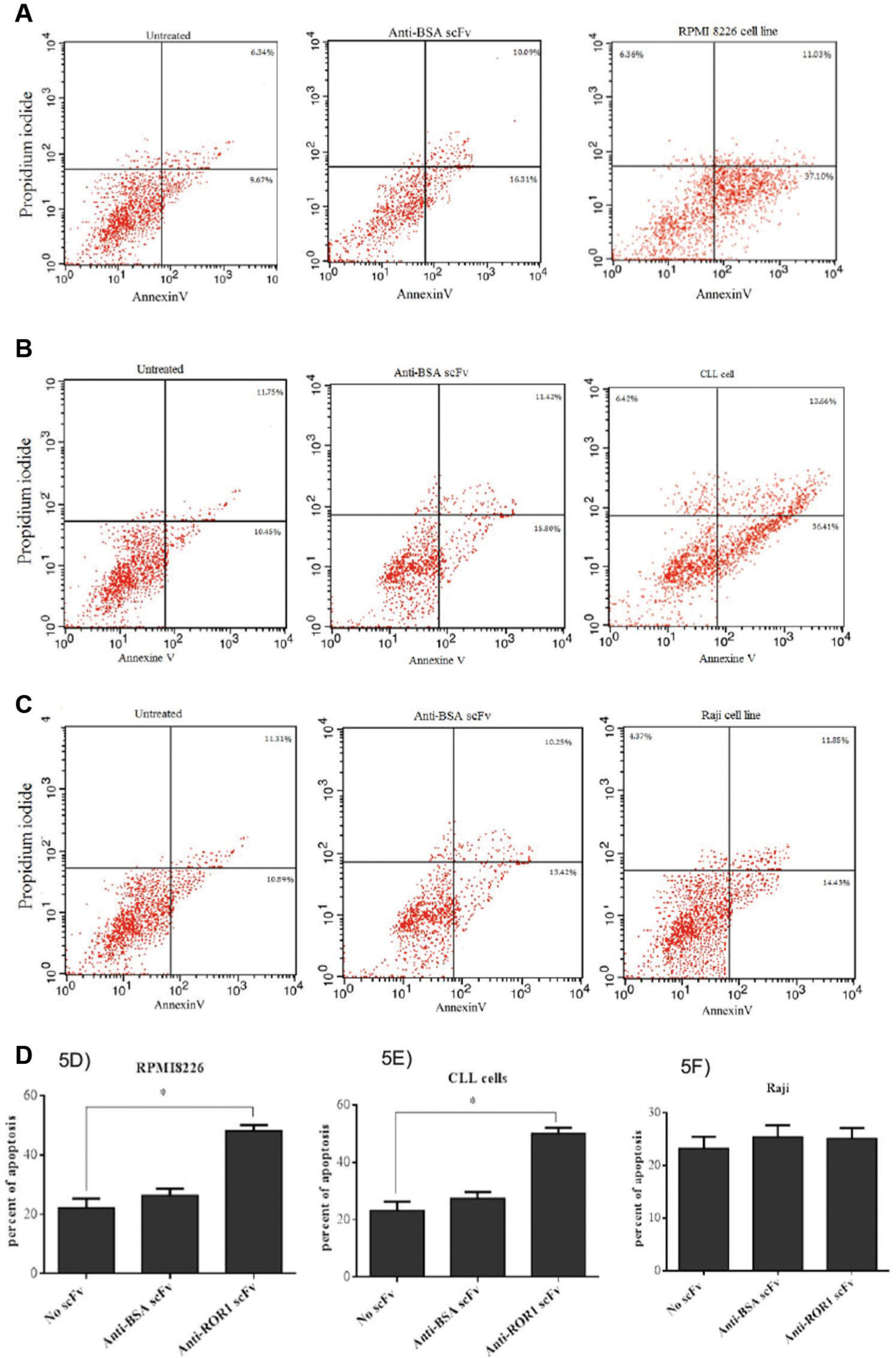
Representative dot plot analysis of apoptosis in RPMI8226, chronic lymphocytic leukemia (CLL), and Raji cells treated with anti-ROR1 single-chain Fragment variable (scFv;
Discussion
Receptor tyrosine kinases play a critical role in normal cell development and in tumorgenesis. 14 ROR1 has been shown to be overexpressed in various cancer types but not in normal tissues. 15 Aggressive tumors expressed higher levels of activated ROR1, and the silencing of the ROR1 gene resulted in specific tumor growth inhibition and apoptosis cell death.16,17
Owing to the impacts of immunotherapy and mAbs in targeted therapy of various diseases, a large number of mAbs have been produced against various biomarkers.18,19 Based on its aberrant expression profile, ROR1 is preclinically investigated as a new targeted therapy of hematopoietic and solid cancers.14,15,20
Phage display technology is a valuable immunotherapic tool that offers rapid, 21 efficient, and inexpensive methods for investigating protein-protein interactions, epitope mapping, functional and accessible sites from antigens, and antibody selection; however, pharmacokinetics is the main disadvantage of scFVs.22–24 Because of different advantages of scFvs over whole antibodies, scFvs have been offered as appropriate tools for targeting cancer biomarkers.23,24
In the present study we select a novel peptide from the Ig-like region of the extracellular domain of ROR1. The identity of the peptide was checked and approved using BLAST analysis. Any homolog sequence with this sequence was not found in other human proteins. The antigenicity of the peptide was further evaluated using online software for B-cell linear epitope prediction including Epic, Lbtope, and ABCpred. In silico prediction also confirmed a high-score immunogenicity thereof in the extracellular domain of ROR1. No potential site for N- or O-linked glycosylations was found in this peptide.
In the present study, successful isolation of the scFv antibodies against ROR1 peptide is presented. ScFv antibody fragments against specific oligopeptide from the extracellular domain of ROR1 were isolated using the human-naïve scFv library. Antiproliferative and apoptotic effects of selected scFv on the primary CLL, Raji, and RPMI8226 cells were investigated. The naïve phage library was panned against ROR1 peptide. The titer of output phages was gradually increased round by round until the fourth round but not in the fifth round. The results of polyclonal phage ELISA were consistent with the phages output titers, indicating successful enrichment of peptide-specific phages during the panning process. Monoclonal phage- and soluble-scFv ELISA results showed that the individual clones specifically reacted with the corresponding peptide compared with the control wells (uncoated wells). The OD values were three to five times greater than the ODs of control wells. In addition, no cross-reactivity was observed with a panel of irrelevant antigens. Colony PCR was performed and analyses of PCR product confirmed the presence of full-length VH and Vκ inserts.
The results of the cell proliferation assay showed that treatment with ROR1 scFv, but not BSA scFv, significantly inhibited the growth of CLL and RPMI8226 cells in a dose-dependent manner, indicating that the tumor growth inhibition was accomplished through specific binding of anti-ROR1 scFv to ROR1. In addition, these results could be related to receptor density because ROR1 has a different expression profile. This differential expression pattern—high expression in CLL and RPMI8226 cells and low expression in Raji cells—has led to an antiproliferative effect.
The result of the annexin V/PI assays was consistent with the MTT results, implying apoptosis cell death as a mechanism underlying the tumor growth inhibition property of the selected scFv. Although the minority of untreated and anti-BSA scFv-treated cells were apoptotic, an evident and remarkable apoptosis was observed in cells treated with anti-ROR1 scFv.
In a similar study by Daneshmanesh et al., 25 a panel of mouse mAbs was produced with the aim to explore ROR1 epitopes for the generation of human ROR1 mAbs for preclinical study. 25 All selected mAbs induced significant apoptosis in CLL cells as compared with control antibodies.25–27 In an in vivo study by Widhopf et al., 28 the effects of anti-ROR mAbs in TCL1 transgenic mice expressing ROR1+/CD5+/B220 low leukemic B cells were analyzed. Two anti-ROR1 antibodies, D10 and 4A5, against different epitopes were produced. D10, but not 4A5, anti-ROR1 mAb reduced phosphorylation of AKT. Leukemic cells were cleared from the blood and spleen of transgenic mice following intravenous injections of D10 mAb but not 4A5. 28
Cui et al. 29 reported that cirmtuzumab, a humanized anti-ROR1 mAb, was internalized and killed malignant cells. In addition, cirmtuzumab resulted in antibody drug conjugate–mediated clearance of ROR1-expressing CLL cells in xenografted mice as well as induced apoptosis cell death of breast and pancreas cell lines in vitro. 29
On the other hand, the inability of a panel of chimeric rabbit/human IgG1 to mediate effective Complement Dependent Cytotoxicity (CDC) and Antibody-Dependent Cell-Mediated Cytotoxicity (ADCC) is attributed as the relatively low cell surface density of ROR1. 30 Because of the low surface density of ROR1 on the cancer cell, development of antibody drug conjugates is speculated as an effective and specific approach in cancer-targeted therapy. 31
In comparison with previously reported mouse, chimeric, and humanized anti-ROR1 antibodies, this new scFv is isolated from a fully human phage display library and thus will not induce immune responses in the treated subject. Other advantages of this novel scFv over whole antibodies is its smaller size, high affinity to ROR1, low cost, and ease of production that make it suitable for targeting ROR1 in hematological malignancies.
In conclusion, the present study introduces a novel human anti-ROR1 scFv with antiproliferative and apoptotic effects and potential for ROR1 targeting therapy in hematological malignancies. However, further studies are demanded to evaluate in vivo tumor inhibitory effects of the selected scFv. A deeper understanding of the physiopathological role of ROR1 in normal and malignant cells is also required to optimally develop ROR1 targeted therapy for hematological as well as nonhematological tumors. Our future plan is to use animal models to confirm the in vivo targeting potential of the selected scFvs for targeting of ROR1. These preliminary findings should be further confirmed and the biological function of ROR1 in malignancies explored. In addition, further investigation needs to be done to establish whether selected scFvs have another in vitro and in vivo efficacy to act as antitumor agents.
Footnotes
Acknowledgements
The authors would like to thank the Immunology Research Center and Drug Applied Research Center, Tabriz University of Medical Sciences, for supporting the work.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.