
Abstract
Exposure to chronic stress, either repeated severe acute or moderate sustained stress, is one of the strongest risk factors for the development of psychopathologies such as post-traumatic stress disorder and depression. Chronic stress is linked with several lasting biological consequences, particularly to the stress endocrine system but also affecting intermediate phenotypes such as brain structure and function, immune function, and behavior. Although genetic predisposition confers a proportion of the risk, the most relevant molecular mechanisms determining those susceptible and resilient to the effects of stress and trauma may be epigenetic. Epigenetics refers to the mechanisms that regulate genomic information by dynamically changing the patterns of transcription and translation of genes. Mounting evidence from preclinical rodent and clinical population studies strongly support that epigenetic modifications can occur in response to traumatic and chronic stress. Here, we discuss this literature examining stress-induced epigenetic changes in preclinical models and clinical cohorts of stress and trauma occurring early in life or in adulthood. We highlight that a complex relationship between the timing of environmental stressors and genetic predispositions likely mediate the response to chronic stress over time, and that a better understanding of epigenetic changes is needed by further investigations in longitudinal and postmortem brain clinical cohorts.
Keywords
Introduction: Defining chronic stress
Exposure to chronic stressors across the human lifespan can multiply an individual’s susceptibility to adverse medical conditions.1,2 In this article, chronic stress refers to (1) a series of intense traumatic events such as accidents, physical assault, sexual assault, natural disasters, or combat exposure, leading to psychopathologies such as complex post-traumatic stress disorder (PTSD) 3 or (2) non-traumatic but major events in life, whereby an individual is exposed to sustained periods of stress, for example, caregiving, difficult divorce, or a stressful work environment leading to burnout. 4 Chronic stress is a robust risk factor for many medical conditions including cardiovascular disease, obesity, cancer, and immune disorders. 5 It is also highly linked with the development of a broad range of psychiatric illnesses, including PTSD, 6 major depressive disorder, 6 bipolar disorder, 7 schizophrenia, 7 anxiety disorders, 8 substance abuse disorders, 9 and addiction. 10
Chronic stress and trauma are associated with several lasting biological changes, particularly of the stress endocrine systems. For example, enduring stress and trauma can lead to permanent changes in the structure of several brain regions important for processing traumatic information (e.g. decreased somatosensory field of genital region after childhood sexual abuse), 11 which might be an adaptive response to protect an individual from future adversity.12,13 Interestingly, such adaptive mechanisms appear to occur in all individuals exposed to similar levels of stress or trauma, regardless of diagnostic outcome. Yet, only a fraction (∼20%) of people who have experienced trauma go on to develop PTSD, while some even experience post-traumatic growth. 14 Additionally, some individuals may not develop PTSD but other psychotic, anxiety, or affective disorders—or a combination of these—highlighting the heterogeneity of responses to stress.15,16 Considering that the outcomes following chronic stress exposure differ greatly between individuals, susceptibility to adversity resulting in psychopathology might be determined by how efficiently an individual is able to compensate for adversity at the molecular level.
In addition to genetic factors, some of the most relevant molecular mechanisms that may contribute to risk or resilience to the effects of stress and trauma are epigenetic. For example, DNA methylation profiles in monozygotic twin pairs diverge as twins grow older, with the differences between these genetically identical individuals at least partially explained by differing environmental exposures such as psychological stress. 17 Further, in genome-wide association studies of stress-related psychiatric disorders, single genetic variants explain only a small proportion of disease risk and overall do not completely explain heritability.
Epigenetics refers to the mechanisms that regulate genomic information by dynamically changing the patterns of transcription and translation of genes without altering the underlying DNA sequence. This occurs by altering the accessibility of DNA to transcriptional regulators by: (i) post-translational modifications of histone proteins, (ii) the addition of chemical groups most commonly at cytosine-phosphate-guanine (CpG) sites (e.g. methylation, hydroxymethylation, or other modification), or (iii) the binding of non-coding RNAs to specific sequences in DNA. A growing body of evidence convincingly shows that exposure to adversity can cause long-lasting epigenetic changes that lead to widespread alterations of gene expression. Such alterations are observed in both brain and peripheral tissues, indicating the widespread effects of stress throughout the whole organism. Epigenetics thus provides a mechanism by which the environment can have a global and lasting influence on the expression of genes in an individual following stress exposure, with epigenetic processes believed to underlie associations of disease burden, environmental risk, and individual phenotype. 18 Elucidating the interface of gene and environment interactions can help lead to a better understanding of the development of psychopathologies, and how they may be prevented or treated. In this review, we summarize preclinical and clinical research findings that support the existence of epigenetic modifications, particularly DNA methylation, in response to conditions of chronic stress, highlighting the effects of chronic and traumatic stressors occurring at different developmental time points and how this may lead to vulnerability to severe psychiatric disorders.
Epigenetics: Overview and Relevance
For environmental exposures to have lasting consequences on biological systems, alterations in gene transcription are required. This can occur by epigenetic modifications that influence chromatin state, and thus the accessibility of transcriptional machinery to regulatory regions such as gene promoters. Epigenetic modifications can be long lasting and can elicit either targeted or genome-wide changes in gene expression by processes including DNA methylation, post-translational histone modifications, and non-coding RNA regulation. One of the best understood epigenetic mechanisms involves the methylation of cytosine (5-methylcytosine or 5mC) on DNA, which occurs most abundantly at cytosines followed by guanine residues.19,20 In most instances, the presence of methylated cytosine recruits repressor complexes that modulate gene transcription by altering chromatin and inhibiting accessibility of the promoter to transcription factors and enhancers. 21 Opposite effects have been reported in a site-specific manner, with inhibition of repressor binding or enhancement of methylation-specific enhancers.22,23 DNA methylation is considered to be a stable epigenetic mark in post-mitotic cells, although increasing evidence indicates that 5mC and 5-hydroxymethylcytosine (5hmC; the stable intermediary between methylation and de-methylation) are dynamic, reversible, and modifiable under certain conditions.20,21
Patterns of gene expression are also regulated by histone modifications, 24 which involve the covalent modification of histone amino-terminal tails by, for instance, methylation, acetylation, phosphorylation, or ubiquination among others. Histone modifications can alter the level of chromatin condensation by either modifying electrostatic charge or recruiting epigenetic “readers” or “erasers” that bind to histones to, respectively, increase or decrease histone modifications. Epigenetic modifications to transcription and translation also extend to the effects of non-coding RNAs. These are short RNA species that act post-transcriptionally to repress protein synthesis, often by inducing degradation of mRNA targets. 25 Patterns of epigenetic modifications are able to elicit functional effects that result in either gene expression repression or enhancement.
Several genetic variants are consistently shown to interact with stress and trauma, with some of these effects long lasting. For example, in rodents exposed to stress early in life—such as stress caused by changes in maternal behavior—the epigenetic landscape of prominent “stress genes” such as NR3C1 and CRH can be altered, with these effects lasting into adulthood.26,27 Similar epigenetic patterns have been observed in humans exposed to childhood maltreatment who develop a mood disorder or PTSD later in life, 28 and epigenetic alterations are found in adults exposed to periods of chronic stress including combat exposure, 29 employment in a high-stress environment, 30 or a high lifetime stress load. 31 Thus, chronic stress and trauma are environmental exposures that can cause lasting epigenetic changes and lead to the development of psychiatric pathology.
A number of studies, ranging from twin to molecular genetic studies in large longitudinal cohorts, indicate that the impact of adverse life events is likely moderated by genetic variants, in the context gene × environment (G×E) interactions.32,33 The studies describe, however, only statistical interactions by nature. Epigenetic marks could provide a mechanism that may link how effects of genetic variants and environmental stimuli converge to regulate gene transcription and in consequences change in brain structure, endocrine responses, or behavior that have been reported following stress exposure. Control of gene expression by epigenetic modifications modulates important cellular processes including neurogenesis, neurotransmission, and synaptic plasticity, which are all disrupted in conditions of sustained stress.34,35 Such effects may influence an organism at not only a molecular level but also at circuitry and systemic levels.
Some evidence suggests that epigenetic modifications may be sustained during cell division and possibly transmitted across generations.32,33 This is important in the context of chronic stress exposure, as progressive but sustained epigenetic changes may be cumulative and could even be a result of prior chronic stress exposure in the parent. Several studies based on periods of deprivation, such as the Swedish famine, 36 Dutch Hunger winter, 37 Montreal ice storm,38,39 and the Holocaust, 40 provide some support for intergenerational transmission of traits through epigenetics, through investigating the offspring of parents who were subject to this trauma. However, the strongest evidence for epigenetic transgenerational inheritance comes from animal studies of paternal transmission, which show that the offspring of chronically stressed adult male mice are more vulnerable to stressful stimuli, have dysregulated stress responses, and altered hypothalamic-pituitary-adrenal (HPA) axis activity compared to control mice.41,42 It has been hypothesized that these apparently heritable effects may be underpinned by an epigenetic mechanism transmitted via sperm. 43 However, these studies remain highly controversial in light of currently accepted models that epigenetic modifications are erased during meiosis.44,45 Further, it remains unclear whether transgenerational stress-related behavioral alterations are truly epigenetic imprinting or a post-conception consequence of change in the uterine milieu or parental behavior.
Several caveats of epigenetic studies should be mentioned here. Firstly, human and animal studies investigating stress-induced epigenetic modifications for the most part report associative or correlational effects, rather than causal effects. They also often do not elucidate the mechanism by which the epigenetic change is induced nor its functional consequence. Secondly, while DNA methylation levels for some CpGs are correlated across brain and blood or saliva and blood,46–48 most are not. However, specific genetic variants or environments that cause the release of hormones with system-wide effects may have common effects on DNA methylation across tissues that may only differ in magnitude.49,50 There is also some evidence that overall, DNA methylation derived from buccal cells may be more closely related to brain, as both tissues are derived from the ectoderm. However, compared to the low relatedness between brain and whole blood, the improvements are small. 48 Additionally, in complex tissues such as the brain, epigenetic mechanisms also differ according to brain regions, sub-brain regions, and cell types. 51 Thus, results from epigenetic studies in peripheral blood or saliva may have to be interpreted with great care with regard to their reflection of causal effects on disease risk.
Methylation-wide approaches have the advantage of giving a broad picture of the methylome, but there are some important considerations for each method. While array-based methods are cost-efficient and technically robust, data pre-processing—including normalization and batch-correction—can have strong influences on the data, especially if small changes are expected. Several publications thus advocate that studies using array technologies should focus on results that survive multiple correction methods.52–54 Further, currently used methylation arrays focus on promoter regions, with lower coverage in enhancers that appear critical for stress-related impact. Sequencing approaches may therefore seem optimal; however, these methods are also subject to limitations. Firstly, DNA methylation does not vary at the majority of CpGs in a given tissue and so untargeted sequencing-based methods may result in lower sensitivity with less reads per CpG generated, reducing cost-efficiency. Importantly, a bias can also be observed with bisulfite conversion. 55 Direct sequencing of unmodified DNA using single molecule real-time sequencing methods might be a superior approach and is being developed by a number of companies. In addition, there is a push for the development of arrays that focus on additional CpGs, and other bases not covered by current array technologies. 56
An important requirement of the field is now to thoroughly catalog which epigenetic modifications occur in response to which types of stress (e.g. nature vs. timing vs. length of stress) in which different cell-types and tissues, and how this characterizes risk profiles and psychopathologies. The development of targeted, highly sensitive DNA methylation detection methods will propel this effort.
Epigenetic Mechanisms Early in Life and Lasting Effects of Early Life Adversity
Stress occurring early in development can cause dramatic effects on physiology and behavior. 57 In this context, early epigenetic changes in the brain are important factors. Changing methylation patterns have been observed in promoter regions of many genes across healthy brain development, especially in the human dorsolateral prefrontal cortex, a brain region which is late to mature and highly implicated in neuropsychiatric disorders. 58 Specifically, a switch from de-methylation to increased methylation of many genes at the genome level characterize the transition from fetal to childhood time points. 59 It is likely that interruption of normal developmental epigenetic patterns by stress or trauma early in life may alter epigenetic trajectories and lead to the development of psychopathology later in life.
A growing number of associations support the link between stress or trauma exposure and the occurrence of widespread epigenetic changes. For example, common genetic variants in DNMT genes appear to moderate the effects of daily life stressors and perinatal adversity on paranoid ideation. 60 DNMTs are enzymes required for the methylation of neurogenic genes, also suggesting that exposure to chronic stressors early in life might disturb brain function in adulthood. 61 Large methylation changes at the beginning of the lifespan, particularly on genes implicated in developing severe psychiatric conditions (e.g. DLG4, DRD2, NOS1, NRXN1, and SOX10) may also indicate vulnerability to the effects of stress and psychiatric disorders by epigenetic mechanisms. 59
Prenatal Adversity
Summary of studies examining epigenetic modifications in response to early life adversity during (a) prenatal time points (b) childhood time points or (c) in adults with a history of early life adversity.
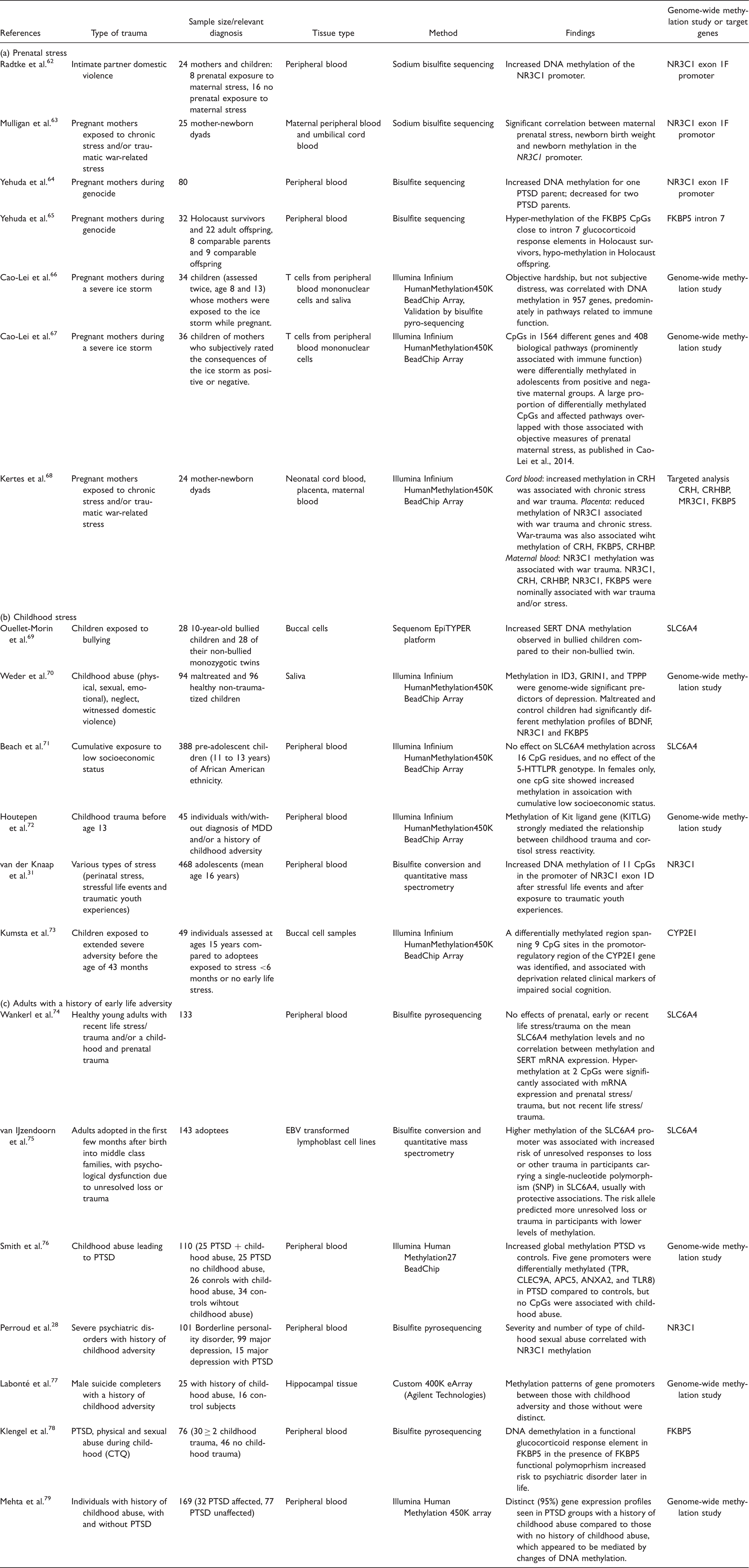
Note. PTSD: posttraumatic stress disorder; EBV: Epstein–Barr virus; MDD: major depressive disorder; SERT: serotonin transporter; CTQ: childhood trauma questionnaire.
Considering the unequivocal role of the GR in stress responsivity, several studies have focused on epigenetic modifications of NR3C1 in response to various types of prenatal stress. For example, in those exposed to intimate partner violence during the prenatal period, DNA methylation was increased in the NR3C1 1F promoter region. 62 Additionally, in offspring of mothers pregnant during the Holocaust genocide, hyper-methylation of the NR3C1 exon 1F promoter methylation was observed much later in life, specifically in offspring who had one parent suffering from PTSD. 64 Methylation was, however, decreased for offspring with two parents who had PTSD, presenting what could be a complex mechanism of potential maternal and paternal effects of PTSD on epigenetic regulation of NR3C1, although it is possible that epigenetic changes occurring in offspring were an effect of living with a parent with PTSD rather than genocide exposure. Together with evidence from animal studies, 81 hyper-methylation of the NR3C1 promoter appears to be a robust epigenetic modification occurring in response to stress early in life. Altogether, there appear to be strong and widespread effects of prenatal stress on the epigenetic regulation of the stress signaling system that lasts into adulthood, although the intergenerational mechanisms for some genes (e.g. FKBP5 and CRH) need to be further explored perhaps using animal models.
Childhood Adversity Assessed During Childhood and Adolescence
Considering that certain epigenetic markers occurring during development may set an individual on a path for developing psychopathology later in life, some studies have aimed to examine methylation patterns in the brains of those exposed to chronic stress during childhood (Table 1(b)). Some of the effects seen with prenatal stress are also seen with stress during childhood. In suicide completers exposed to childhood abuse, hyper-methylated sites were observed in the NR3C1 variant 1 promoter region at a non-canonical binding site for nerve growth factor–induced protein A. This was further associated with down-regulated expression of the NR3C1 1F splice variant in the hippocampus. 82 While these localized findings for epigenetic regulation of the GR are quite interesting, the effects of childhood adversity is likely epigenome-wide, as methylation profiles in the hippocampus of suicide completers with a history of childhood abuse were reportedly bidirectional and dynamic, with both methylation and de-methylation of numerous genes observed. 77 Hyper-methylation at promoters of genes that encode ribosomal RNAs were also reported in abused suicide victims compared to controls, possibly indicating widespread effects as ribosomal RNA methylation and transcription directly affect cellular function and synaptic plasticity associated with fear memory and cognitive dysfunction. 83
Additional studies have examined epigenome-wide methylation differences in children exposed to childhood maltreatment or trauma (e.g. physical abuse, sexual abuse, neglect, deprivation, emotional abuse, witnessed domestic violence) compared to non-traumatized individuals.72,70,73 Weder et al. 70 reported that methylation patterns at the genes ID3, GRIN1, and TPPP were genome-wide significant predictors of depression later in life. Maltreated children relative to control children had significantly altered methylation profiles at the BDNF, NR3C1, and FKBP5 genes, which are all genes that respond to stress exposure and implicated in psychopathology. In a sample of English and Romanian adoptees, individuals chronically exposed to severe deprivation before 43 months of age showed hyper-methylation across nine CpGs spanning the promoter of the CYP2E1 gene at the age of 15 years. Hyper-methylation of CYP2E1 was also associated with impaired social cognition, a clinical marker of deprivation. 73 In addition, Houtepen et al. 72 reported that methylation of the KIT ligand gene KITLG, which is involved in haematopoiesis and the cortisol response, strongly mediated the relationship between childhood trauma and cortisol stress responsivity. This would suggest a link between stress occurring early in life and epigenetic modifications lasting into adulthood that may increase risk to psychopathology in adulthood by altering the responsivity of the stress hormone system.
Another group of studies suggest epigenetic changes in peripheral tissue at the SLC6A4 locus and the consequences of stress in older children. 74 For example, in a study by Beach et al., 71 children aged 11–13 years with cumulative exposure to low socio-economic status showed hyper-methylation at the SLC6A4 locus. A similar effect was observed in a second study, with increased methylation of SLC6A4 in bullied twins at age 10 compared to their non-bullied monozygotic twins. 69
Effects of Childhood Adversity Lasting Into Adulthood
The studies assessing epigenetic modifications that occur in response to prenatal and childhood adversity strongly support evidence that stress during development leads to epigenetic modifications that likely influence gene expression later in life, 84 especially in genes involved in stress responsivity. During these critical developmental time points, changes in expression of “stress genes” might alter an individual’s ability to cope with stress and set the stage for vulnerability to psychopathology later in life. Some groups have assessed clinical populations of individuals who were exposed to childhood adversity and who developed PTSD in adulthood (Table 1(c)). These investigations collectively support that childhood adversity is associated with specific epigenetic modifications in those that develop a psychiatric disorder.
Other groups assessing childhood abuse in adults with severe psychiatric disorders have focused on stress candidate genes. In one report, the severity and number of exposures to childhood sexual abuse was positively correlated with NR3C1 hyper-methylation in subjects with severe psychiatric disorders including PTSD and comorbid major depressive disorder. 28 In another cohort of subjects with PTSD exposed to childhood adversity, DNA de-methylation was found to be decreased in a glucocorticoid response element in FKBP5, specifically in the presence of an FKBP5 functional polymorphism that increased risk to psychiatric disorder later in life. 78 These data mirror the findings from prenatal adversity studies, supporting the inference that such epigenetic modifications to the stress system occur in response to early life adversity and may last into adulthood to increase development of a severe psychiatric condition.
Stress and Trauma During Adulthood
In addition to early life adversity, experiencing extreme levels of stress or trauma during adulthood can also increase the risk of developing severe psychiatric conditions. However, the mechanism is likely distinct between these two scenarios. Firstly, early life experiences occur during a particularly sensitive time, where environmental influences can have strong effects on the brain and body, causing molecular, structural, and behavioral disturbances that stay with the individual into adulthood. While severe stress or trauma occurring exclusively in adulthood can also result in detrimental biological consequences, body systems are already matured thus possibly causing a different cascade of events. In fact, we could show that patients suffering from PTSD with either only adult trauma or a combination of child abuse and adult trauma had distinct gene expression and DNA methylation signatures in peripheral blood. 79
Epigenetic Changes in Preclinical Stress Models
Adult animals exposed to severe acute or non-traumatic chronic stress have shown various epigenetic changes attributable to stress exposure. Considering a hallmark of PTSD is the formation of fear memories, many studies have used fear learning paradigms for exploring the consequences of trauma exposure. 85 These studies have shown that fear conditioning induces the expression of DNA methyl-transferases type 3A and 3B (Dnmt3A/3B), leading to altered DNA methylation at genes that are important for memory formation and plasticity (e.g. Reelin, Pp1, and Bdnf).20,86–88Others have highlighted a role for histone acetylation in long-term fear recall and spatial memory. For example, environmental enrichment was shown to improve memory deficits in the brain; this was associated with up-regulation of histone acetylation and activity of histone deacetylases, which modulate synaptic plasticity via altering the expression of genes including glutamate receptor subunits and Bdnf.89–91 These findings have linked stress to brain plasticity and behavior, by long-lasting epigenetic modifications.
Clear epigenetic changes have also been observed in animal models of chronic stress. 92 One of the primary findings from genome-wide epigenetic studies in rodent models is that chronic stress induces widespread alterations in histone modifications, particularly in the hippocampus. 93 For example, chronic variable stress caused alterations to the expression of transcripts that encode histones or recruit histone writers and erasers.94,95 With regard to methylation, associations between chronic defeat stress and Dnmt3A expression was shown in the nucleus accumbens, which is a brain region that plays a significant role in processing of aversion, motivation, and rewards. 96 Interventions (mainly pharmacological such as antidepressants), which induce H3K9 di- or tri- methylation, have been shown to promote resilience to stress and depressive-like behavior.97,98 Chronic stress in adult animals also exerts epigenetic modifications to key genes involved in HPA-axis signaling, 99 such as Crh,100,101 Hsp90, 102 and Bdnf.103,104
Epigenetic changes in response to severe and chronic stress can clearly be associated with stress exposure in animal models. While this is more complex in human studies, clinical studies complement these findings, supporting the existence of lasting epigenetic effects in response to trauma or prolonged stress within both the central and peripheral nervous systems. In the following sections, we will discuss examples of epigenetic changes observed in humans exposed to acute traumatic events or non-traumatic chronic stress in adulthood.
Epigenetic Changes Due to Acute Traumatic Events in Adults
Summary of studies examining epigenetic changes in response to trauma in adulthood, with a focus on genome-wide methylation studies and candidate gene studies in PTSD.

Genome-wide methylation studies support the existence of epigenetic differences between trauma-exposed individuals with PTSD compared to psychiatrically healthy controls, with cross-sectional differences in DNA methylation observed in cohorts from Atlanta 76 and Detroit. 105 These studies have shown that epigenetic differences were able to differentiate those who have developed PTSD compared to those who did not. Later studies from longitudinal cohorts of US military personnel exposed to combat-trauma further suggested significant differences in global methylation in PTSD patients relative to controls,109,112 particularly in genes involved in immune105,112 and nervous system function. 113 Such longitudinal studies allow the dissection of separate contributions of trauma exposure versus disease development, which is not readily possible in cross-sectional studies.
A larger number of studies have investigated DNA methylation of specific candidate genes, including genes involved in the regulation of the stress hormone axis. Here, studies have focused on NR3C1 (exon 1F) and FKBP5. In one study from saliva, NR3C1 methylation at CpG site 42 was increased in men with PTSD; 115 however, in another study from blood, methylation at CpG site 28 was reduced in PTSD. 29 The differences may represent tissue- or CpG site-specific differences, variability in the trauma type or its timing, or sex and age. 122 In an earlier study, veterans exposed to severe war trauma underwent prolonged exposure therapy and were assessed pre- and post-exposure treatment. 114 The study showed that methylation of NR3C1 exon 1F locus pre-treatment predicted treatment outcome, but methylation was not significantly different in responders or non-responders to the exposure therapy at post-treatment or follow-up. 114 Methylation of the FKBP5 exon 1 promoter also did not predict treatment response, but was decreased in association with recovery. 114 This study provides preliminary evidence that these methylation markers may underlie the severity of PTSD symptoms, and that reversal of the methylation changes may tract with effective therapeutic intervention.
The immune system is one of the main effectors of HPA axis activity, and in turn, changes in methylation are associated with total life stress, PTSD diagnosis, and a history of childhood abuse. 76 Epigenetic effects to genes involved in immune function are a consistent finding in PTSD, supporting that sustained sympathetic nervous system activity in response to trauma stimulates immune system activation. For example, DNA methylation of genes associated with immune function was found to be largely negatively correlated with traumatic burden in PTSD. 105 Others consistently reported altered methylation of pro-inflammatory cytokines, such as interleukin 12 and 18, in individuals with PTSD compared to those without.76,112,118 Analysis of histone and DNA methylation markers suggest that widespread epigenetic changes in PTSD, including those in immune genes, might occur by alterations to DNMT1 118 or miRNAs, 119 which are molecules that induce and maintain global methylation patterns.
In addition to identification of epigenetic markers associated with diagnostic outcomes, some studies have begun to additionally assess associations of DNA methylation to endophenotypes of psychiatric disease, including neuroimaging phenotypes. For example, a genome-wide significant variant, rs717947 (positioned in an inter-genic locus at chromosome 4p15), was identified as a methylation quantitative trait locus associated with altered cortical activation in response to fearful faces. 123 Another study showed associations between the methylation status, a polymorphic site in the 3’UTR of the SKA2 gene—involved in mitosis—with reduced thickness of several cortical areas and symptom severity in PTSD. 121 Finally, methylation in peripheral blood samples of ADCYAP1R1, a gene involved in regulating the cellular stress response, was associated with PTSD diagnosis and symptom severity, specifically in females. 106 Collectively, these studies support that epigenetic changes in response to stress may “mediate” particular stress phenotypes.
Several studies have now investigated DNA methylation in the context of G×E including candidate genes involved in neurotransmitter turn-over such as SLC6A4 (encoding the serotonin transporter SERT), SLC6A3 (encoding the dopamine transporter), and COMT (encoding enzymes that degrade neurotransmitters such as dopamine). In one study, reduced SLC6A4 methylation levels were associated with more traumatic events and increased risk for PTSD, only in those carrying a specific SLC6A4 risk allele genotype, while higher methylation appeared protective against the development of PTSD. 108 In an independent cohort, individuals with PTSD carrying the SLC6A3 9-repeat allele were at higher risk to PTSD when also having higher methylation in the SLC6A3 promoter locus. 110 Finally, increased COMT promoter methylation was associated with impaired fear inhibition in individuals with PTSD carrying the COMT met/met genotype; this genotype has been extensively associated with a range of psychiatric conditions and phenotypes. 124 These studies suggest that epigenetic changes following trauma often only occur in the context of specific genetic variation and that genetic risk may only manifest with concomitant epigenetic changes.
Epigenetic Changes Due to Sustained Non-traumatic Stress in Adults
Summary of studies examining epigenetic changes in response to non-traumatic chronic stress exposure in adulthood.

For example, in workers employed in the manufacturing industry in environments of high or low stress, Myaki et al. 126 examined DNA methylation of the tyrosine hydroxylase (TH) gene. TH encodes an enzyme involved in synthesis of catecholamines including serotonin, dopamine, epinephrine, and norepinephrine, which are all altered in rodent models of chronic stress.128,129 Methylation of CpG sites in the TH gene and the flanking 5′ region was significantly higher in individuals exposed to high occupational stress compared to individuals working in low-stress environments. 126 The same group also explored altered methylation in the BDNF gene based on a priori hypothesis that this gene is also epigenetically affected in a preclinical model of PTSD. 103 Slightly increased methylation was observed across the whole gene in subjects exposed to high-stress compared to low-stress environments, although the groups showed no difference in promoter-specific methylation. 125 Work-related stress in this study was assessed using a self-administered questionnaire, with methylation levels of groups from the lowest and highest job-strain quartiles compared. 125
Another group focused on methylation of the SLC6A4 gene due to some evidence that methylation differences in this gene occur in stress-induced psychopathologies such as PTSD (discussed in section “Epigenetic changes due to acute traumatic events in adults”) and depression. 130 In shift-working nurses in high-stress work environments compared to those in low-stress work environments, five CpG sites in the promoter region of SLC6A4 were examined. 30 Higher methylation across the tested SLC6A4 CpGs was found in those working in a high- compared to low-stress environment, and this was associated with burnout and the perception of stress. 30 Further investigation into the effects of non-traumatic chronic stressors, such as work-related stress or caregiving, is limited. This is likely as the length and severity of non-traumatic stress is often not as well defined as traumatic stress, and many confounding factors make the results of these studies less clear. These studies might also be confounded by the number of stressors, which might have an additive effect, and further studies are needed to disentangle this.
Future Directions and Conclusions
Studies examining epigenetic changes in response to stress are providing a potential opportunity to identify vulnerable and resilient populations (e.g. based on combined prior exposure, other environmental factors and genetics) and biomarkers of risk for those exposed to chronic stress. However, additional studies of how different types and lengths of stress lead to epigenetic changes are required as well as of combined effects of positive and negative environments. The field of epigenetics is in fact relatively young, and studies examining epigenetic modifications in conditions of chronic stress are limited. One example is that, to the best of our knowledge, no studies have examined epigenetic modifications in postmortem brain samples derived from individuals with PTSD. Additionally, human studies are also limited in that there are few longitudinal studies with repetitive measurements from the same individual at different time points relative to the stress exposure, and currently, many studies published are cross-sectional and underpowered. Changes to gene expression by epigenetic modifications are prominent gene and environment interactions that are an important aspect of future chronic stress and trauma research. Particularly, epigenetic patterns in peripheral tissues may act as biomarkers that may serve as sentinels of when genetic predisposition followed by environmental exposure leads to trajectories of risk to develop psychiatric symptoms and syndromes. For mechanistic understanding, tissue specificity of these epigenetic changes must be addressed and understood. Overall, more research in this area will lead to a better understanding of stress-induced epigenetic effects and mechanisms as well as their potential moderation, allowing for their therapeutic exploitation and the development of new interventions and treatments.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by an ERC starting grant (under the FP7 frame work, #281338) to EBB and an NHMRC grant (ECF #1105445) held by NM. The work was also supported by an IBRO Fellowship to NM and Alexander von Humboldt Fellowships to NM and CC.