
Abstract
Introduction
Psychiatric disorders have their genesis in early life. Standard screening approaches during well child visits for pathological infant distress, maternal depression, and dysfunctional parenting behaviors are likely inadequate. Microbiome measures and infant vocalizations have promise as scalable psychiatric biomarkers for infants. The purpose of this study was to examine associations among infant gut colonization based on microbiome measurements with maternal distress and maternal depressive symptoms in a sample of infants.
Methods
This study sought to examine infant microbiome correlates of infant distress, parent-infant interactions, maternal distress, and maternal depressive symptoms. We collected (N = 31) microbiome samples, infant vocalizations during vaccination, and behavioral measures during a 4 month well child visit (WCV) and did a battery of clinical assessments to assess for maternal depression, parent-child interactions, family characteristics and family stress. Whole-genome SHOTGUN sequencing was utilized to identify three types of associations: alpha-diversity using Shannon and Inverse-Simpson indexes, beta-diversity using Bray-Curtis and Jaccard distances, and differential abundance using LinDA. Spectral measures of infant cries were also modeled to assess potential relationships with clinical assessments and the microbiome.
Results
There were 19 phyla, 417 genera, and 1246 species identified with taxonomic classification. Maternal distress as measured by PHQ-9 scores obtained when infants were 2 months old were associated with 4 bacterial species (Actinomyces johnsonii, Bilophila wadsworthia, Clostridium dakarense and Ruminococcus flavefaciens; FDR < 0.1) and beta-diversity (p = 0.006-BC; p = 0.005-Jaccard). Infant cries with greater high frequency band power (p < 0.03) and a greater high-to-mid frequency ratio–metrics (p < 0.05) were associated with altered α-diversity of the microbiome. No correlations were present between maternal PHQ-9 at 4 months, PSI-IV and microbiome diversity.
Conclusion
The present findings suggest that an infant stress (assessed by quality of crying) is associated with lower microbiome diversity. Decreased diversity reflects an unhealthy microbiome. Parental depressive symptoms may also influence infant microbiome. Future interventional studies focused on the quality of the infant-caregiver relationship should examine related changes in intestinal microbiota.
Introduction
It is well established that infancy is considered the most important time for caregiver-infant interaction to affect the child's socio-emotional, neurobehavioral and cognitive outcomes. Soon after birth, mothers initiate species-typical behaviors including positive affect, close gazing, and affectionate touching. In the setting of a distressed infant, parents of newborns opt for physical soothing techniques and these techniques shift to the use of sounds and objects by 3 months.1–3 Maternal depression impairs parental attunement and the functionality of the dyad. Mothers with depression are more disengaged, show less positive affect, are more intrusive, and engage in less play during interactions with their infant as comparted to mothers who are not depressed.4,5 Current public health approaches for identifying maternal depression and attachment problems are ineffective.6–8
New mothers list a number of barriers to sharing their depressive symptoms during their obstetrics and gynecology or primary care appointments. These include shame, stigma, distrust in health care, worries about their newborn and how their depression might affect them as well as financial concerns. Additionally, it is difficult to assess the impact of maternal distress on infant's development and health. Studies have examined the use of potential biomarkers, such as the gut microbiome, as a correlating factor to maternal stress. 9 Rationale for the use of microbiome is the link between the infant gut microbiome and neurocognitive development.10,11 One study showed that higher abundance in Bacteroides species correlated with the highest cognitive performance and lower abundance correlated with lower communication, personal and social skills. On the other hand, there was no correlation between alpha diversity and neurodevelopment, but increased alpha diversity was associated to poorer cognitive performance.10,11 A study by Zijlmans et al tried to link maternal stress and its effect on infant's microbiota composition by following a cohort of 56 infants over 110 days after vaginal birth. 9 They measured maternal stress by having mothers complete a battery of self-reported questionnaires regarding anxiety and stress. In addition, they collected 5 saliva samples for cortisol concentration. Microbiome samples were collected from infants at 9 months of age and showed that infants’ microbiota composition was significantly affected by maternal stress that was either self-reported or measured by cortisol levels or both. Interestingly, the study found a weak correlation between the cortisol measure and self-reported questionnaires and the strongest association with the infant microbiota. Another study by Esposito et al 12 examined acoustical components of young infant's cry that gave different responses to depressed versus non-depressed mothers. The study found that depressed mothers interact with their babies less and are less responsive to their needs. It also showed that depressed mothers tend to respond to infants cries at higher fundamental frequencies and shorter durations. The study findings were limited in their generalizability due to the fact that it included mostly highly educated and low risk patients. Scalable, noninvasive, biomarkers, such as measurements of the gut microbiome and vocalizations, would have utility in augmenting standard screening procedures for maternal and family functioning during early well child visits.
This pilot study sought to examine scalable biomarkers during a routine well child visits that could predict the association between parental stress and style and infant stress on the infant microbiome. First, we examined the link between dynamic changes in infant gut colonization based on microbiome measurements at 4 months and the relationship style between infants and their caregivers based on behaviors observed during vaccination appointments. We also examined how infant cries during vaccination appointments correlate with their microbiome at 4 months of age. It was hypothesized that caregiver's soothing behaviors post immunization at 4 months appointment, maternal depressive symptoms, and parenting stress and style will have an impact on dynamic changes in infant gut colonization based on microbiome measurements at 4 months. We also hypothesized that infant cries measured during vaccination appointments and analyzed for spectral power will correlate with their microbiome at 4 months of age.
Methods
Participants
Prior to any research related activities, the study was approved by the XXX IRB (#XXX). All methods were performed in accordance with the relevant guidelines and regulations. Prior to participation, at least one parent completed written informed consent.
The research team recruited caregiver and infant dyads (N = 31) presenting for Well Child Visits (WCV) at 4-month appointments. Well child visits are primary care health maintenance visits for all children in the United States which occur at recommended intervals as recommended by the American Academy of Pediatrics including at 2 and 4 months of age.13 During the 4-month appointment, the infants received 5 different vaccines (pneumococcal conjugate vaccine (PCV13), rotavirus vaccine pentavalent (RV5), Diphtheria-Tetanus-acellular Pertussis and inactivated poliovirus with Haemophilus B influenzae conjugate vaccine (DTaP-IPV/Hib). Mothers were screened for post-partum depression at 2- and 4-months appointments. Pediatricians also collect infant height, weight, and general health information during the visit. Participants were recruited from the Pediatric and Family Medicine clinics at XXX, in XXX city, XXX state. Healthy infants born after 36 weeks of gestation were included. Exclusion criteria included: Infants born prematurely (36 weeks or younger), or infants with developmental delays, neurological impairments, or known congenital impairments (such as congenital heart disease) identified by diagnostic code during the first 4 months of life. Additional exclusions included caregivers not fluent in English as the consent process required this, children who were adopted or if their parents who did not give authorization to participate in the study, or families who did not consent to vaccination.
Data Collection
The following information was obtained via chart review on the infants: age, medical history, history of development delays or neurological issues, family history, social history, history of NICU stay, and medication use. The following information on mothers was obtained via chart review: age, medical history, psychiatric history, family history, social history, complications during pregnancy, labor and delivery history, history of postpartum psychiatric concerns, and the 9-item Patient Health Questionnaire (PHQ-9).
Microbiome Collection
To assist with obtaining the microbiome stool collection at the 4-month WCV, identified patients were contacted prior to the visit. At the 4-month appointment, caregivers were asked to bring a diaper of fecal material from the previous day or the day of their WBV. Once received, approximately 200 mg of fecal material from the diaper was placed in a tube (OMNIgene.GUT, OMR-200). These samples were stored at −80 °C until analysis.
Measures and Analyses
Behavior Patterns
Measure of Adult and Infant Soothing and Distress (MAISD) is a validated instrument that was used to determine adult and infant behaviors and the influence of the distress during immunization. 14 The MAISD measure was obtained at the 4-month appointment. Videotapes of caregiver and infant soothing behaviors were obtained during the immunization. Next, the videotapes were coded for 5 s epochs over 1 min period before the needle (Pre-Needle), and 30 s after the needle (30 s After-Needle) based on the presence (1) or absence (0) of each behavior. The following groups of behaviors were analyzed from the appointment: a) parent behavior including distract, offer object, offer pacifier, offer food, nurse (breastfeed), rub/massage/pat, kiss, hug, rock, verbally reassure, not visible on videotape, and b) infant behaviors including engaging in distraction, play with object alone, suck, consume food, smile/coo, cry, scream, flail, not visible (see Cohen et al 2005 for detailed description of the MAISD scale). 14 The three coders coded the data based on the review of the recording.
Parenting Stress
The Parenting Stress Index-IV-Short Form (PSI-4-SF) is a screening measure to evaluate parental stress and identify potential concerns that may lead to problems in the child's or parent's behavior. 15 PSI-4-SF includes 36 items which focuses on three major domains of stress: child characteristics, parent characteristics and situational/demographic life stress. The response for 33 of the 36 questions ranges from 1 (strongly disagree) to 5 (strongly agree). Three questions have different response options (see Lee et al for more details). 16 Scores were summed and calculated separately for the three subscales: defensive responding (DR), parental distress (PD) and parent-child dysfunctional interaction (P-CDI) (12 items on each domain, range from 12 to 60). A total stress score is the sum of 3 subscales with possible scores ranging from 36 to 180. Greater total stress scores indicate higher parental stress levels.
Depression Symptoms
Maternal depressive symptoms were assessed by the PHQ-9 collected at both the 2 and 4-month WCV appointments. 17 This scale is widely used to identify patients at risk of depression by 9-item questionnaire with each item scored on a 4-point scale ranging from 0 to 3. The PHQ-9 total score ranges from 0 to 27; higher scores represent more severe depressive symptoms. This scale has the following cutpoints for scores of 5 (mild), 10 (moderate), 15 (moderately severe) and 20 (severe depression).
Infant Cries
Audio during inoculation was recorded using Zoom H1n Digital Handy Recorder. Recordings were manually edited to isolate infant cries, excluding mother vocalizations, medical provider vocalizations, and other noises using Audacity software (Audacity Team, 2018). Infant cries were analyzed for spectral power in high (>5000 Hz), mid (500-5000 Hz), and low (<500 Hz) frequency bands using a Fast Fourier transformation using previously published methods. 18 Spectral analysis was selected to capture individual differences in the full range of acoustic frequencies, which include laryngeal and pharyngeal effects on cry features. High, mid, and low frequency cut offs were selected based on their association with auditory sensitivities in humans and their use for social and distress cues, with high frequencies having association with distress.19,20
Microbiome Analysis
An aliquot from the stool sample was stored at −80°C for subsequent microbiome analysis. Starting with 100–250 mg aliquot of stool, DNA was isolated using the DNeasy PowerSoil Pro HTP kit (Qiagen Inc., Germantown, MD, USA). Sample quality was assessed by MiSeq Nano. v2 Chemistry, library prep made using ¼ reaction Illumina Nextera XT, and paired end sequenced using the MiSeq 2 × 150 PE (Illumina, San Diego, CA) applying 31 samples per flowcell. DNA isolation and sequencing were performed at the University of Minnesota Genomics Center in Minneapolis, MN.
Quality control was performed using BBMap, BioBloomTools, and SHI7. SHOGUN was used to perform taxonomic assignment of the reads passing QC. In total, 5,621,003 reads (median: 143 732 reads per sample, range: 73 063 to 285 840 reads per sample, lower and upper quartile: 119 169 and 188 111 reads per sample) remained after quality control and taxonomic classification in the 37 study samples. All samples remained after filtering for low read count. Taxonomic classification identified 19 phyla, 417 genera, and 1246 species using the RefSeq Rep82 database.
SHOTGUN sequencing was used to examine all metagenomic DNA for identification and relative quantification of bacterial taxa. Reads were subjected to adapter trimming, quality trimming, and entropy filtering via BBDuk (v38.69). 21 The reads were then screened for host genomic content using BioBloomTools (v2.3.2) 22 with hg38 used as the reference. The screened reads were further processed with SHI7 (v1.0.0). 23 Taxonomic assignment was performed on the quality-controlled reads using SHOGUN (v1.0.8) 24 and RefSeq82 as the reference database.
Statistics
We evaluated three types of associations with the study variables of infant distress, parent-infant interactions, maternal distress, and maternal depressive symptoms to identify the component of the microbiome revealed by whole-genome shotgun sequencing.
For each of the variables of interest, we looked at three types of associations:
Beta-diversity, using Bray-Curtis and Jaccard distances (association measured using PERMANOVA, p < 0.05). The distances and tests were calculated using the vegdist and adonis functions from the R vegan package (v2.5-7).
25
Alpha-diversity, using Shannon and Inverse-Simpson indexes generated by the R phyloseq package (v1.34.0)
26
with associated measures using linear models, p < 0.05. Differential abundance analysis was performed using the R LinDA package (v0.1.0)
27
(FDR-adjusted p-value <0.1) with a prevalence threshold of 0.1, and a winsorization cutoff of 0.97. The differential abundance analyses for taxa identified as significant in the primary LinDA analysis was repeated using MaAsLin2,
28
another popular differential abundance analysis method based on different modeling assumptions and inferential strategies.
Cry data analyses were performed using linear models and base R code. All analyses were performed using version 4.0.3 of the R programming language.
Results
Demographics and clinical characteristics of the sample are summarized in Table 1. We recruited 31 parents and infants dyads and none were excluded The majority of the mothers did not have a psychiatric history (58%), while 25.8% had a history of depression and 16.13% had postpartum depression. During pregnancy, a minority used antidepressants and only one participant had a history of substance use. The median PHQ-9 score at 2 months was 1 (IQR = 2) with three scores missing.
Demographic Data, Child's and Mother's Characteristics Summary and Scores for PHQ-9 and PSI-SF.

PHQ-9 score ranges: 2 month = 0 to 13; 4 month = 0 to 10.
PSI ranges: Defensive Responding: 7 to 21; Parental Distress: 12 to 38; Parent-Child Dysfunctional Interaction: 12 to 29; Difficult Child: 12 to 35; Total Stress: 36 to 89.
Similarly, at 4 months the median score was 2 (IQR = 3) with 5 scores missing. PSI-4-SF total stress scores ranged between 36 and 89. The data is shown in Table 1.
Symptoms of maternal depression were associated with beta diversity of infant gastrointestinal microbiome (Figure 1). Bilophila wadsworthia had the greatest fold change and strongest association with maternal PHQ-9 scores at 2 months (log2 fold-change of 2.1, below our significance threshold (ie, FDR < 0.1)).

Maternal depression and Beta diversity of infant microbiome. Symptoms of maternal depression were associated with beta diversity of infant gastrointestinal microbiome. Differential abundance analysis of taxa in the feces of infants in relation to the depression scale PHQ-9. The PHQ-9 scores obtained at 2 months were associated with four species: Ruminococcus flavefaciens, Actinomyces johnsonii, Clostridium dakarense and Bilophila wadsworthia (the false discovery rate, FDR < 0.1).
To assess the beta-diversity, which measures the change in species diversity from one environment to another, the infant microbiome was separated into two groups according to the PHQ-9 score at 2 months: the first group having a score between 0 and 4 (no depression) and the second group with scores between 5 and 13 (mild to moderate depression). The overall median 2 month PHQ-9 score was 1 and the sample numbers for the two groups [0,4] and [5,13] were n = 20 and n = 6, respectively. For 4-month PHQ-9, the overall median score was 2 and the sample numbers for the [0,4] and [5,10] groups were n = 21 and n = 5, respectively. It was found that there was a statistically significant dissimilarity between with the 2 month PHQ-9 but not with the 4 month PHQ-9. Both Bray-Curtis and Jaccard analyses showed that 2 month PHQ-9 scores were associated with beta-diversity (p = 0.006 and p = 0.0005 respectively) (Figure 2).
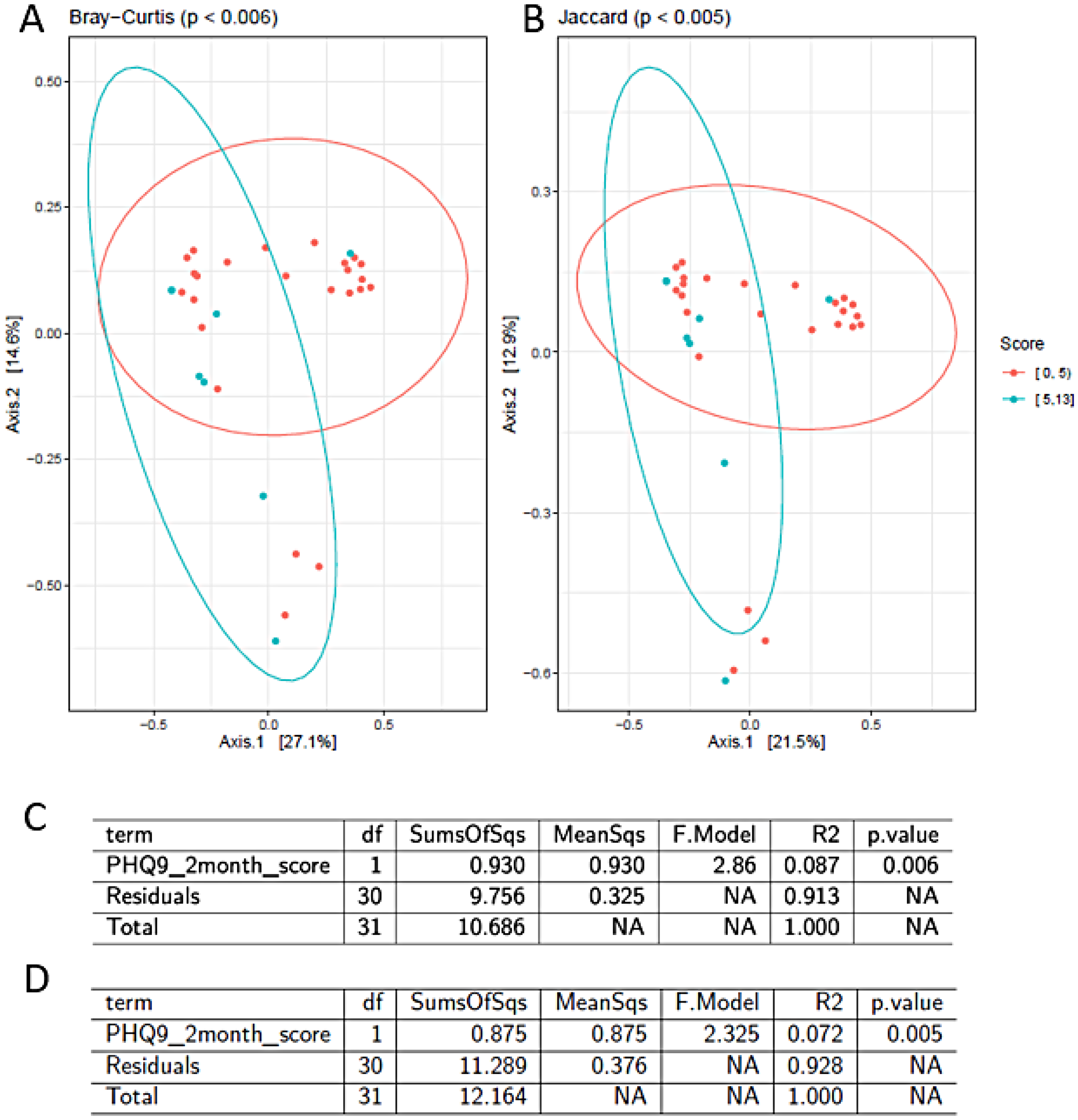
Separation of the infant microbiome based on the severity of maternal depression assessed by PHQ-9. Beta-diversity of the infant microbiome and maternal PHQ-9 score collected at 2-month were associated based on analysis. (A) Bray-Curtis and (B) Jaccard analysis. Orange dots represent individuals with no depressive symptoms (PHQ-9 scores between 0 and 4), Blue dots represents individuals with present depressive symptoms (PHQ-9 scores between 5 and 13). The tests reveal a statistically significant dissimilarity between the two groups (p < 0.05). Axes explain 27.1% (Axis 1 in Figure 2A) and 21.5% (Axis 1 in Figure 2B) of the variation, respectively. (C) Results of omnibus univariate PERMANOVA tests using features from beta diversity metrics and clinical indices of material depression assessed by PHQ-9. PHQ-9 scores were associated with beta-diversity via both Bray-Curtis and Jaccard distances analysis. The group-wise differences of beta diversity analysis showed significant associations via Bray-Curtis dissimilarity matrix (p = 0.006) and (D) the Jaccard distances (p = 0.005). Both analyses tested associations using A univariate PERMANOVA (p < 0.05).
Concerning the differential taxa abundances in the feces of the infants in relation to the post-immunization behavior based on the MAISD scale, only two maternal behaviors (kiss and distract) had significant associations with the infant microbiome diversity. Ten species were associated with distract behavior, with Actinomycetes urogenitalis and Actinomyces turicensis being the most abundant. On the other hand, eighteen species were associated with the maternal kiss, with Ruminococcus lactaris and Eubacterium hallii being the most common (Figure 3). Using MaAsLin2, the direction and overall pattern of associations were consistent with the LinDA results, and the key findings were qualitatively similar.
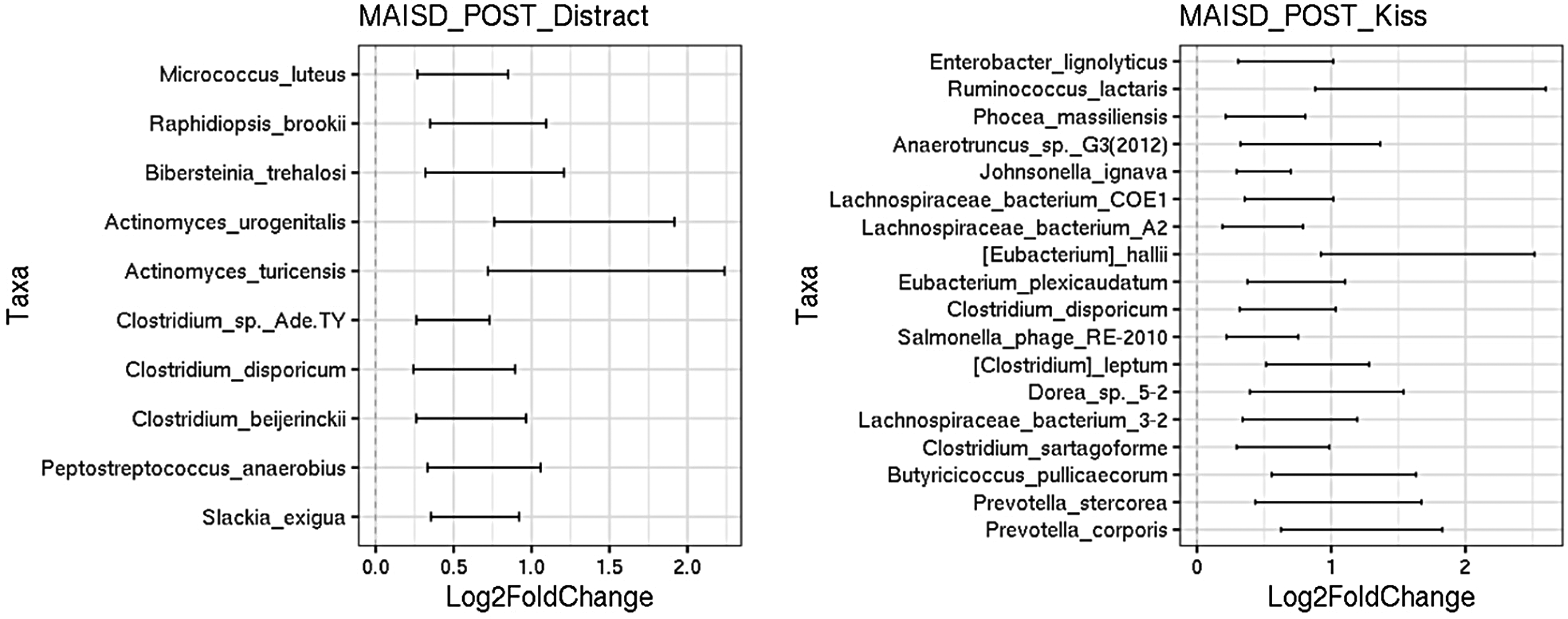
Differential taxa abundances are linked to post-immunization maternal behavior assessed by the MAISD scale. Box plot of the log2-fold-change in the relative abundance of taxa in the feces of the infants in relation to post-immunization (POST) behavior based on MAISD scale. The different bacterial taxa (Y-axis) were identified as significantly different along with the log2 fold change (X-axis). Two (distract, kiss) out of nine (Offer Object, Offer Pacifier, Offer Food, Nurse, Rub/Massage/Pat, Hug, Rock, Verbally Reassure, Not Visible) maternal behavior analyzes showed significant associations with infant microbiome diversity. (A) Association between distract behavior and bacterial taxa was found for 10 species. Actinomycetes urogenitalis and Actinomyces turicensis were the most abundant species. (B) Association between maternal kiss and bacterial taxa was found for 18 species. Ruminococcus lactaris and Eubacterium hallii had the largest effect size.
The Shannon diversity index was used to characterize the presence of differences in diversity of the species across the samples (Figure 4). Infant cries with greater high frequency band power (p < 0.03) and a greater high-to-mid frequency ratio–metrics (p < 0.05) were associated with altered α-diversity of the microbiome (Figures 4 and 5). It is also important to note that Bibersteinia trehalosi was the only species found to be associated with high frequency band power cry data.

The relationship between alpha diversity (via Shannon) and high frequency infant cry. (A) Observed values (circles), predicted values (connected by the blue line), and confidence intervals (gray shading). Infant cries with greater high frequency band power (p < 0.03) and a greater high-to-mid frequency ratio–metrics (p < 0.05) associated with higher infant stress were related to lower α-diversity. (B) Bibersteinia trehalosi enrichment was found to be associated with HighPow cry data.

Log2-fold-change in the relative abundance of taxa in feces of infants with high-to-mid ratio cry (FDR < 0.1). Infant cries with a greater high-to-mid frequency ratio–metrics (p < 0.05) were related to altered α-diversity of the microbiome. Each data point represents a different taxon (y-axis) identified as significantly different along with the log2 fold change (X-axis).
Discussion
This pilot study found that infant stress, as demonstrated by high frequency band power and high-to-mid frequency ratio-metrics with crying, was associated with lower alpha diversity, a marker of an unhealthy gut microbiome. Parental depressive symptoms were also associated with less microbiome diversity and unhealthy gut microbiome composition. It is important to identify biomarkers for gut microbiome health that can be measured in the clinical setting.
Our finding that infant distress, defined by crying quality is associated with lower alpha-diversity is consistent with previous research. 29 Even though a greater gut microbiome diversity is known to be beneficial to overall human health, it is still unclear whether it has any direct effects on health of the human brain and mental health. For example, in studies of people with depression, we see no alteration in alpha diversity, less diversity or high diversity. 30 The association between infant cry acoustic features and microbiome may be mediated in part by the vagus nerve, the function of which is responsive to stress and shaped by experiences. 31 Infant cry acoustic features are modulated by the larynx and pharynx, structures that are directly innervated by the vagus, which also forms afferent and efferent connections with the gut and forms a critical pathway of the microbiota-gut-brain axis. 32 Our population was healthy, term infants and future larger, cohort studies need to also evaluate infants who have experienced stress or have underlying medical conditions.
This study confirmed previous study findings that maternal depressive symptoms are associated with microbiome composition but is the first study to link this is increased diversity. Depressive symptoms as assessed with PHQ-9 scores obtained at 2 months were associated with 4 bacterial species. These findings weren’t found with the PHQ-9 scores at 4 months which may be secondary to the overall low (good) scores and our small sample size in this pilot study. The association of PHQ-9 scores with microbiome results needs to be explored in larger cohorts with more variability in PHQ-9 scores.
Previous studies have shown that maternal distress, including postpartum depression, increases the offspring's risk of developing mental health disorders (ie, attention-deficit/hyperactivity disorder, depression, anxiety, conduct problems), 33 however, the mechanism is still unknown. This study shows that beta diversity and composition of the infant fecal microbiota may be linked to maternal depressive symptom severity. This is partly consistent with prior literature which demonstrated an association with maternal depressive symptom severity and fecal composition but not diversity. 34 It is possible that different microbiome analysis methods may identify diversity differently. 35 Our study found an increased association between the PHQ-9 scores at 2 months and the pro-inflammatory and pathogenic bacteria, such as Ruminococcus flavefaciens, Actinomyces johnsonii, Clostridium dakarense from the Firmicutes phylum and Bilophila wadsworthia from the Proteobacteria phylum which had the greatest association to the PHQ-9 scores. These results are consistent with what was discovered previously, where an increased abundance of bacteria belonging to both Firmicutes and Proteobacteria phyla was linked to exposure to maternal postpartum depression symptoms. 34 It is important to note that Proteobacteria is gram-negative and has a lipopolysaccharide (LPS) membrane which confers virulence to the bacteria and increased Proteobacteria levels have been seen in patients with depression.36,37 Previous studies demonstrated that the increased abundance of bacteria belonging to the gram negative Proteobacteria phylum was linked to the exposure of maternal stress.9,38 However, our analysis found no significant correlation between the PSI-4-SF scores and bacterial taxa. A possible reason for this was the low level of overall maternal concerns on the PSI-4-SF questionnaires. These overall findings highlight the important point that it is critical that the optimal analysis methods for microbiome analysis are identified.
Undiagnosed and untreated depression in new mothers is detrimental to their health and their children. It is important for clinicians to screen for depression in new mothers as well as any signs of distress in their infants. Current screening methods of obtaining a maternal PHQ-9 during well-child appointments may have limitations as this relies on self-report from mothers who are likely multitasking and more focused on the needs of their infant.39,40 The present study implemented innovative screening methods with two scalable, noninvasive biomarkers suggesting that microbiome samples and infant cries during routine vaccination visits have merit for future study. With the growth of AI (Artificial Intelligence) and computational technologies, data obtained from these procedures could be analyzed rapidly and be readily available to guide clinicians in their decision-making processes as well as refine predictions.
The present study included an assessment of the maternal psychological state with two questionnaires: PSI-4-SF and PHQ-9. However, the study had limitations to note. First, the PHQ-9 had missing data, 3 missing scores at 2 months and 5 missing scores at 4 months. Second, the sample was relatively healthy with most PHQ-9 scores falling within the normal range. Therefore, the general population, which includes subjects with pathological conditions that might have more severe phenotypes, is not entirely represented in the current study population. A third limitation was reliance on self-report assessments of prenatal and maternal distress. Fourth, we did not track how long this stool was stored at room temperature before collection by the study team. The total time was less than 48 h and previous studies have shown reasonable microbiome stability over this time period. 41 Finally, with our pilot study approach and small study population, we were unable to adjust for possible mediators like mothers with underlying of medical issues, complications during pregnancy, and mode of infant delivery. Additionally, there are other modifiers that should be considered with future studies including infant feeding choice, infant antibiotic exposure (which may directly influence the microbiome and/or be a marker for infant stress), mother's stress during pregnancy, and mother's existing microbiome.
One future research direction would be to pool our data with other similar datasets to better evaluate co-variates and make the findings more robust. With this approach, effect estimates derived from comparable analyses across independent cohorts could be combined to improve robustness and generalizability. At the same time, direct pooling of microbiome datasets across studies is often challenging due to well-documented batch effects arising from technical heterogeneity, including differences in sample collection protocols, DNA extraction methods, sequencing platforms, library preparation, and bioinformatic processing pipelines. These sources of variation can induce systematic shifts in microbial profiles that may exceed or obscure true biological signals, making naïve data aggregation statistically inappropriate. For these reasons, direct combination of external datasets was not pursued in the current pilot study. We agree, however, that meta-analytic strategies represent a more statistically principled approach to leveraging multiple studies while accounting for between-study heterogeneity. In which effect estimates derived from comparable analyses across independent cohorts could be combined to improve robustness and generalizability.
Conclusions
The results of this study indicate a connection between higher stress in infants and lower microbiome diversity (when higher diversity is more desirable). Parental depressive symptoms were also associated with lower microbiome diversity and unhealthy gut microbiome composition. Additional larger cohort studies are needed to understand how the quality of the infant-caregiver relationship influences intestinal microbiome diversity.
Footnotes
Abbreviations
Acknowledgements
We wish to thank Juliet Dahlgren for manual checking and editing of audio files. We thank Jacek Kolacz, PhD, for his contribution in running the acoustic analysis for the study.
Ethical Statement
Prior to any research related activities, the study was approved by the Mayo Clinic IRB (17-010417). Prior to participation at least one parent completed written informed consent.
Authors Contributions
AL, MS, MPSF, SJ, JC, BAL, SVA, JF, JVV, PEC, MR collected the samples and produced the data. AL, BAL, PEC, MR designed the study. AL, MS, SJ, JC, PEC, MR analyzed data and wrote the paper. All authors reviewed and approved the final draft.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study is supported by the Palix Foundation.
Conflict of Interest Statement
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Dr Croarkin has received research support from the Agency for Healthcare Research and Quality (AHRQ), National Institutes of Health (NIH), National Science Foundation (NSF), Brain and Behavior Research Foundation, and the Mayo Clinic Foundation. Dr Croarkin has received research support from Pfizer, Inc. He has received grant-in-kind equipment support from Neuronetics, Inc. and MagVenture, Inc. for investigator-initiated studies. He received grant-in-kind supplies and genotyping from Assurex Health, Inc. for an investigator-initiated study. He served as the principal investigator for a multicenter study funded by Neuronetics Inc., a site principal investigator for a study funded by NeoSync, Inc., and a site principal investigator for a study funded by Innosphere. Dr Croarkin served as a paid consultant for Engrail Therapeutics, Meta Platforms, Inc., MindMed, Myriad Neuroscience, Procter & Gamble Company, and Sunovion. Dr Croarkin is employed by the Mayo Clinic. He receives compensation as the editor-in-chief for the Journal of Child and Adolescent Psychopharmacology.
Dr Romanowicz received grant-in-kind supplies from Kiddo Company, Inc. for an investigator-initiated study.
No other authors had any conflicts of interest to report.
Availability of Data and Material
Sequence files and metadata for all samples used in this study have been deposited in the BioProject ID PRJNA821920.