
Abstract
Telomere length is a biomarker of cell aging that is hypothesized to contribute to women’s greater longevity. Although most previous studies have found no sex difference in telomere length at birth, it is well established that females have longer average telomere length than males during adulthood. Proposed biological mechanisms underlying sex differences in adult telomere length include differences in sex steroid hormones and body size, which emerge during the pubertal transition. The purpose of this study was to examine the total effect of sex on telomere length during early adolescence and to examine estradiol, total testosterone, and body surface area (BSA; a measure of body size) as potential mediators of sex differences in telomere length. Data were from a population-based sample of 126 female and 109 male Hispanic adolescents aged 8 to 14 years from the Early Life Exposures in Mexico to ENvironmental Toxicants (ELEMENT) study. Relative telomere length (T/S ratio) was measured by the quantitative polymerase chain reaction method; sex steroid hormones were measured using an automated chemiluminescent immunoassay, and BSA was calculated using measured height and weight. Adjusting for age and pubertal status, we found that girls had significantly longer telomeres than boys (β = .13; P < .01), but there were no significant indirect effects of sex on telomere length through any of the proposed mediators. We conclude that sex differences in telomere length are evident during early adolescence but are not explained by cross-sectional differences in sex steroid hormones or body size.
It has been hypothesized that sex differences in telomere length, a biomarker of cell aging, may help explain why women live longer, on average, than men. 1 -3 While there is little evidence of a difference in telomere length between the sexes at birth 1,4 -7 (for an exception, see 8 ), it is well established that telomeres are longer in adult females than in adult males. 2,9 -13 Due to a dearth of studies in children and adolescents, it is currently unknown when sex differences in telomere length appear in humans and what the mechanisms are by which these differences occur. 14 Understanding when and how sex differences in telomere length emerge may provide insight into why women have longer average life expectancy than men. 1,3,15 In this study, we used data from a population-based sample of Hispanic children aged 8 to 14 years to examine sex differences in leukocyte telomere length during the pubertal transition and to explore potential biological explanations for sex differences in telomere length during this stage of development, including differences in sex steroid hormones and body size.
The Telomere Hypothesis of Aging
Telomeres are protective caps of repeated nucleotide sequence at the ends of eukaryotic chromosomes that promote chromosomal stability. 16 DNA replication is semiconservative and requires that the newly synthesized strand be constructed in the 5′ to 3′ direction. 17 Due to this directional requirement, replication of the lagging DNA strand requires RNA primers of 8 to 11 nucleotides, from which DNA replication machinery may begin synthesis of the new DNA strand. This helps create the end-replication problem, whereby a varying amount of single-stranded DNA remains following replication due to RNA primer location, RNA primer length, and other processing factors. 17 This single-stranded DNA is subsequently removed by repair mechanisms, meaning that telomeres naturally shorten with mitosis. 18 Oxidative damage and DNA replication stress also contribute to telomere loss. 16,19 When telomeres become critically shortened, cellular senescence is triggered and cells lose their ability to divide. 20,21
This end stage of cellular senescence plays a role in the pathophysiology of biological aging and is hypothesized to contribute to the development of the aging phenotype and age-related diseases. 22,23 A number of epidemiologic studies have shown that leukocyte telomere length is associated with adult morbidity 24 -27 and mortality 28 -30 independent of chronological age. Furthermore, while leukocyte telomere length has traditionally been viewed as a noncausal biomarker of aging, growing evidence suggests that telomere shortening may be a causal determinant of age-related disease and longevity. 31,32
Sex Differences in Telomere Length
Several hypotheses have been proposed to explain why adult women tend to have longer telomeres than adult men. First, the sex steroid hormone estrogen, which is found at higher levels in women than men, is thought to activate telomerase, an enzyme that lengthens telomeres, most notably in cancer cells. 2,33 Additionally, it is known that reactive oxygen species lead to increased oxidative stress and damage telomeres, 1,11,34,35 and estrogen is thought to possess antioxidant properties that may protect telomeres from extensive oxidative damage. 1,36 Second, women may have longer telomeres not due to the protective effects of estrogen but rather due to lower levels of exposure to testosterone, which is found at higher levels in men. Testosterone lacks antioxidant properties and may increase the deleterious effects of oxidative stress on telomeres. 2,37 Finally, the sexual size dimorphism hypothesis proposes that larger body size results in more cell divisions, and therefore, shorter telomeres. 2,3 As human males typically have larger bodies than human females, this hypothesis suggests that males would naturally have shorter telomeres.
Puberty and Telomere Length
The hormonal and body size hypotheses discussed earlier suggest that sex differences in telomere length may emerge during the pubertal transition, which is characterized by a series of hormonal changes that lead to sexual maturation, reproductive capability, and rapid changes in body size, shape, and composition. 38,39 On average, females enter puberty earlier than males, but the tempo of puberty varies greatly between individuals. 39 The first hormonal changes typically occur between the ages of 5 and 6 years when the production of adrenal androgens begins to increase. 38 Around the age of 9 or 10 years in girls and slightly later in boys, an increase in pituitary gonadotropins results in the growth of ovaries and testes as well as an increase in the production of estradiol and testosterone. 40 Puberty is normally completed between 2 to 4 years following this stage of development, which is known as gonadarche. 38 About 25% of human growth in height occurs during the pubertal transition, resulting in an average sexual size dimorphism of 13 cm between adult males and females. 39,41
Hypotheses
Given that changes in sex steroid hormones begin as early as age 9, we hypothesized that girls would have longer telomere length than boys in this sample of 8- to 14-year-olds. Next, we hypothesized that girls would have higher levels of estradiol, lower levels of testosterone, and smaller body surface area (BSA; a measure of body size) than boys and that that estradiol would be positively associated with telomere length, while testosterone and BSA would be inversely associated with telomere length. Finally, we hypothesized that sex differences in estradiol, testosterone, and body size would partially mediate or explain sex differences in telomere length.
Materials and Methods
Study Participants
This study used data from the Early Life Exposures in Mexico to ENvironmental Toxicants (ELEMENT) study, which was designed to investigate how environmental exposures affect the health of pregnant women and their children. Participants were recruited from maternity hospitals in Mexico City and have been followed for over 2 decades. Detailed study information can be found in Gonzalez-Cossio et al 42 and Tellez-Rojo et al 43 . During a follow-up visit in 2010, study personnel collected urine and blood samples from 249 Hispanic children aged 8 to 14 years. Additional data were collected via clinical examinations and questionnaires administered to caregivers. A total of 13 participants were excluded from this analysis due to missing data on telomere length, estradiol levels, total testosterone levels, or physician-assessed Tanner stage. One additional participant with an outlying average Tanner score of 5 was excluded, resulting in a final analytic sample size of 109 boys and 126 girls.
Measures
Telomere length
Blood was collected in PaxGene tubes and stored frozen (−70°C or lower) until DNA was isolated. The PaxGene Blood DNA kit (PreAnalytix Feldbachstrasse, Switzerland) was used to extract DNA from blood leukocytes. Nanodrop was used for DNA quantification. The telomere length assay was performed in the laboratory of Dr Elizabeth Blackburn at the University of California, San Francisco, using the quantitative polymerase chain reaction method to measure telomere length relative to standard reference DNA (T/S ratio) as described in detail elsewhere. 44 Each sample was assayed at least twice. The T/S ratios that fell into the 7% variability range were accepted, and the average of the 2 was taken as the final value. A third assay was run for samples with >7% variability, and the average of the 2 closest T/S values was used. The average interassay coefficient of variation was 2.2%.
Sex
Sex was coded 0 for male and 1 for female and was based on sex at birth.
Sex hormones
Sex steroid hormone levels were measured in serum that was stored in cryogenic tubes at −70°C on the day of collection in Mexico prior to shipping to the University of Michigan where they were stored at −80°C. Levels of total testosterone and estradiol were measured by the Clinical Ligand Assay Service Satellite Laboratory (University of Michigan School of Public Health, Ann Arbor, Michigan 45 ). The levels of each hormone were measured using an automated chemiluminescent immunoassay (Bayer Diagnostics Newbury, UK, ACS 46 :180.
Body size
Body surface area was measured in m2. We employed Mostellers
47
formula for the calculation of BSA as shown below:
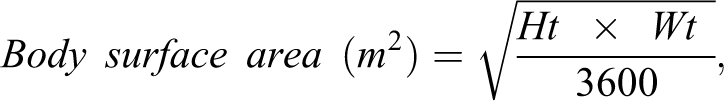
where Ht is height in centimeters and Wt is weight in kilograms. This simplified formula has been validated for use in children 48 and was recommended following a comparison study of the various BSA calculation techniques, including early 3D scanning methods. 49
Covariates
Age and pubertal development were considered potential confounders and were therefore included as covariates in all models. Age was measured in years. Pubertal development was measured using physician-assessed Tanner staging, ranging from 1 (no development [preteen]) to 5 (mature stage [adult]). 50 -52 Consistent with prior research, 53 we used the average Tanner score for physician-observed pubic hair development and breast development to measure pubertal development in girls and the average Tanner score for physician-observed pubic hair development and genital development to measure pubertal development in boys.
Analytical Design
Our first hypothesis was that girls would have longer telomere length than boys. We regressed telomere length on sex, controlling for age and average Tanner score, to generate an estimate of the total effect (c) of sex on telomere length (Figure 1, Panel A). Next, we hypothesized that girls would have higher levels of estradiol, lower levels of testosterone, and smaller body size than boys and that that estradiol would be positively associated with telomere length, while testosterone and body size would be inversely associated with telomere length. We regressed each hypothesized mediator on sex and the covariates to determine whether there were significant sex differences in estradiol, testosterone, and/or BSA. Finally, we hypothesized that sex differences in estradiol, testosterone, and BSA would partially mediate, or explain, sex differences in telomere length. We regressed telomere length on sex, the mediators, and the covariates (Figure 1, Panel B) to obtain estimates of the direct and indirect effects of sex on telomere length. The indirect effect of sex on telomere length through each of the mediators was calculated by multiplying the path coefficient from sex to the mediator by the path coefficient from the mediator to telomere length (a1-3b1-3). The direct effect (c’) was calculated as the difference between the total effect (c) and the sum of the total indirect effects ([a1b1] + [a2b2] + [a3b3]). This approach assumed no residual confounding of the mediator–outcome relation and no interaction between the mediators and sex. 54,55 We used a directed acyclic graph to select the appropriate covariates for the models, 56 and we examined interactions between sex and each of the potential mediators (no significant interactions were found; results not shown). We used the PROCESS macro for SAS designed by Hayes 57 to test the hypotheses. Bias-corrected bootstrap confidence intervals (CIs) were used to test significance of indirect effects. 58 The indirect effect is significant at the P < .05 level if the 95% CI does not contain 0.
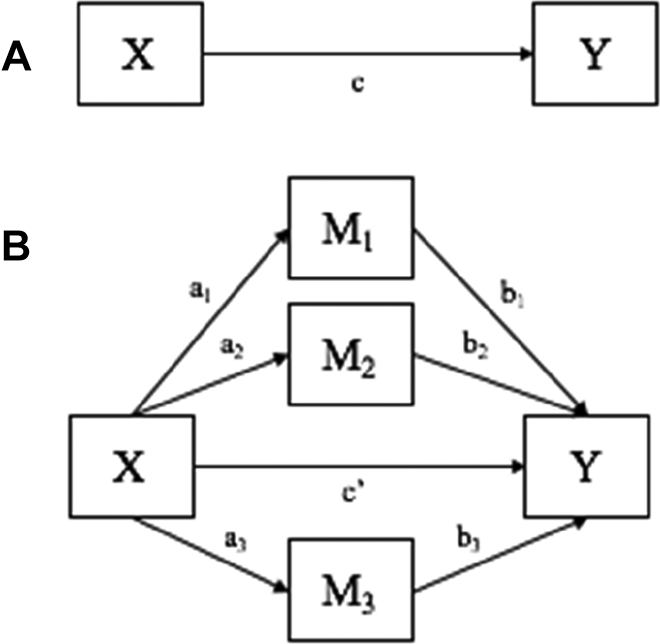
Conceptual models. Panel A shows the total effect model. Panel B shows the mediation model, where X is sex; Y is telomere length; M1-M3 are estradiol, total testosterone, BSA, respectively; c is the total effect of X on Y; c′ is the direct effect of X on Y; a1b1 is the indirect effect of X on Y through M1; a2b2 is the indirect effect of X on Y through M2; a3b3 is the indirect effect of X on Y through M3.
Results
Descriptive statistics for the full analytic sample and by sex are shown in Table 1. Median telomere length (T/S ratio) was significantly longer for girls (1.48, interquartile range (IQR) = 0.28) than for boys (1.38, IQR = 0.33; P < .01), and girls had higher median levels of estradiol (22.50 pg/mL, IQR = 25.80) than boys (16.60 pg/mL, IQR = 6.60; P < .01). There were no significant sex differences in age (girls: 9.90, IQR = 2.20; boys: 9.90, IQR = 2.30; P = .35), average Tanner score (boys and girls: 1.00, IQR = 0.50; P = .25), testosterone (girls: 19.30 ng/dL, IQR: 15.70; boys: 20.80 ng/dL, IQR = 40.50; P = .14), or BSA (girls: 1.18, IQR: 0.26; boys: 1.16, IQR: 0.21; P = .98).
Descriptive Statistics for all Study Variables.a
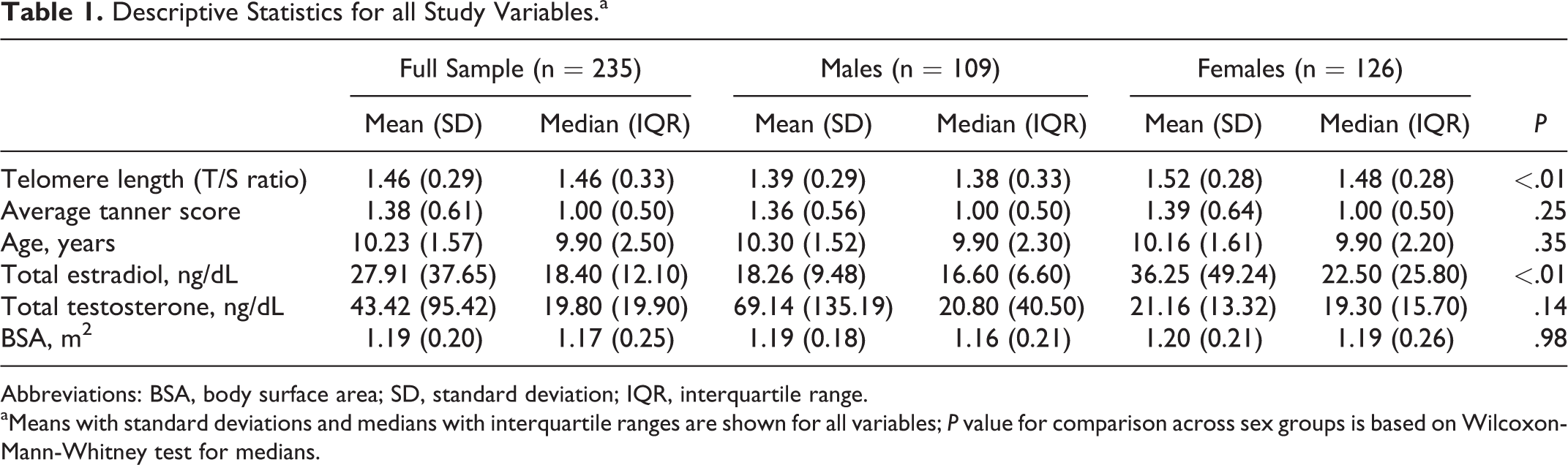
Abbreviations: BSA, body surface area; SD, standard deviation; IQR, interquartile range.
aMeans with standard deviations and medians with interquartile ranges are shown for all variables; P value for comparison across sex groups is based on Wilcoxon-Mann-Whitney test for medians.
As shown in Table 2, there was a significant positive total effect (c) of female sex on telomere length (β = 0.13, P < .01), controlling for age and average Tanner score. In fully adjusted models, there was a significant positive effect of female sex on estradiol (a1 ; β = 0.46, P < .01); however, there was no significant effect of female sex on total testosterone (a2 , β = −0.34, P = .10), and there was no significant effect of female sex on BSA (a3 , β = 0.01, P = .57). Neither estradiol (b1 , β = 0.02, P = .56), total testosterone (b2 , β = 0.02, P = .08), nor BSA (b3 , β = −0.20, P = .14) were significantly associated with telomere length. After accounting for the hypothesized mediators, there was a significant, positive direct effect (c′) of female sex on telomere length (β = 0.12, P < .01). There were no significant indirect effects of sex on telomere length through estradiol (a1b1 , β = 0.01, 95% CI: −0.02 to 0.04), total testosterone (a2b2 , β = −0.01, 95% CI: −0.03 to 0.00), or BSA (a3b3, β = −0.00, 95% CI: −0.01 to 0.00). The total indirect effect of sex on telomere length through all the mediators was not statistically significant ([a1b1] + [a2b2] + [a3b3]), β = 0.00, 95% CI: −0.03 to 0.03).
Regression Results for the Mediation of the Effect of Female Sex on Telomere Length (T/S ratio) by Estradiol (ng/mL), Total Testosterone (ng/dL), and BSA (m2), n = 235.a
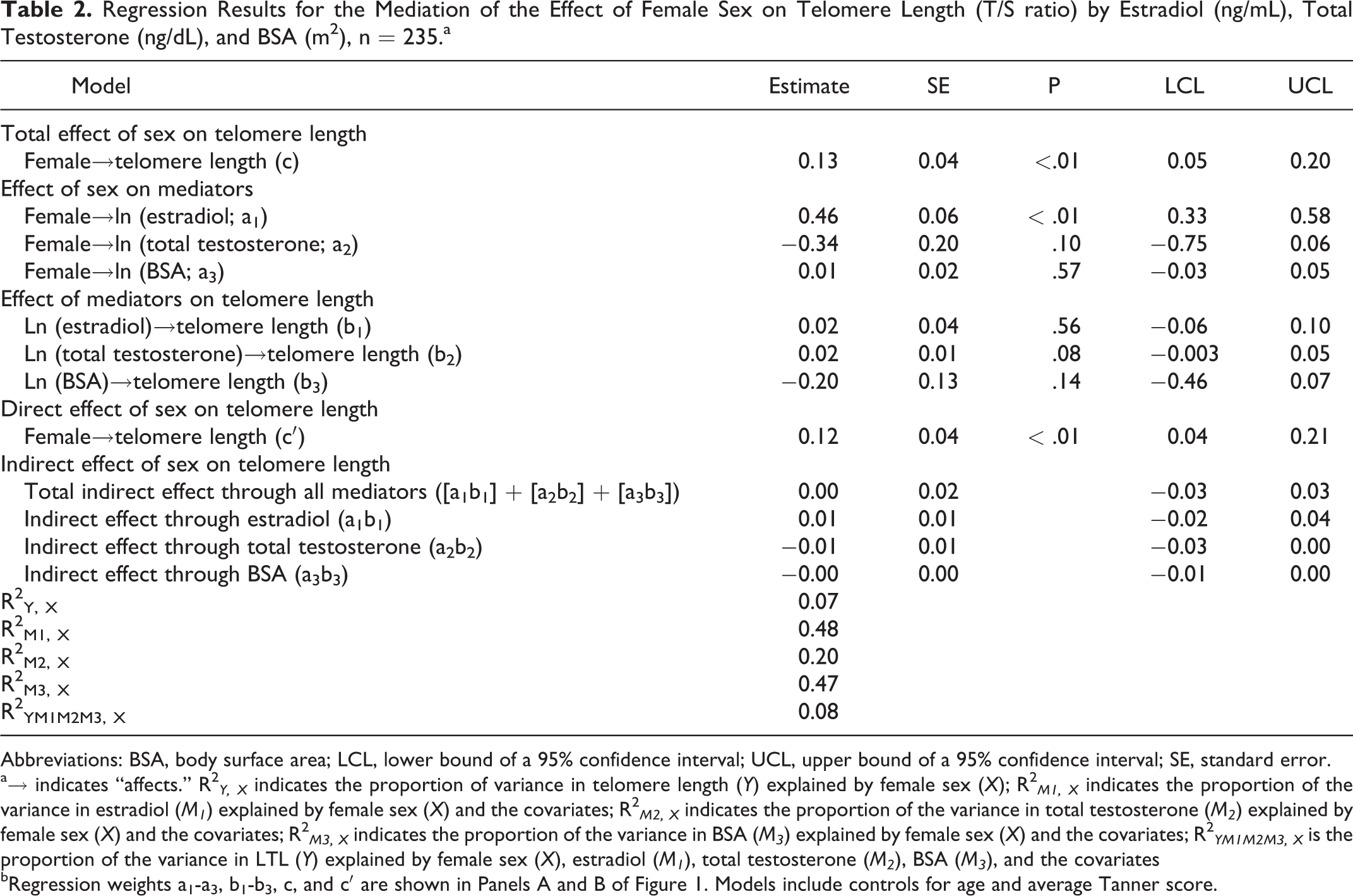
Abbreviations: BSA, body surface area; LCL, lower bound of a 95% confidence interval; UCL, upper bound of a 95% confidence interval; SE, standard error.
a→ indicates “affects.” R2 Y, X indicates the proportion of variance in telomere length (Y) explained by female sex (X); R2 M1, X indicates the proportion of the variance in estradiol (M1 ) explained by female sex (X) and the covariates; R2 M2, X indicates the proportion of the variance in total testosterone (M2 ) explained by female sex (X) and the covariates; R2 M3, X indicates the proportion of the variance in BSA (M3 ) explained by female sex (X) and the covariates; R2 YM1M2M3, X is the proportion of the variance in LTL (Y) explained by female sex (X), estradiol (M1 ), total testosterone (M2 ), BSA (M3 ), and the covariates
bRegression weights a1-a3, b1-b3, c, and c′ are shown in Panels A and B of Figure 1. Models include controls for age and average Tanner score.
Female sex and the covariates explained 7% (R2 Y, X ) of the variance in telomere length, while female sex and the covariates explained 48% of the variance in estradiol (R2 M1, X ), 20% of the variance in testosterone (R2 M2, X ), and 47% of the variance in BSA (R2 M3, X ). Taken together, female sex, estradiol, total testosterone, BSA, and the covariates explained 8% (R2 YM1M2M3, X; Table 2) of the variance in telomere length.
Discussion
In this study, we investigated sex differences in telomere length in a population-based sample of children aged 8 to 14 years. While most previous studies have found no sex difference in telomere length at birth, 1,4 -7 a large body of evidence suggests that adult females have significantly longer telomeres than adult males. 2,9 -13 Based on these findings, explanations for the sex difference in adult telomere length have largely focused on sex differences in biological factors, such as sex steroid hormones, that emerge at some point during sexual maturation. Studies in children and adolescents may help us understand when and why sex differences in telomere length first emerge and could provide important insight into the biological mechanisms underlying differing life expectancies between males and females.
Consistent with expectations, we found that girls had longer telomeres than boys. However, we did not find support for the hypothesis that sex differences in telomere length are explained by male/female differences in estradiol, testosterone, or BSA. We expected to find that female sex would be related to increased estradiol, decreased testosterone, and decreased BSA, which would in turn be related to longer telomere length. While girls had significantly higher levels of estradiol than boys, we found that estradiol did not have a significant effect on telomere length. On average, girls had lower levels of testosterone than boys, but the sex difference was not statistically significant, and there was no association between testosterone and telomere length. Finally, there was no sex difference in BSA, and BSA was not associated with telomere length. In this small study of early adolescents, we failed to find support for 2 of the leading biological explanations that have been proposed to account for sex differences in telomere length. 3,59,60
Limitations, Strengths, and Directions for Future Research
Limitations of this study include small sample size, minimal variability in pubertal development, the use of an aggregate measure of pubertal onset, the cross-sectional design, and a racially/ethnically homogenous sample. First, due to small sample size, this study may have been underpowered to detect indirect effects of sex on telomere length through the hypothesized biological mediators. The failure to detect significant associations of telomere length with sex hormones and BSA may be due to a lack of statistical power. Larger studies are needed to confirm or refute the findings presented here. Next, because most respondents were prepubertal (median average Tanner score was 1, which indicates no pubertal development), we were unable to determine whether sex differences in telomere length were larger at later stages of pubertal development. The use of an aggregate measure of pubertal onset is also a limitation, and future studies should examine individual milestones for which there is greater intra- and interindividual variability. Longitudinal studies starting at birth would allow us to determine when the sex difference in telomere length emerges and would enable us to determine whether within-person changes in sex hormones and BSA are associated with within-person change in telomere length. A longitudinal design would facilitate a comparison of sex differences in the rate of telomere attrition during the pubertal transition and would provide stronger evidence for a causal relationship between the hypothesized biological mediators and telomere length. Additional DNA samples were collected for 223 of our study participants during a second peripubertal examination, which was completed 3 to 5 years later (mean [standard deviation (SD)] age was 13.5 [1.8] years for girls and 13.7 [1.8] years for boys at the second examination). These stored samples could be used to obtain repeat measures of telomere length for future research on telomere attrition during the pubertal transition. Finally, future studies should consider whether the findings reported here apply to other racial/ethnic groups and to individuals in countries other than Mexico.
Balancing these limitations are several key strengths, including the focus on early adolescence, the use of a population-based sample, and the availability of state of the art measures of sex steroid hormones, body size, and telomere length. While sexual maturation is not complete until mid to late adolescence, the hormonal and physical changes in puberty begin during early adolescence, 38 making this an ideal developmental stage in which to study the emergence of sex differences in telomere length as well as the hypothesis that male/female differences in sex steroid hormones and body size contribute to sex differences in telomere length. Furthermore, the use of a population-based sample, rather than a clinical sample, enhances generalizability of study results to the entire population of early adolescents. Finally, the availability of sex steroid hormone data, measured weight and height, and data on telomere length in a sample of both girls and boys enabled us to examine 2 of the leading hypotheses for sex differences in telomere length.
An important direction for future research is the examination of alternative biological mechanisms that could explain sex differences in telomere length. In addition to sex hormones and body size, it has been proposed that chromosomal differences may contribute to sex differences in telomere length. It has been suggested that because some telomere maintenance genes are found on the X chromosome, males naturally have a disadvantage because they only have 1 X chromosome and therefore are more susceptible to mutation, malfunction, or damage as they do not have backup copies of these genes. 61 -63 However, females “turnoff” no less than ∼75% of the genes located on their “extra” X chromosome through X inactivation. 64 It has also been proposed that females benefit over time because X inactivation favors the preservation of the chromosome with longer telomeres, whereas men only have 1 X chromosome and cannot benefit from positive selection of longer telomeres. 65,66 Yet, initial X inactivation in females occurs in utero, 1,67 and if there were selective mechanisms for the preservation of activity for X chromosomes with longer telomeres, this should theoretically result in sex differences in telomere length at birth, which, as mentioned earlier, 1,4,68 is not believed to be the case. In fact, the skewing of X inactivation at birth shows no favor for 1 chromosome over the other—the ratio is 50:50 1 —and the skew toward 1 X chromosome over the other is not significantly noticeable until after age 60. 69
Chromosomal explanations for sex differences in telomere length imply that sex differences emerge over the course of the human life span, which is already known to be the case; however, these explanations lack information regarding when sex differences in telomere length become apparent in humans. It is known that many telomere maintenance genes reside on the X chromosome, but it is not known whether these genes are some of the ∼25% that escape inactivation in females and would thus be overexpressed in females. 1,64,70 The skewing of X inactivation toward one chromosome over another in females is not noticeable until after age 60, and the exact mechanism behind this skew is not well-understood—meaning that preservation of the chromosome with longer telomere length may not be the determining factor. 69
In addition to examining alternative biological explanations for sex differences in telomere length, future studies should consider whether differences in environmental, psychosocial, and/or behavioral factors contribute to sex differences in telomere length during early adolescence. If boys spend more time playing outdoors than girls, this could result in different patterns of exposure to environmental contaminants such as lead in soil and air pollution, which have been linked to shorter telomere length. 71,72 Given that social support is positively associated with telomere length—and numerous other health-related outcomes—among adults, 73,74 the tendency for girls to have closer, more supportive relationships with parents, teachers, and peers 75 could also help explain why girls have longer telomeres than boys during early adolescence. Finally, differences in health behaviors, including smoking and dietary patterns, which have previously been found to contribute to sex differences in telomere length among older adults, 76 may help explain the emergence of male/female differences in telomere length during early adolescence.
Conclusions
In this sample of children aged 8 to 14 years, we found that girls had significantly longer telomere length than boys. While we did not find support for the hypothesis that sex differences in telomere length are attributable to male/female differences in sex steroid hormones or body size, larger studies with greater variability in pubertal development are needed to confirm or refute these findings. Moreover, longitudinal studies are needed to estimate associations of within-person change in sex hormones and body size with within-person change in telomere length during the pubertal transition. Future research should also consider whether environmental, psychosocial, and behavioral factors contribute to sex differences in telomere length during this stage of development. Gaining a better understanding of the mechanisms that underlie the emergence of sex differences in telomere length could help explain why women tend to live longer, healthier lives than men. 77
Footnotes
Acknowledgments
The authors wish to thank the American British Cowdray Medical Center for providing study facilities.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the NIEHS/EPA funded University of Michigan School of Public Health Formative Children’s Environmental Health and Disease Prevention Research Center (P20 ES018171/RD834800), NIEHS/EPA funded Lifecourse Exposures and Diet: Epigenetics, Maturation, and Metabolic Syndrome (P01ES022844/RD-83543601), and the National Institute of Public Health/Ministry of Health of Mexico