
Abstract
Background
Ketamine has rapid antidepressant effects and shows great promise as a novel treatment for depression, but its limitations including its abuse potential are poorly understood. Given that the prevalence of depression is twice as high in women as in men and that depression and substance use disorders are highly comorbid, we hypothesized that a sex-specific responsivity to behavioral assays that characterize addiction-like behavior may arise in rats with prior exposure to chronic stress and therapeutically relevant ketamine.
Methods
Male and female rats that underwent chronic mild stress were treated with four 1.47 mg/kg intravenous ketamine infusions once every fourth day and underwent operant self-administration of 0.5 mg/kg/infusion ketamine. Measures of anhedonia (or lack of pleasure, a signature feature of depression), anxiety-induced neophagia, motivation to obtain ketamine, and craving were assessed using the sucrose intake test, novelty-suppressed feeding test, progressive ratio schedule of reinforcement, and incubation of craving following abstinence, respectively. Finally, dendritic spine density in the nucleus accumbens core was measured.
Results
Ketamine infusions reduced anxiety-induced neophagia in both male rats and female rats but had no effect on measures of anhedonia. Female rats with prior exposure to chronic mild stress had greater motivation to obtain ketamine compared to nonstressed female rats, an effect not observed in male rats. Additionally, female rats who received antidepressant ketamine infusions had a higher threshold for displaying ketamine addiction-like behavior than saline-treated female rats as well as increased thin spine density in the nucleus accumbens core. These effects were not observed in male rats.
Conclusion
This study shows that repeated low-dose ketamine does not increase abuse potential of subsequent ketamine. It also highlights an important female-specific effect of stress to increase ketamine addiction-like behavior, which requires further investigation for clinical populations.
Introduction
Depression carries the highest global burden of disease of any mental illness.1,2 Treatment options are limited due, in part, to its heterogeneous symptomatology and high comorbidity with other conditions including substance use disorders. 3 The etiology of depression and substance use disorders is poorly understood, especially with respect to females who, despite having a twofold increased risk for depression, have been historically underrepresented in neuroscience research.4,5 Additionally, both human and rodent studies indicate that females escalate drug use more rapidly than males and may be more prone to relapse, depending on drug type.6,7 Preclinical research must address these intersecting factors underlying disease susceptibility.
The discovery of the rapid antidepressant effects of subanesthetic intravenous (i.v.) ketamine (KET) invigorated a field limited by monoaminergic antidepressant drugs that require several weeks of treatment to produce effects.8–10 However, KET’s therapeutic benefits are hindered by its abuse potential at high doses in humans, 11 and the fact that rats readily self-administer KET.12–15 Questions regarding KET’s safety require careful consideration before widespread integration into medical practice as an antidepressant. 9 This current work attempts to address two factors that may underlie susceptibility to KET’s effects: sex and prior exposure to chronic stress as a precipitating factor for both drug relapse and depression. We utilized the unpredictable chronic mild stress (CMS) model16,17 to induce a depressive-like behavioral profile in male and female rats, followed by i.v. slow, passive infusions of KET and subsequent operant KET self-administration to test the hypothesis that subjects—especially females—with a history of CMS and KET treatment would have an increased propensity to abuse KET.
Most behavioral models studying KET’s antidepressant-like effects have utilized intraperitoneal (i.p.) injections. In clinics, patients typically receive 0.5 mg/kg KET i.v. delivered over a 40-min period, two to three times per week. 9 Therefore, to more closely recapitulate clinical approaches, we used an i.v. dose of 1.47 mg/kg/40 min, previously shown to match N-methyl-D-aspartate receptor occupancy in rats equivalently to clinically relevant doses. 18
Dendritic spines are highly plastic structures that correlate with changes in synaptic strength. Both stress and exposure to drugs of abuse dynamically mediate spine density within corticolimbic structures, including the nucleus accumbens (NAc), where an increase is typically observed.19,20 Our laboratory demonstrated that sensitization to low-dose KET increases spine density in the NAc shell subregion of males and females, whereas only females had an increase in the NAc core (NAcc). 21 NAc spine density alterations following incubation of KET craving are unknown, but incubated cocaine craving results in persistent induction of NAc spine formation over the course of withdrawal, 22 associated with increased excitability and thought to mediate the persistence of craving. 23 Therefore, we assessed NAcc dendritic spine density following incubation of KET craving in stressed and KET pretreated rats.
In this work, we therefore asked whether or not limited and repeated treatments with low doses of KET enhance vulnerability to KET addiction-like behavior in both sexes of rats.
Materials and Methods
Drugs
Ketathesia (Henry Schein, Melville, NY) was diluted in sterile saline (SAL) from a 100 mg/mL racemic solution. For therapeutic infusions, KET was diluted to 1.47 mg/mL by body weight. For operant KET self-administration, 0.5 mg/kg/infusion in a 50 µL volume was used.
Subjects
Adult male and female Sprague-Dawley rats (initially seven to eight weeks old, weighing 226–250 g and 161–180 g, respectively) from Charles River (Raleigh, NC) were used. Animals were given four days to acclimate to the vivarium and were handled twice to acclimate to the experimenter handling. Males and females were same-sex double-housed in the same room (except where noted) in 43 × 21.5 × 25.5 cm Plexiglas cages on a 12/12 h light/dark cycle. Food and water were provided ad libitum except where noted. Behavioral experiments were conducted in six cohorts with each experimental group represented. All experiments were carried out in accordance with the National Institutes of Health Guide for Care and Use of Laboratory Animals 24 , and all protocols were approved by the Florida State University Institutional Animal Care and Use Committee.
Surgical Procedures
Jugular Catheterization and Catheter Maintenance
Surgery and catheter maintenance were performed as previously described, 12 with minor modifications, described in Supplementary Methods. Rats were allowed three to five days postoperative recovery before behavioral testing resumed.
Intra-NAcc Viral-Mediated Gene Transfer
Anesthetized rats were prepared for stereotactic surgery under standard sterile conditions. Two small craniotomies were made corresponding to NAcc coordinates: AP + 1.5, ML ± 1.2, and DV − 7.6. The viral construct herpes simplex virus (HSV)–cytomegalovirus–green fluorescent protein (GFP) (obtained from the McGovern Institute Viral Core Facility) was bilaterally delivered, 1 µL per side (1 × 109 units/mL), at 0.1 µL/min. After 5 min for virus distribution, the needles were raised, craniotomies were covered, and the incision was closed. Rats were allowed three days rest to allow for optimal HSV-GFP expression.
Experimental Procedures
Experimental Time line
See Figure 1 for layout of behavioral experiment and Supplementary Methods.
Sucrose Pellet Operant Training
Rats were shaped in operant chambers as previously described (see also Supplementary Methods). 12
Sucrose Intake Test
During testing, rats were separated into individual cages with two counterbalanced bottles: one containing water and one containing 1% w/v sucrose solution. Bottle weights were taken before and after the 12 h access period during the dark cycle. Three baseline measurements were taken before CMS onset, three tests occurred each week of CMS, and four tests occurred after each therapeutic KET infusion. Intake values are expressed as percentage change from the average of each animal’s three baseline measurements.
CMS Procedure
Rats underwent CMS or nonstressed (NS) conditions. The CMS procedure (outlined in Table 1 and adapted from Li et al. 25 ) utilized several different stressors presented pseudorandomly over 21 days, two stressors per day. Sucrose intake tests were conducted on days of social isolation. NS rats were single-housed and tested on matched days. For details, see Supplementary Methods.
Therapeutic KET Infusions
After CMS and jugular catheterization, rats underwent four i.v. KET or SAL infusions every fourth day for a total of 16 days. A 1 × 1 × 1 meter acrylic open-field arena divided into four quadrants was used as a neutral setting to administer infusions. Rats were tethered to drug delivery lines and given 5 min to habituate. KET (1.47 mg/kg/40 min) or SAL was delivered using a variable-speed syringe pump.
Novelty-Suppressed Feeding Test
This test was administered as previously described, 26 24 h after the first KET infusion. See Supplementary Methods for a detailed description.
KET Self-Administration
Twenty-four hours after the final sucrose intake test, rats began i.v. self-administration. The seven daily sessions consisted of 2 h access to 0.5 mg/kg/infusion KET under a fixed-ratio 1 (FR1) schedule of reinforcement, with 100 maximum possible infusions. The active nose-poke hole delivered i.v. KET and drug-paired cues identical to the sucrose pellet training sessions, while the inactive hole had no programmed response. This was followed by three daily progressive ratio (PR) sessions, where the response requirement for one infusion increased by 5 × e(0.2 n)−5, where n equals the infusion number. 27 Therefore, one infusion of KET required 1, 2, 4, 6, 9, 12, 15, 20, 25, 32, 40, … active nose pokes. The session ended if rats failed to complete a ratio in 1 h. Three more FR1 sessions followed, then rats underwent a period of home cage abstinence. Rats were returned to the operant chambers 1, 7, and 21 days after their final FR1 session to test for incubation of KET craving in 2 h sessions, where discrete cues were presented upon active nose poke, but KET was not available. Twenty-four hours after the final test for incubation of craving, intra-NAcc HSV-GFP was delivered as described above to animals randomly selected across each cohort.
Immunohistochemistry, Confocal Imaging, and Dendritic Spines Analysis
See Supplementary Methods for details. Briefly, animals were transcardially perfused three days after HSV-GFP injections, and tissue was processed for immunohistochemical detection of GFP. Z-stack projections were acquired on a laser-scanning confocal microscope, and dendritic branches were three-dimensional-reconstructed using Neurolucida 360 (version 2017.01.1; MBF Biosciences, Williston, VT). Automatic classification of spine type (thin, stubby, and mushroom type) was based on established parameters. 28 Data are presented as number of spines per 10 µm of dendrite segment.
Statistical Analysis
Two-way analysis of variance (ANOVA) was used to analyze cumulative body weight gain during CMS as well as novelty-suppressed feeding test (NSFT) feeding latency. For sucrose intake and self-administration data, mixed-model ANOVAs were used. Where appropriate, sex, stress, and treatment were the between-subjects factors and sessions were the within-subjects factor. To determine whether an increase in intake during FR1 KET self-administration occurred, the parameter estimates of the mixed model output were observed. Tukey’s post hoc was used when appropriate. Grubb’s outlier test was conducted for each behavioral test, and animals that were statistical outliers for one test were removed. Prism (GraphPad, version 8.01) and R (3.3.1) with the packages nlme 29 and lsmeans 30 were used for all statistical analyses and figures. Significance was set at α = 0.05.
Results
Sucrose Pellet Training
Rats reach criteria in three to six days; therefore, the last three sessions per rat were analyzed. Main effects of both sex and sessions were observed for sucrose pellet intake (F(1, 125) = 44.224, p < 0.0001; F(2, 254) = 77.431, p < 0.0001, Supplementary Figure 1A), active nose pokes (F(1, 125) = 33.815, p < 0.0001; F(2, 254) = 34.07, p < 0.0001), and inactive nose pokes (F(1, 125) = 30.937, p < 0.0001; F(2, 254) = 13.065, p < 0.0001), respectively (Supplementary Figure 1B, n = 26–41 per group). Therefore, while all rats increased their intake over time, females displayed overall heightened operant responding. Importantly, there were no significant differences a priori between stressed (CMS) and NS groups. Subsequent analyses of operant behavior were split by sex.
Weight Gain and Estrous Cycle During CMS
For weight gain during CMS, a significant interaction between sex and stress was observed (F(1, 123) = 26.81, p < 0.0001, n = 26–41 per group, Figure 2(a), such that CMS males gained less weight than NS males (p < 0.0001), while CMS females and NS females were not statistically different. Additionally, vaginal lavage of females during the last week of CMS/NS indicated that a majority of CMS females lost regularity in their estrous cycle (data not shown).
Time line of experiment. Depression-like behavioral assays including NSFT and sucrose intake test (S1–K4) are indicated in blue, while incubation tests D1, D7, and D21 are indicated in red. CMS: chronic mild stress; NS: nonstressed; KET: ketamine; SAL: saline; FR1: fixed-ratio 1 schedule of reinforcement; PR: progressive ratio; GFP: green fluorescent protein; HSV: herpes simplex virus; NSFT: novelty-suppressed feeding test. Consequences of CMS and subsequent KET treatment on depressive-like behaviors. (a) Cumulative weight gain during the CMS/NS procedure resulted in decreased weight gain in CMS males (***p < 0.0001, n = 26–41 per group). (b) Novelty-suppressed feeding test revealed an increased latency in CMS rats, which was abolished in KET-treated rats (*p < 0.05, **p < 0.01, pairwise group comparisons, n = 9–18 per group). Sucrose intake in males (c) and females (d), expressed as percentage change from baseline, shows that CMS reduced both males’ and females’ intake (*p < 0.05, n = 7–15 per group). Data are presented as mean ± standard error of the mean. CMS: chronic mild stress; NS: nonstressed; KET: ketamine; SAL: saline. Unpredictable chronic mild stress procedure.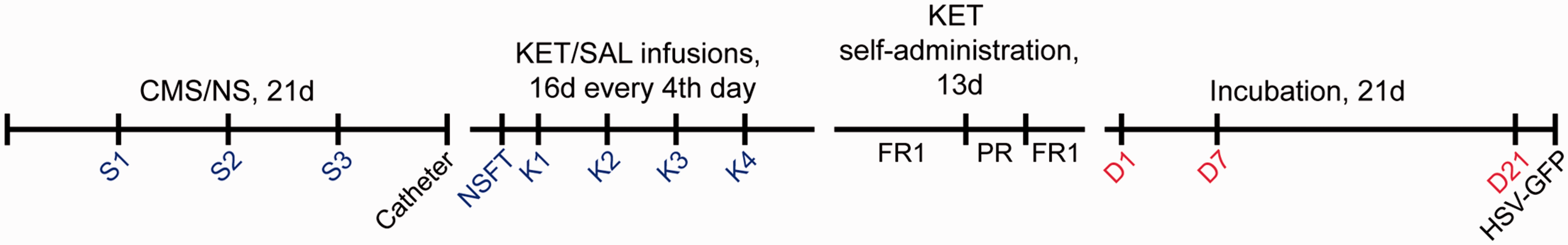


Novelty-Suppressed Feeding Test
Latency to approach, the food at the center of the arena (Figure 2(b)) revealed an interaction between stress and treatment for both males and females together (F(1, 119) = 7.35, p = 0.008, n = 9–18 per group). Since no effect of sex was observed, this factor was collapsed. Pairwise comparisons with Tukey-adjusted p values were conducted to analyze effects within levels. CMS + KET-treated rats had significantly shorter latencies than CMS+SAL-treated rats (p = 0.0002). CMS+SAL-treated rats had significantly longer latencies to approach the food than NS+SAL-treated rats (p = 0.017). Importantly, there were no differences between NS+SAL and NS+KET (p = 0.683) and CMS+KET versus NS+KET (p = 0.804). Together, this indicates that KET reduced NSFT latency in both sexes exposed to CMS.
Sucrose Intake Test
To determine if CMS induced anhedonia and if KET infusions ameliorated this effect, percentage change in sucrose intake compared to pre-stress baseline during CMS/NS and KET/SAL infusions (S1-3 and K1-4, respectively) was analyzed, shown in Figure 2(c)for males and (d) for females (n = 7–15 per group). There was a stress × sessions interaction for males and females (F(6, 263) = 6.163, p < 0.0001; F(6, 144) = 2.485, p = 0.026, respectively). Post hoc analyses revealed that CMS males had a significantly reduced intake compared to NS males during S2 and S3 (p < 0.05), and CMS females had a reduced intake versus NS females during all three sessions (p < 0.05). There were no differences between NS and CMS animals during the infusion period (K1–K4). Additionally, there was no effect of KET treatment for males or females, nor were there differences in water intake (data not shown). To account for body weight differences due to sex and/or stress conditions, we also analyzed sucrose intake normalized to body weight (data not shown). A main sex effect was observed (F(1,83) = 30.370, p < 0.0001). Females had a stress × sessions interaction (F(9, 334) = 2.548, p = 0.008), such that CMS had a lower intake than NS on S1, S2, and S3 (p < 0.05). Males had a significant stress × sessions interaction (F(9, 401) = 3.797, p < 0.0001), such that CMS had a lower intake than NS on S2 (p < 0.05). Together, this indicates that CMS resulted in decreased sucrose intake not explained by body weight, and that cessation of CMS resulted in a rescue of sucrose intake rather than an effect of KET treatment.
KET Self-Administration
For all KET self-administration comparisons, the number of subjects were n = 11–15 per group. For infusions during the first seven sessions, a main sex effect was observed (F(1, 103) = 14.043, p = 0.003). For males (Figure 3(a)), a main sessions effect was observed (F(6, 318) = 18.876, p < 0.0001), without stress or treatment effects, indicating an increase over time regardless of condition. For females (Figure 3(b)), a treatment × sessions interaction was observed (F(6, 275) = 7.276, p < 0.0001, right), such that SAL-treated females had higher intake than KET-treated females on sessions 6 and 7 (p = 0.034, p = 0.001, respectively), trending on session 5 (p = 0.082). Additionally, an overall stress effect was observed (F(1,48) = 6.391, p = 0.015), such that CMS females had a higher intake than NS females. For active nose pokes (Figure 3(c) and (d)), both males and females had a main sessions effect (F(6, 318) = 18.876, p < 0.0001; F(6, 275) = 18.695, p < 0.0001, respectively), without stress or treatment effects, indicating that active responding increased over time regardless of experimental manipulation. When comparing inactive and active nose pokes for both males and females (Figure 3(e) and (f)), nose poke type × sessions interactions were indicated (F(6, 689) = 8.659, p < 0.0001; F(6, 596) = 7.716, p < 0.0001, respectively). Post hoc comparisons indicated that at each session, active nose pokes were higher than inactive nose pokes (p < 0.01 for all time points), indicating a meaningful discrimination between the reinforced and nonreinforced operanda. Taken together, both sexes found KET reinforcing, but KET pretreated females administered less than SAL-treated females and CMS increased KET intake in females.
First seven days of FR1 KET self-administration. KET infusions for males (a) and females (b) indicate that CMS females have a higher intake and KET-treated females have a lower intake. Active nose pokes for males (c) and females (d). Inactive nose pokes for males (e) and females (f). **p < 0.001, main stress effect; *p < 0.05, KET- versus SAL-treated females within each session. Data are presented as mean + standard error of the mean, n = 11–15 per group. CMS: chronic mild stress; NS: nonstressed; KET: ketamine; SAL: saline.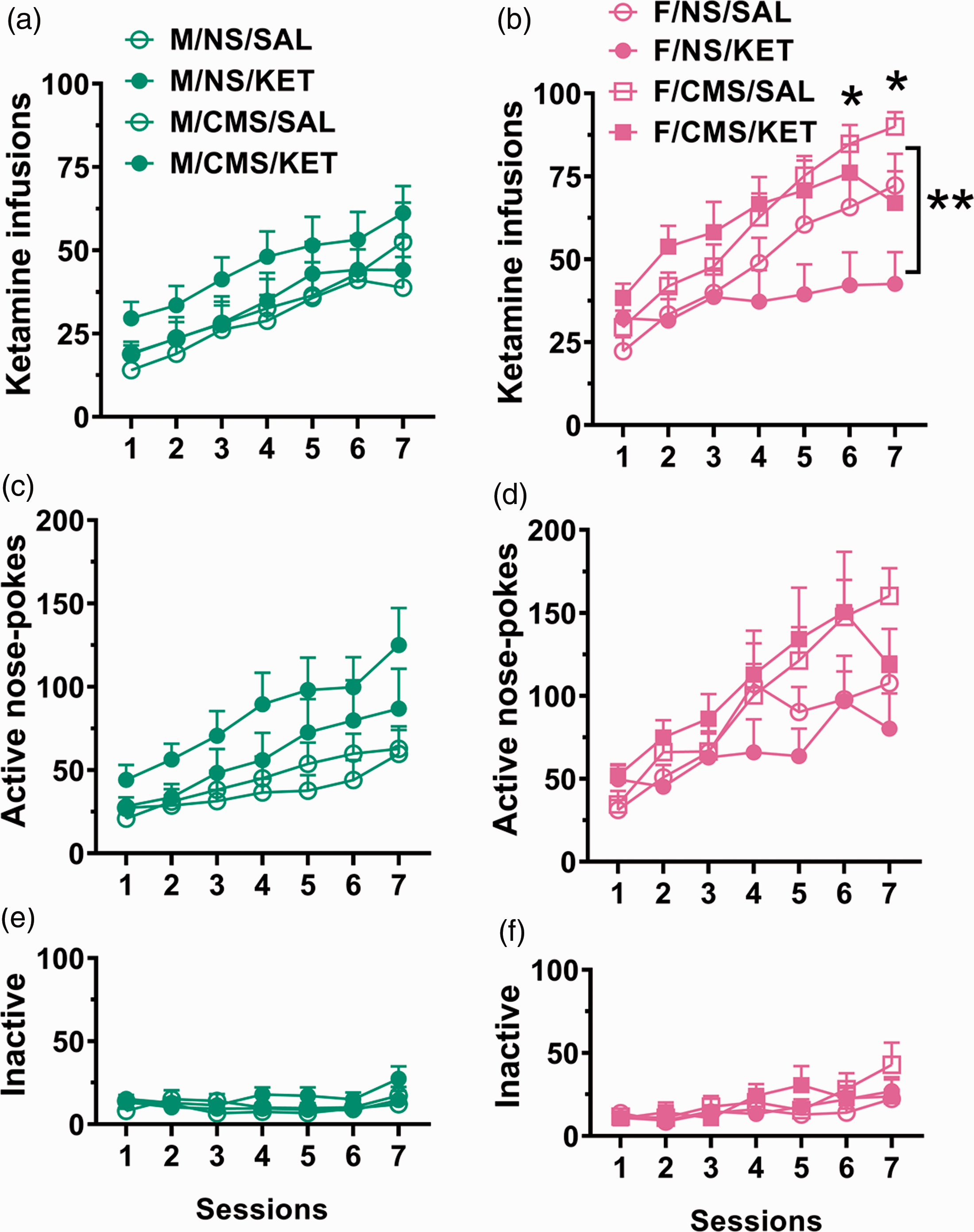
For active nose pokes during the PR sessions (Figure 4(a) and (b)), a main effect of sex was observed (F(1, 101) = 15.424, p < 0.0001). Males had a stress × treatment interaction (F(1,52) = 10.883, p = 0.002; post hoc tests show that KET-treated NS males had higher responses than KET-treated CMS males, p = 0.001), and females had a stress × sessions interaction (F(2,83) = 4.154, p = 0.019; post hoc tests indicates significantly greater response from CMS females than NS, particularly on session 2, p = 0.002). Both sexes successfully discriminated between active and inactive nose pokes (F(1, 506) = 48.933, p < 0.0001). Together, this indicates that the CMS procedure differentially affected males and females, where CMS females displayed higher motivation to obtain KET, an effect not observed in CMS males.
Motivation to obtain KET. Active nose pokes during PR sessions for males (a) and females (b). (a) **p < 0.005, stress × treatment interaction and (b) **p < 0.005, stress effect. Data are presented as mean + standard error of the mean, n = 11–15 per group. CMS: chronic mild stress; NS: nonstressed; KET: ketamine; SAL: saline; PR: progressive ratio.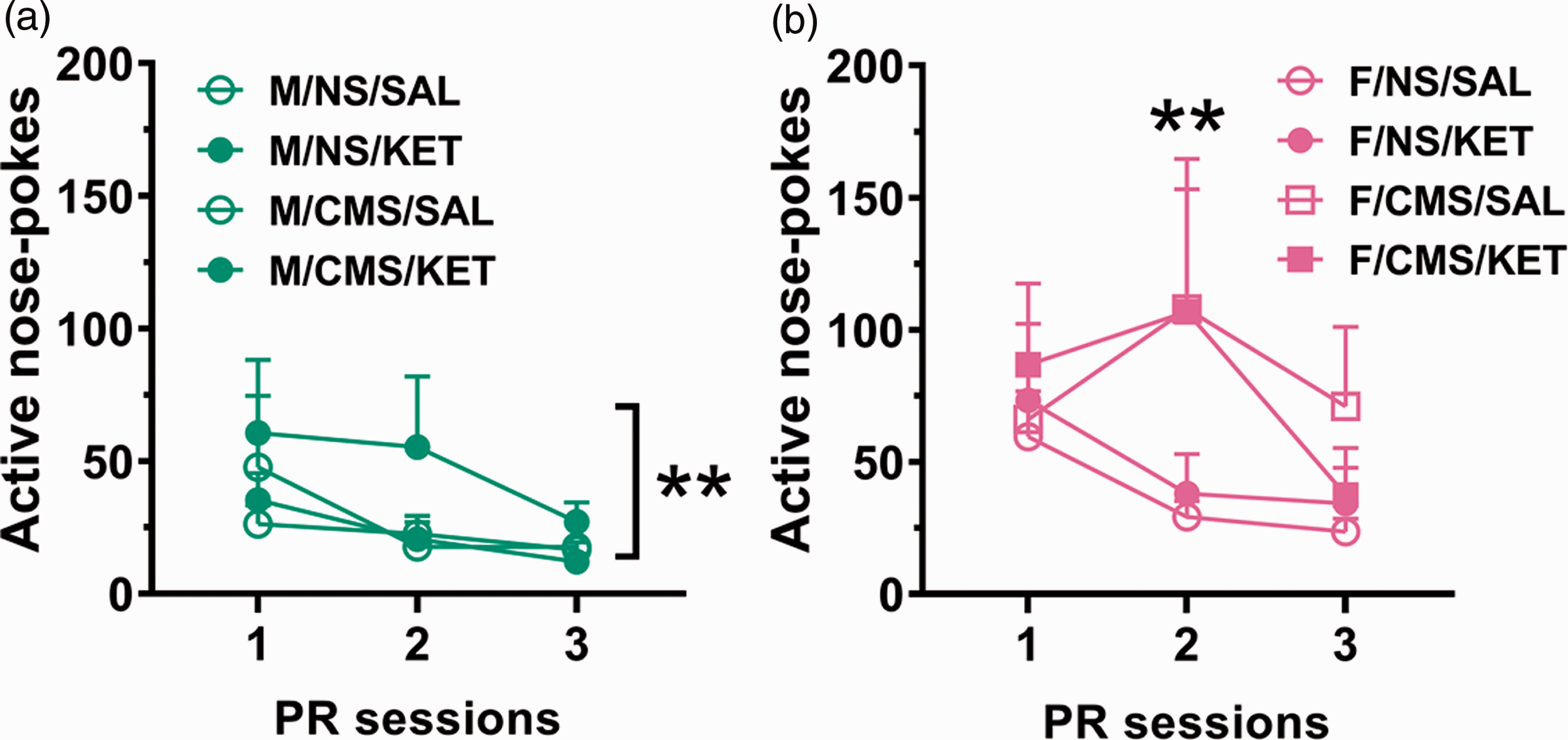
For infusions during the last three FR1 sessions (Figure 5), parameter estimates of the linear mixed model indicated that intake increased for both males and females over the course of the three sessions. Males had a main sessions effect F(2, 104) = 4.321, p = 0.016), and females had main stress, treatment, and sessions effects (F(1,44) = 9.398, p < 0.0001; F(1,44) = 9.335, p = 0.004; F(2,86) = 12.848, p < 0.0001), such that KET-treated females maintained their reduced intake compared to SAL-treated females (p < 0.001), regardless of stress condition. Active nose pokes revealed a main sessions effect for males (F(2, 104) = 6.783, p = 0.002) and a stress effect for females (F(1,44) = 7.618, p = 0.008). Inactive nose pokes were significantly lower than active nose pokes (F(1, 476) = 552.298, p < 0.0001), indicative of continued discrimination between reinforced and nonreinforced operant behavior.
Last three days of FR1 KET self-administration after PR tests. (a and b) Infusions, (c and d) active nose pokes, and (e and f) inactive nose pokes for males (a, c, and e) and females (b, d, and f). While all rats increase their intake over time, CMS females continue to have an increased KET intake compared to NS females. (a to c) ***p < 0.0001, main effect across sessions and (b and c) *p < 0.05, stress effect for females. Data are presented as mean + standard error of the mean, n = 11–15 per group. CMS: chronic mild stress; NS: nonstressed; KET: ketamine; SAL: saline; FR1: fixed-ratio 1 schedule of reinforcement.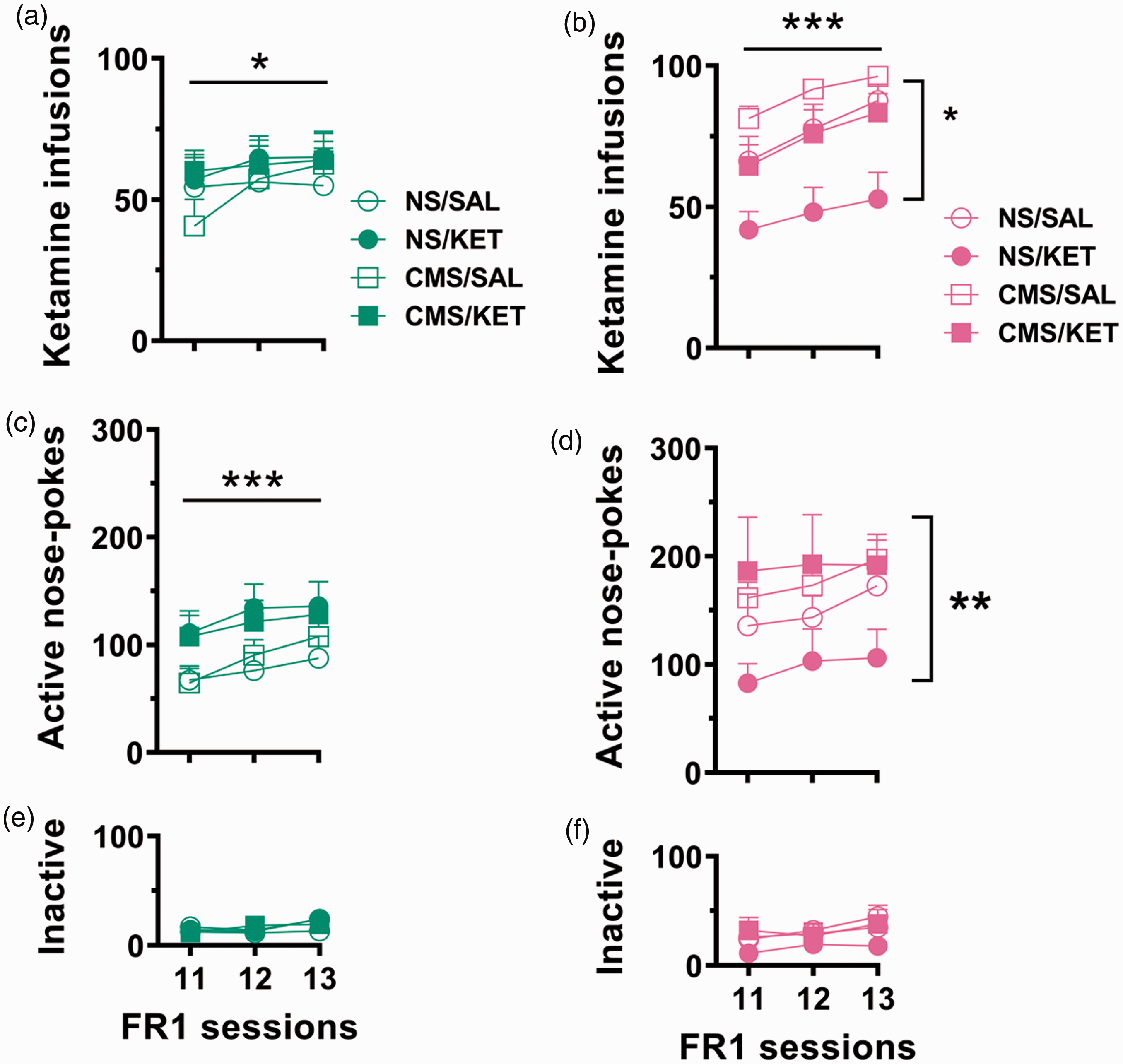
Active nose pokes during the three incubation tests were analyzed (Figure 6(a) and (b)). A main sex effect was observed (F(1,93) = 22.815, p < 0.0001). Males and females both had main effects across sessions (F(2, 105) = 25.577, p < 0.0001; F(2,84) = 16.703, p < 0.0001, respectively). There were also significant sessions effects for males’ inactive nose pokes (F(2, 110) = 5.402, p = 0.006) but not females. Importantly, an interaction between sessions and nose poke type was observed across all time points (F(1, 512) = 12.675, p < 0.0001). Taken together, both sexes show incubated craving of KET.
Incubation of KET craving. Males (a) and females (b) both show persistent craving for KET with each test. ***p < 0.0001, sessions effect. Data are presented as mean + standard error of the mean, n = 11–15 per group. CMS: chronic mild stress; NS: nonstressed; KET: ketamine; SAL: saline.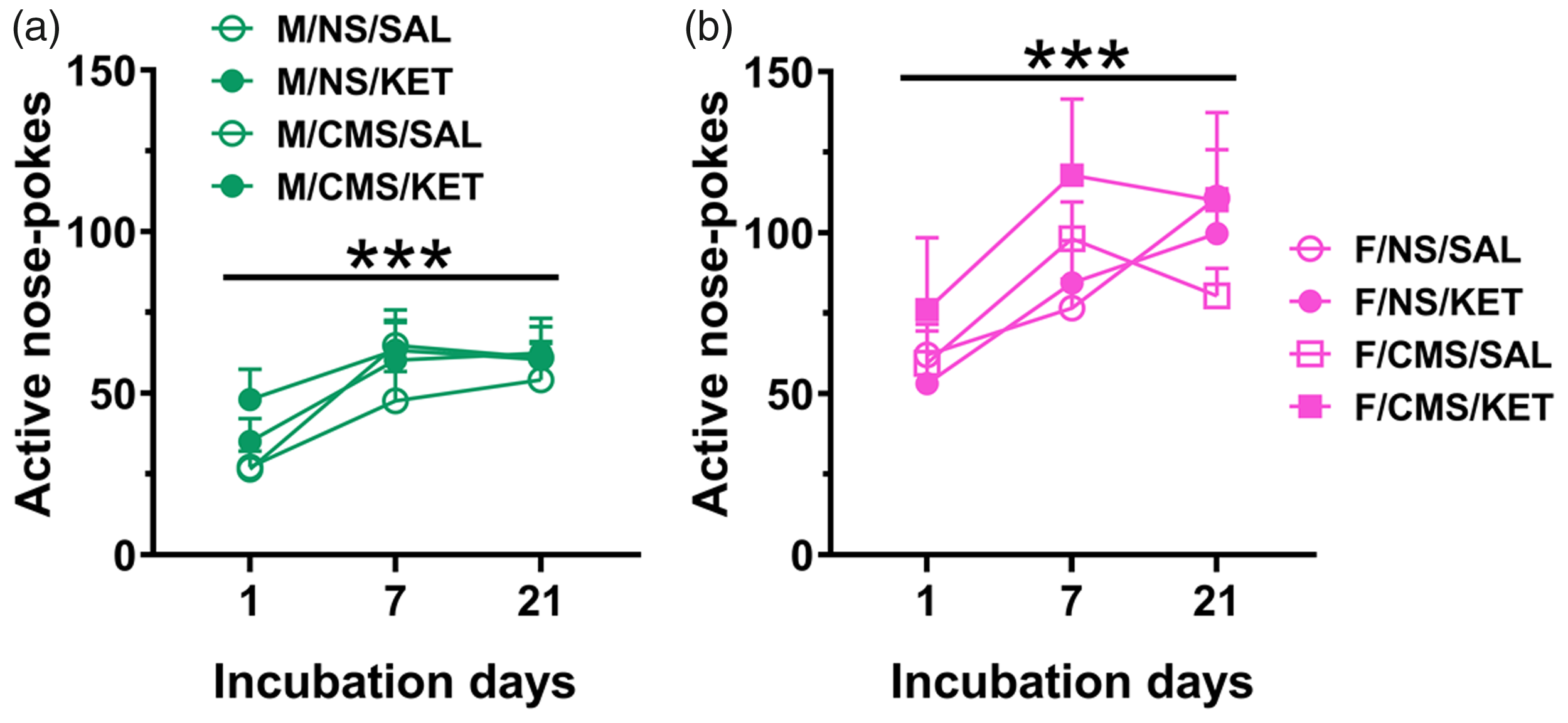
Spine Density and Morphology
Representative z-stack composite images from each group were analyzed (Figure 7(a), n = 4–6 per group). For total spine density (data not shown), there was no main effect of sex, stress, or self-administration. When separated by spine type (thin, stubby, and mushroom; Figure 7(b) and (c)), analyzing thin spines revealed a main effect of treatment (F(1,41) = 4.519, p = 0.039) such that KET treatment increased spine density, with no sex or stress effects. Split by sex, females had a trending effect of treatment (F(1,19) = 4.049, p = 0.059), while males had no significant group effects. There were no differences in mushroom-type or stubby-type spine densities as a function of sex, treatment, or stress conditions. Taken together, these results indicate that KET pretreatment increases dendritic spine density in the NAcc, an effect driven by thin spine formation.
Dendritic spine density in the nucleus accumbens core (NAcc). (a) Representative images of z-stack composites featuring dendritic spines in the NAcc. Scale bar = 10 µm. Stress (+= CMS,−= NS), KET treatment (− = SAL, + = KET), and KET self-admin (− = SAL self-administration, + = KET self-administration). (b and c) Spine density broken down by subtypes for males (b) and females (c). p = 0.05, main KET effect for females. When collapsed by sex, a main KET effect was observed (p < 0.05). Data are expressed as number of spines per 10 µm of dendrite + standard error of the mean. n = 4–6 rats per group, eight branches per rat. CMS: chronic mild stress; NS: nonstressed; KET: ketamine; SAL: saline.
Discussion
The purpose of this study was to assess the effects of stress exposure and KET pretreatment in males and females on measures of KET addiction-like behavior and changes in dendritic spine density in the NAcc. KET treatment resulted in a partial amelioration of depression-related behaviors and an increase in thin spine density in the NAcc. CMS enhanced KET addictive-like behavior in females only. Finally, females with prior KET treatment displayed decreased FR1 KET intake. This is the first study to investigate sex differences in KET addictive-like behaviors within the context of stress.
In the current study, first-pass indicators of CMS impact included reduced body weight gain in males and estrous cycle disruptions in females. The lack of body weight gain changes in females was expected and has been previously demonstrated, as females do not gain as much body weight as their age-matched male counterparts. 31 Anehdonia, a hallmark of depression, was assessed using the sucrose intake test, where a decrease in consumption of a sweet solution is usaally reduced after CMS. 16 In the present study, CMS reduced total sucrose intake in both males and females as compared to same-sex NS counterparts. This is a significant indicator of CMS’s impact on hedonic consummatory behavior and is in line with findings from other species exposed to stressful stimuli. 32 Importantly, the decreased intake was still observed after normalizing intake by body weight, suggesting the effect is not due to secondary impacts of CMS.
KET treatment had no effect on sucrose intake; rather, cessation of CMS returned sucrose intake to levels comparable to NS counterparts. Our 21-day CMS protocol was adopted from Li et al. 25 , where acute KET (10 mg/kg i.p.) increased sucrose preference in CMS male rats lasting up to 7 days. Similar findings were seen with male mice and sucrose intake.33,34 However, others found no effect of KET in male or female mice on CMS-induced anhedonia, 35 and only a transient effect of KET on sucrose preference in socially isolated males, but not females. 36 Another depression model, vicarious social defeat, 37 produces a depressive-like behavioral profile (including anhedonia) that is reversed by KET in female mice, 38 suggesting that stressor type can impact KET’s effectiveness. There are notable methodological differences among these studies that could explain these discrepancies. Drugs delivered via i.p. injection undergo initial absorption in the liver before reaching the brain, 39 therefore introducing greater possible effects of drug metabolites. Together, KET’s effects on anhedonia in preclinical studies remain unclear. As these measures were originally developed to test the efficacy of traditional (i.e., monoaminergic) antidepressants predominantly in males, more work is needed to optimize procedures testing rapid-acting antidepressants in females.
The NSFT demonstrated KET’s ameliorating effects on CMS-induced neophagia in both males and females, in line with previous findings.25,26,40 While these studies utilized i.p. injections of acute KET, this study uses i.v. infusions at a clinically relevant dose to reverse CMS-induced behavioral deficits in both sexes.
Rats underwent operant KET self-administration. Due to females’ higher sucrose pellet intake during initial shaping, analysis for males and females was conducted separately. Pharmacokinetic sex differences lend further support for separating the analysis for self-administration by sex, as females have slower clearance and longer elimination half-lives than males. 41 Females previously exposed to CMS displayed greater addiction-like behavior than NS females, demonstrated by increased intake during FR1 and enhanced motivation during PR sessions, an effect absent in CMS males. The dose of KET used for self-administration (0.5 mg/kg/inf) is a high dose commonly used.13,15,42,43 However, none of the aforementioned studies included females or tested incubation of craving. This is the first study to demonstrate persistent, incubated craving of KET after 7 to 21 days of forced abstinence. Animals show incubation of craving to many other drugs of abuse with differing temporal profiles. 44 In the current study, the expression of craving plateaus between the D7 and D21. It is well known that stress is a potent trigger for drug craving and precipitation of relapse in humans, especially for women. 45 Furthermore, female rodents are more susceptible to stress-induced augmentation of addiction-related behavior.46–49 This could be due to a positive interaction between circulating estradiol and extracellular dopamine in the NAc following chronic stress. 48 Indeed, behavioral responsivity to KET is influenced by gonadal hormones.50,51 More work is needed to understand the role of gonadal hormones in mediating stress-induced drug-seeking behavior, especially regarding KET.
During FR1 sessions, all animals increased their KET intake over time and maintained those levels after PR testing, indicating robust reinforcement. Among males, there were no differences due to stress conditions or prior KET treatment. Conversely, KET-treated females (regardless of stress condition) reduced their KET intake relative to SAL-treated females after the fifth FR1 session that persisted after PR testing. This suggests that KET pretreatment may have enhanced the motivation to obtain high doses of KET, since their PR and incubation of craving were similar to animals with high intake during FR1. Future studies involving extended access paradigms will help to further characterize this effect.
KET preexposed animals also had increased density of thin spines in the NAcc following incubation of KET craving. Immature spines (thin and stubby subtypes) are more motile and dynamically regulated as compared to the more stable mushroom spines. 52 These alterations correlate with increased synaptic excitability, as more synapses form and more glutamate receptors are inserted into the postsynaptic membrane, 53 thereby altering subsequent responsivity to drug- or stress-paired stimuli. NAc medium spiny neurons undergo robust structural and physiological changes following exposure to both drugs and chronic stress: in both cases generally, thin and/or stubby dendritic spine densities increase.20,54–56 While our results for males are in accordance with a previous study using socially defeated rats, 57 our results in females were unexpected because the last therapeutic KET infusion occurred 40 days prior to analysis. Additionally, another study with CMS males found decreased NAc spine density 58 as well as a clinical study showing decreased NAc volume following KET infusion. 59 In addition to methodological differences, our effect in females may be an additive result of KET treatment and KET self-administration. Previously, we reported increased spine density in the NAcc only in females, while both sexes had increases in the shell 2 h after an acute KET challenge (i.p.). 21 This effect of KET pretreatment on NAcc spines suggests long-lasting neuroadaptations, possibly due to upstream regulation of the medial prefrontal cortex and hippocampus, where KET reverses stress-induced deficits in spineogenesis. 60 Although these changes appear to be independent of the expression of KET addiction-like behavior, they may be related to the decreased FR1 intake observed in females and are unlikely to be linked to a specific antidepressant/therapeutic behavioral effect due to lack of specificity to stressed females and males.
Taken together, we have demonstrated a sex-specific responsivity to KET addiction-like behavior following exposure to chronic stress in females, an effect not observed in male counterparts. Additionally, using a novel i.v. KET treatment protocol similar to clinically relevant dosing, we showed that KET treatment partially ameliorates depression-related effects of CMS in both sexes without increasing propensity to self-administer KET as well as increased NAcc dendritic spine density. Finally, KET-treated females displayed decreased KET FR1 intake. Together, these findings suggest a differential response to KET self-administration that was not manifested in the expression of addiction-like behaviors but may still underlie neuroadaptations in reward circuitry relevant to responsivity to stress or drug-related stimuli. While these findings highlight an important question regarding the abuse potential of repeated low-dose therapeutic KET in rats, these factors require investigation in clinical population.
Supplemental Material
Supplemental Material1 - Supplemental material for Sex-Dependent Ketamine Addiction-Like Behavior Profile Following Exposure to Chronic Mild Stress
Supplemental material, Supplemental Material1 for Sex-Dependent Ketamine Addiction-Like Behavior Profile Following Exposure to Chronic Mild Stress by Katherine N. Wright, Devin P. Hagarty, Caroline E. Strong, Kristofer J. Schoepfer and Mohamed Kabbaj in Chronic Stress
Supplemental Material
Supplemental Material2 - Supplemental material for Sex-Dependent Ketamine Addiction-Like Behavior Profile Following Exposure to Chronic Mild Stress
Supplemental material, Supplemental Material2 for Sex-Dependent Ketamine Addiction-Like Behavior Profile Following Exposure to Chronic Mild Stress by Katherine N. Wright, Devin P. Hagarty, Caroline E. Strong, Kristofer J. Schoepfer and Mohamed Kabbaj in Chronic Stress
Footnotes
Authors’ Contributions
K. N. W. and M. K. conceptualized and designed the experiments; K. N. W., D. P. H., C. E. S., and K. J. S. conducted experiments; K. N. W. and D. P. H. ran statistical analysis; K. N. W., D. P. H., and K. J. S. wrote the manuscript; all authors provided critical feedback on the writing of this manuscript.
Acknowledgments
The authors would like to thank Rachael Neve from the Viral Gene Transfer Core, MIT, for the viral constructs, and Ruth Didier for her assistance with the confocal microscopy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by R01MH087583, R01MH099085, and R01 DA043461 to M. K.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.