
Abstract
Introduction:
The long head of the biceps (LHB) is often resected in shoulder surgery. However, its contribution to inflammatory processes in the shoulder remains unclear. In the present study, inflamed and noninflamed human LHBs were comparatively characterized for features of inflammation.
Materials and methods:
Twenty-two resected LHB tendons were classified into inflamed (n = 11) and noninflamed (n = 11) samples. For histological examination, samples were stained with hematoxylin eosin, Azan, van Gieson, and Masson Goldner trichrome. Neuronal tissue was immunohistochemically visualized. In addition, specific inflammatory marker gene expression of primary LHB-derived cell cultures were analyzed.
Results:
Features of tendinopathy, such as collagen disorganization, infiltration by inflammatory cells, neovascularization, and extensive neuronal innervation were found in the tendinitis group. Compared to noninflamed samples, inflamed LHBs showed a significantly increased inflammatory marker gene expression
Conclusion:
Structural and biomolecular differences of both groups suggest that the LHB tendon acts as an important pain generator in the shoulder joint. These findings can, on the one hand, contribute to the understanding of the biomolecular genesis of LHB tendinitis and, on the other hand, provide possibilities for new therapeutic approaches.
Keywords
Introduction
The long head of the biceps (LHB) tendon is an anatomical structure within the shoulder which is a frequent source of pain and pathology. The LHB tendon originates from the superior labrum and the supraglenoid tubercle of the shoulder, giving rise to its intra-articular portion of the tendon. 1 While crossing the rotator cuff interval between the anterior edge of the supraspinatus tendon and the superior edge of the subscapularis tendon, the LHB tendon is stabilized in the bicipital groove by the capsuloligamentous biceps pulley complex. 2 The LHB tendon may be affected by several pathologic mechanisms. Due to its complex anatomical composition and course, the LHB tendon has been identified as a frequent source of anterior shoulder pain and is likely to be symptomatic in shoulders with concurrent soft tissue pathologies such as rotator cuff (RC) tears or labral tears and cartilage degeneration. In fact in over 90% of the cases, disorders of the LHB tendon are associated with RC tears. 3 However, the biomolecular processes responsible for the onset and extent of shoulder pain and degeneration associated with LHB tendon pathology remain unclear. In cases with LHB pathology, tenotomy and removal of the tendon have consistently provided appropriate pain relief while biceps muscle function could be successfully maintained. 4 In the clinical practice, predominantly two different morphologies of the LHB stump after tenotomy can be observed—inflamed and noninflamed. In this work, we analyzed the LHB stumps with and without macroscopic signs of inflammation for their characteristics in the context of inflammation on a tissue, cellular, and molecular level.
Materials and methods
Tissue harvest
After informed consent, LHB tendon samples were obtained from patients undergoing open or arthroscopic surgery for RC repair, LHB tenodesis, or shoulder arthroplasty. The study was approved by the local ethics committee.
Biceps tendon samples after tenotomy were gathered from 22 shoulders in total (mean donor age 59.6 years, range 44–80 years, 13 male, 9 female) of which 11 showed clear macroscopic signs of tendinitis, which were defined as redness and extensive vascularization (see Figure 1). All of these 11 cases presented with clinical signs that might be consistent with LHB pathology, that is, tenderness on palpation of the LHB in the bicipital groove and a positive Yergason and Speed’s test. However, these tests are reported to be often false positive. 5 The donors’ features are presented in Table 1. Any insertion sites (bony/capsular) and the synovial sheaths were removed and tendons were rinsed with saline. The LHB tendons were each cut longitudinally in halves, which were then allocated to further histological or cellular/molecular analyses
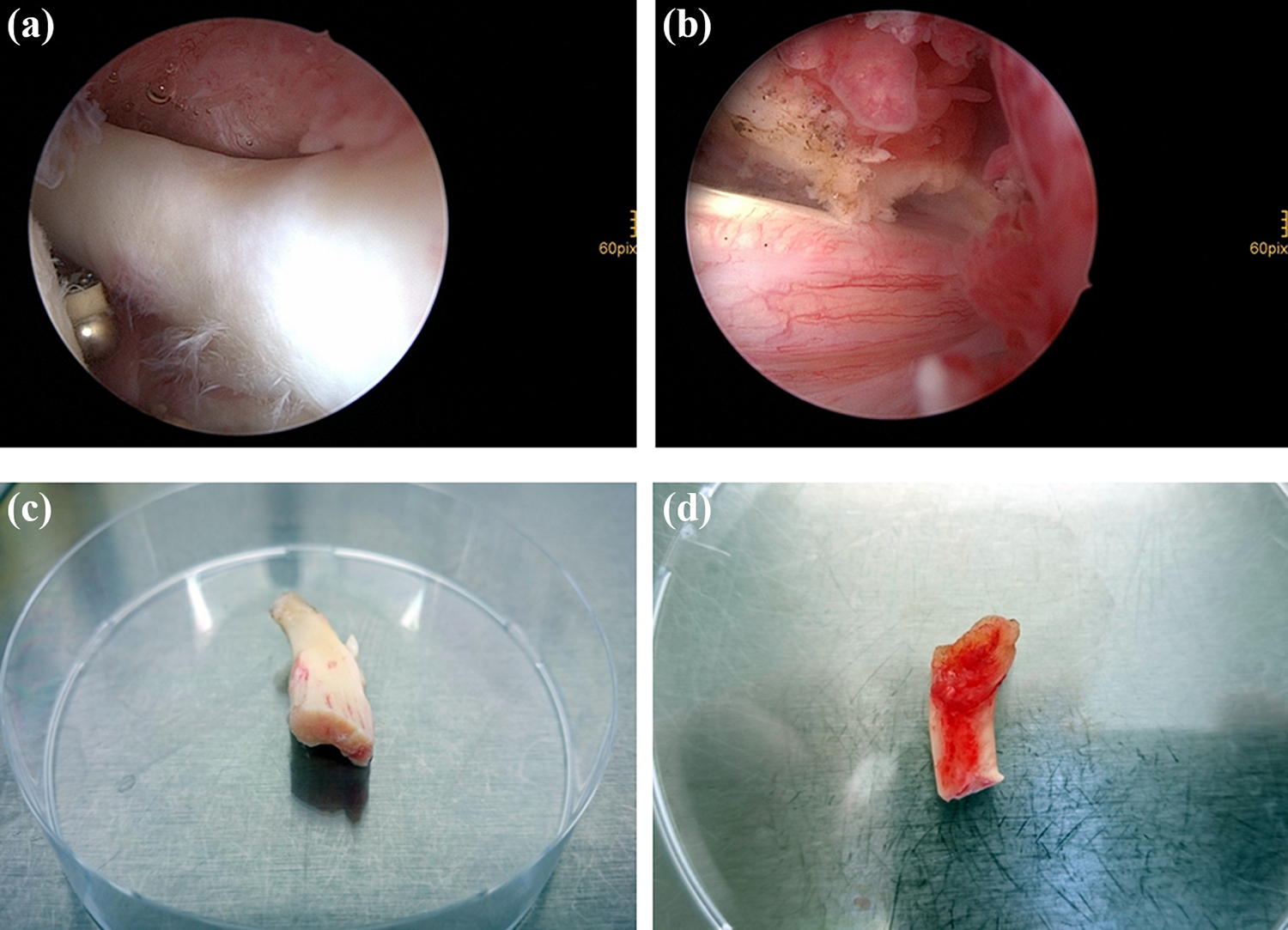
Macroscopic view of inflamed and noninflamed tendon samples. LHB tendon samples with (b, d) and without (a, c) tendinitis are shown macroscopically. Images (a) and (b) show the intraoperative arthroscopic view. Images (c) and (d) show the tendon samples before processing in the laboratory in a 10-cm petri dish. LHB: long head of the biceps.
Donor features including mean age, sex, and reason for surgery.
Histological analysis
Tendon portions for histological examination from the intraarticular portion of the tenotomized LHB were first embedded in 4% formaldehyde. Before staining, the tendon samples were dehydrated in increasing concentrations of ethanol, cleared in xylene, and embedded in paraffin, sectioned at 5 µm thickness and mounted on microscope slides. Tissue morphology was examined by hematoxylin and eosin (HE), Azan, van Gieson, and Masson Goldner trichrome staining. Morphological cell assessment was performed using a phase-contrast microscope (Zeiss, Jena, Germany). Immunohistochemical neuron-specific enolase (NSE) staining was performed to visualize nerve roots in the LHB samples. Therefore, the sections were deparaffinized, and antigen unmasking was performed by incubating in saponin (0.05%) for 30 min. The endogenous peroxidase activity was blocked by treatment with 0.3% hydrogen peroxide in methanol for 30 min. Nonspecific binding sites were blocked by incubation with horse serum (Vectastain ABC kit, Vector Laboratories, Brulingame, California, USA) at a concentration of 200 µl/field for 30 min at room temperature. The NSE antigen was labeled with the primary antibody rabbit-2-human-neuron specific enolase (Spring Bioscience, Pleasanton, California, USA) diluted 1:300 in phosphate-buffered saline (PBS) and incubated overnight at 4°C followed by five washings with PBS. A secondary biotinylated antibody was added at 200 µl/field for 10 min at room temperature to bind to the primary antibody followed by washing for five times with PBS. Afterward, Vectastain ABC reagent (Vector Laboratories) was used as an enzyme label to bind the secondary antibody and after washing with PBS peroxidase substrate solution vector NoraRed (Vector Laboratories) was applied for colorimetric detection. Finally, the sections were counterstained with hematoxylin and mounted in glycerol–gelatin (1:1) (Sigma-Aldrich, Munich, Germany).
Isolation of primary cells
The tendon tissues were minced into small pieces and incubated with collagenase (0.175 U/ml; Sigma-Aldrich) in Dulbecco’s modified Eagle’s medium (DMEM)/Ham’s F12 (1:1) (Life Technologies GmbH, Germany) for 2 h at 37°C. After washing off the digested tendon material and centrifugation to high-density pellets, the released cells could be collected and plated in 75 cm2 tissue culture flasks (Greiner Bio-One, Frickenhausen, Germany). The cells were cultured in DMEM/HAM’s F12 (1:1) supplemented with 10% fetal calf serum (FCS, Biochrom AG, Berlin, Germany) and 1% penicillin/streptomycin in an incubator in a humidified atmosphere (37°C, 5% carbon dioxide) with a change of medium every 2–3 days. As soon as cultured cells reached 90% confluence, they were harvested for RNA isolation or seeded in well plates.
RNA isolation, RT-PCR, and qRT-PCR
Total RNA was harvested from cells passage 0 at confluence. Samples were isolated and real-time polymerase chain reaction (RT-PCR) as well as quantitative RT-PCR (qRT-PCR)were performed as described previously. 6,7 Elongation factor 1α (EF1A) served as the housekeeping gene, target-specific sense, and antisense primers used for RT-PCR analyses are listed in Table 2, which also provides a summary of the primer details. Sequence specific primers for qRT-PCR were obtained from biomers.net GmbH (Ulm, Germany) or Qiagen GmbH (Hilden, Germany) and are listed in Table 3, which also shows the primer specific details. Dissociation curve analysis for qRT-PCR results was carried out to verify the absence of primer dimers and/or nonspecific PCR products. The expression of the genes of interest was normalized against the ribosomal protein S27a (RPS27A) housekeeping gene using the delta–delta C t method. 8
RT-PCR primer sequences and PCR conditions.

A: antisense; Col1A1: collagen type 1α1; COL3: collagen type 3, DCN: decorin; EF1A: elongation factor 1α; IL1B: interleukin 1β; MMP (-1/-3/-9/-13): matrix metalloproteinase (1/3/9/13); S: sense.
qRT-PCR primer sequences and PCR conditions.

A: antisense; Col1A1: collagen type 1α1; COL3: collagen type 3, DCN: decorin; EF1A: elongation factor 1α; IL1B: interleukin 1β; MMP (-1/-3/-9/-13): matrix metalloproteinase (1/3/9/13); S: sense TNFα: tumor necrosis factor α; TIMP1: tissue ihibitor of matrix metalloproteinase 1; VEGF: vascular endothelial growth factor.
Genomewide gene expression profiling of cells isolated from inflamed and noninflamed LHB
For genomewide gene expression profiling, hybridization experiments were performed and analyzed using Affymetrix Gene Chips HG-U133 Plus 2.0 (54,000 probesets for 47,400 transcripts and 38,500 genes, High Wycombe, UK) as described previously. 9 Total RNA expression of three inflamed and two noninflamed LHB samples was analyzed separately using the Affymetrix Gene Chip Scanner 3000, Santa Clara, California, USA, the Affymetrix GeneChip Operating Software 1.4, and comparatively evaluated using the significance analysis of microarrays (SAM) approach. To assess differentially expressed genes between inflamed and noninflamed tendon samples, predefined conditions were established: the number of “present” calls for a given gene had to be >50% in at least one of the groups and only those genes were taken into account that displayed a fold change (FC) <0.5 and >2. Probesets that showed an FC between 0.5 and 2 were stated as “not differentially expressed.” To obtain reliable data, the q-value, that is, false discovery rate had to be <10%. For heatmap generation, mapping was performed with all differentially expressed probesets using the web service for microarray data analysis CARMAweb. 10
Statistical analysis
Statistical analysis was performed using IBM SPSS 22.0, and p values <0.05 were considered significant. Determination of the statistical significance between groups was performed using Student’s t-test or the Mann–Whitney U test as indicated.
Results
Clinical classification in inflamed and noninflamed tendons
Classification in inflamed and noninflamed tendons was realized during surgery upon clinical judgment. Tendons with clear macroscopic signs of inflammation including synovitis and tissue thickening were assigned to the “tendinitis” group, whereas tendons without macroscopic signs of inflammation were assigned to the “no tendinitis” group. Macroscopic images were taken from all samples and representative photographs are shown in Figure 1.
Histological characterization of tissue sections from the LHB
HE stain illustrates the general cellular composition of the tissue with its structural units. Three different collagen stains were used, showing positive staining for Azan, van Gieson, and Masson Goldner trichrome, thus verifying the rich presence of collagens within the biceps tendon. The tendinitis group showed histological evidence of tendinopathy such as decreased tissue organization with dramatic collagen disorganization with increased, randomly oriented tendon fibers as well as fiber cross-linking and excessive infiltration with inflammatory cells representing typical signs of fibrotic degeneration and scar tissue formation (Figure 2). In addition, neovascularization and increased neural innervation could be observed. To confirm these findings, NSE immunohistochemical staining was performed. A high density of free nerve endings could be observed especially at the transition zone to the paratenon in inflamed tendons compared to no tendinitis samples (Figure 3).
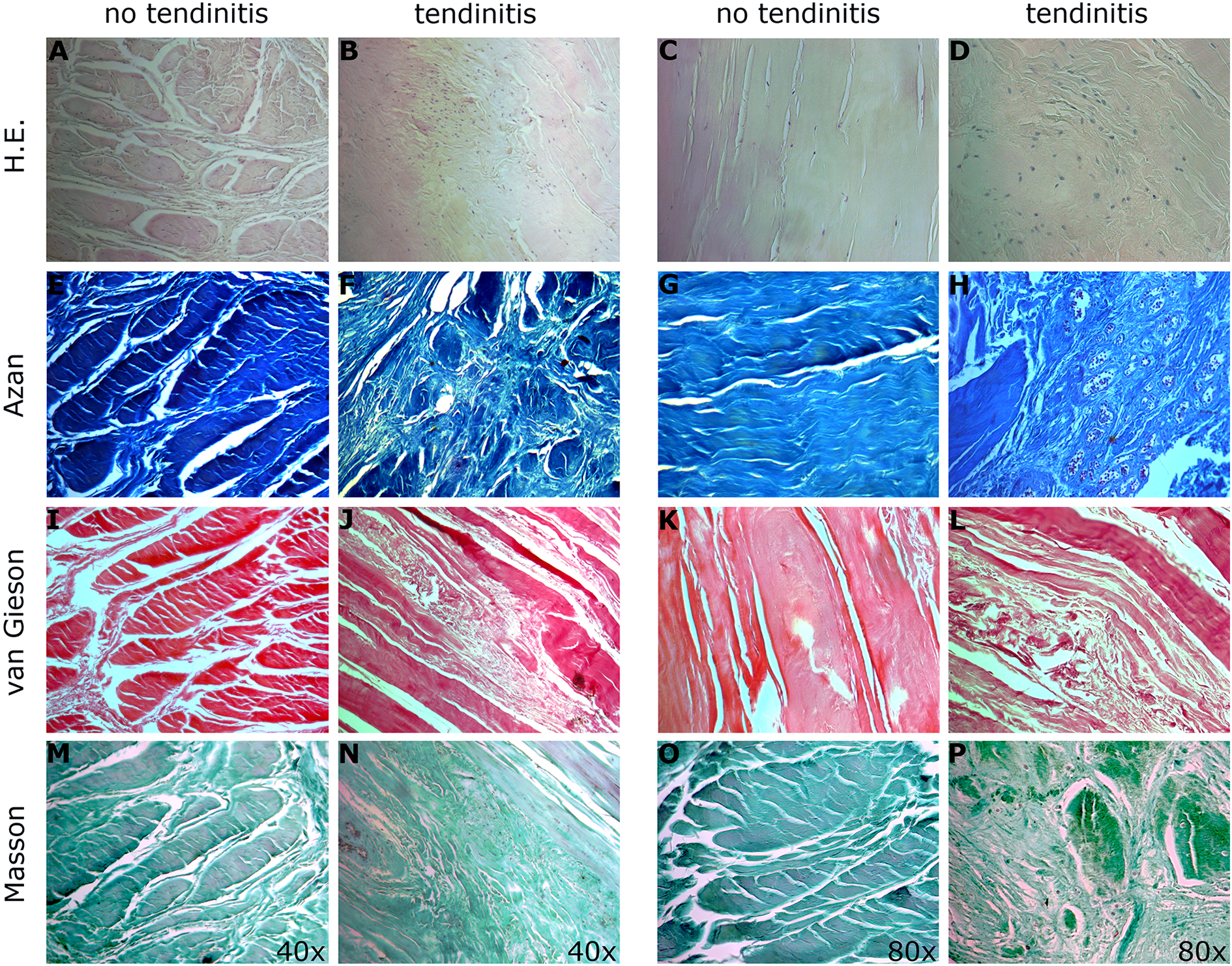
Histological evaluation. Tissue sections were stained for gross histological evaluation with hematoxylin and eosin (a–d) and for collagen distribution using Azan (e–h), van Gieson (i–l), and Masson Goldner trichrome (m–p). Representative images of LHBs with or without tendinitis were obtained from four different donors and representative images are shown. Images are taken at ×40 and ×80 as indicated.

Immunohistochemical NSE staining for visualization of nerve roots. Images a, c, and e show a noninflamed LHB in different magnifications (×5, ×40, ×80) without any nerve tissue. Images b, d, and f (×5, ×40, ×80) display histological evidence of NSE-positive nerve roots in LHBs with massive tendinitis. A high density of free nerve endings can be observed at the transition zone to the paratenon. NSE: neuron-specific enolase; LHB: long head of the biceps.
Qualitative and quantitative RT-PCR analysis
In RT-PCR analyses (Figure 4(a)), inflammatory marker genes like interleukin 1β (IL1B), matrix metalloproteinase 1 (MMP-1), matrix metalloproteinase 3 (MMP-3), and matrix metalloproteinase 9 (MMP-9) as well as matrix metalloproteinase 13 (MMP-13) were upregulated in inflamed LHB samples. Furthermore, genes influencing extracellular matrix (ECM) production such as decorin (DCN), collagen type I alpha 1 (COL1A1), and collagen type III (COL3) were overexpressed in comparison to the noninflamed tendons. Similar findings were revealed by qRT-PCR analysis (Figure 4(b)) showing significant overexpression of pro-inflammatory marker genes such as interleukin 1α (IL1A), IL1B, and tumor necrosis factor α (TNFα), MMP-1, and MMP-9 in the tendinitis group. In contrast, anti-inflammatory marker genes such as tissue inhibitor of matrix metalloproteinase 1 (TIMP-1) were significantly upregulated in noninflamed LHB samples compared to inflamed tendons. The expression of vascular endothelial growth factor (VEGF) was significantly higher in LHB tendons with inflammation. Considering the expression of ECM marker genes, a significant upregulation of the COL3 expression could be shown in the tendinitis group, with the COL1 to COL3 expression ratio being significantly decreased in inflamed tendon samples compared to noninflamed ones.
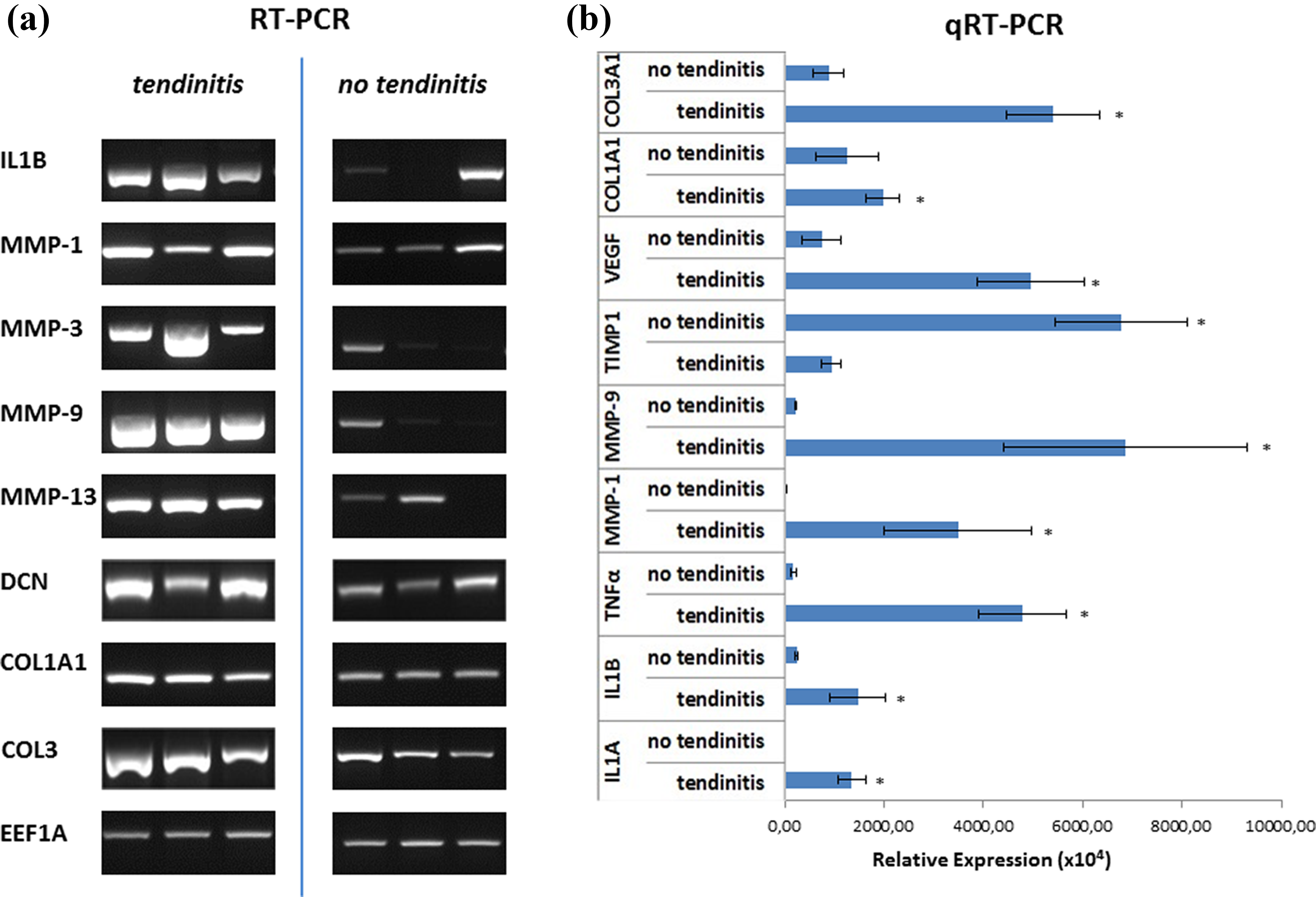
RT-PCR (a) and qRT-PCR (b) analysis of inflamed and noninflamed tendon samples. Expression patterns of Col1A1, Col3A1, DCN, IL1A, IL1B, MMP-1, MMP-3, MMP-9, MMP-13, TIMP1, TNFα, and VEGF in LHB samples with and without tendinitis. For RT-PCR EF1A and for qRT-PCR RPS27A served as housekeeping genes for normalization of the expression values. Significant differences between the two groups are indicated by asterisks as determined by t-testing. RT-PCR: real-time polymerase chain reaction; qRT-PCR: quantitative real-time polymerase chain reaction; Col1A1: collagen 1 type alpha 1; Col3A1: collagen 3 type alpha 1; DCN: decorin; IL1A: interleukin 1 alpha; IL1B: interleukin 1 beta; MMP-1: matrix metalloproteinase 1; MMP-3: matrix metalloproteinase 3; MMP-9: matrix metalloproteinase 9; MMP-13: matrix metalloproteinase 13; TIMP1: tissue inhibitor of matrix metalloproteinase 1; TNFα: tumor necrosis factor alpha; VEGF: vascular endothelial growth factor; EF1A: elongation factor 1α; RPS27A: ribosomal protein S27a.
Microchip hybridization of RNA from inflamed and noninflamed LHB tendon samples
Comparative microchip hybridization analysis of the RNA from three donors of inflamed and two donors of noninflamed LHB tendon samples revealed a different gene regulation pattern. Significance analysis of microarray (SAM) is summarized in Figure 5(b), showing a Venn diagram, depicting the upregulation of 895 probesets in cells isolated from LHB tendons showing macroscopic signs of inflammation compared to cells isolated from noninflamed LHB tendons, and the downregulation of 484 probesets in contrast, as well as a total of 25,954 unregulated probesets. The 50 most important upregulated and downregulated probesets are depicted in a heatmap (Figure 5(a)), showing remarkable expression differences between cells from inflamed and noninflamed tendons, for example, in genes associated with pain such as Cathepsin H (CTSH), interferon-inducible protein variant 5 (CXCL5), potassium voltage gated channel, shaw-related subfamily, 3 (KCNS2), and ATPase 6 (ATP6V1 H). 11,12

Comparison of microchip hybridizations for RNA from cells isolated from inflamed and noninflamed tendon samples. Regulation of selected probesets from two donors without tendinitis and three donors with tendinitis. Upregulated probesets are represented by red areas, downregulated by green ones with light colors indicating stronger regulation than darker colors. Next to the heatmap probeset IDs and the abbreviation of the examined genes (a) are found. SAM revealed the number of probesets, which were upregulated (red circle) and downregulated (green circle) in the tendinitis group compared to the no tendinitis group as well as the number of unregulated probesets (intersection) (b). SAM: significance analysis of microarray.
Discussion
The LHB tendon is often implicated in shoulder pain and a degenerated or inflamed LHB frequently coexists with RC tendinopathy and impingement syndrome; however, its role in shoulder pain remains controversial and incompletely understood. 13
Our study focused on the molecular inflammatory pattern in the LHB tendon comparing samples with and without macroscopic signs of tendinitis in combination with a histological analysis of changes in tendon structure to reveal new information for future therapeutic approaches. All patients with macroscopic signs of LHB tendinitis presented with clinical signs that might be consistent with LHB pathology, that is, tenderness on palpation of the LHB in the bicipital groove and a positive Yergason and Speed’s test. In contrast, only 3 of the 11 patients without macroscopic signs of tendinitis clinically presented with positive LHB tendinitis tests. These three patients underwent isolated biceps surgery due to a superior labral tear from anterior to posterior (SLAP). However, in histology and in gene expression, these samples did not show any sign of inflammation. In contrast, histological analysis revealed typical signs of fibrotic degeneration such as disorganization of collagen fibers and infiltration with inflammatory cells in the tendinitis group. In addition, immunohistochemical staining of inflamed samples showed a high density of nerve endings especially in the transition zone to the peritendineum and an increased vascularization. Singaraju et al. observed an increased number of nerve roots in LHB samples of symptomatic patients with RC disease confirming these findings. 14
Genetic upregulation of cytokines, including IL1A, IL1B, and TNFα, are intimately involved in the facilitation of inflammation and the generation of pain. 15 Genetic upregulation of these cytokines has been shown to play an important role in the development of symptoms in osteoarthritis in an animal model and is known to induce overexpression of catabolic enzymes such as MMP-1, -3, -9, and -13. 16,17 The overexpression of these pain-related cytokines and enzymes is supported by the results of the genomewide expression profiling showing a significant upregulation of pain-related genes such as CTSH and CXCL5 in the tendinitis group. These genes have been shown to be associated with pain in multiple independent experiments. 11,12
Furthermore, overexpression of these proinflammatory genes has been attributed to the cause of apoptosis in RC tendinopathy. 18 In fact, apoptotic features such as upregulation of pro-apoptotic Bcl-2 family proteins have been demonstrated in chondrocytes and synoviocytes treated with IL1B or TNFα, and these cytokines may induce apoptosis by provoking oxidative stress through nitrogen oxide (NO) production. 19,20 MMPs are a family of zinc-dependent endopeptidases responsible for maintaining the dynamic homeostasis of the extracellular matrix (ECM). Tissue inhibitors of MMPs (TIMPs) are the natural, endogenous opponents of MMPs and are essential players in the reparative and degradative processes of the extracellular matrix. Notably, both matrix metalloproteinase activity and pro-inflammatory cytokine production are associated with RC disease. 21 Bedi et al. showed that local inhibition of the expression of MMPs at the footprint in a rat supraspinatus repair model results in an increased fibrocartilage and reduced collagen degradation in the MMP-inhibited group compared with controls. 22 Location and RC-tear size may influence MMP biology as LHB samples showing clear signs of degeneration taken from patients with articular-sided partial-thickness RC tears were reported to have higher levels of MMP-1 and MMP-9 compared to patients with partial tears on the bursal side. 23 This study shows a significant overexpression of MMP-1, -3, -9, and -13 in inflamed LHB tendons, whereas the TIMP1 expression was downregulated indicating a severe imbalance between destructive and protective enzymes.
VEGF is known to have potent angiogenic effects, and hypoxia has been recently correlated with increased apoptosis in the RC. 24 In our study, an increased neovascularization in the peripheral parts of inflamed LHB tendons could be observed in histology, in addition, VEGF expression levels were significantly higher in the tendinitis group.
Regarding ECM production, inflamed samples showed increased ECM turnover and revealed DCN overexpression—an important regulator of collagen fibrillogenesis. 25 Increased expression of collagen type III, as it could be observed in the tendinitis group, was related to tendinopathy including tears in various studies. 26,27 Collagen type III fibrils have a thinner diameter and are more elastic as well as less organized than collagen type I. The ratio of collagen type I to collagen type III was decreased in inflamed specimens. A decreased collagen type I—collagen type III ratio may be interpreted as a sign of tissue remodeling and has previously been associated with decreased mechanical stability. 28 Collagen type III is normally synthesized in early stages of wound healing and is considered to be an immature form of collagen type I. 28,29
This data are in agreement with upregulation of the markers associated with inflammation, matrix turnover, and vascularization in inflamed bursa tissues, indicating that similar basic molecular mechanisms are present in tendon tissue and bursa upon inflammatory shoulder disease. 7
Conclusion
In this study, the structural and the biomolecular differences of inflamed and noninflamed LHBs are shown. In samples with tendinitis, the gene expression of the pro-inflammatory cytokines IL1A, IL1B, and TNFα and of the catabolic enzymes MMP1,-3,-9, and -13 was upregulated, whereas the expression of the anti-inflammatory gene TIMP1 was downregulated. Furthermore, vast histological changes such as disorganization of the collagen fibers, neovascularization, and increased neural innervation could be observed in the tendinitis group. These findings can contribute to the understanding of the disease mechanisms of LHB tendinitis, which might be harnessed for the development of novel therapeutic approaches.
Footnotes
Acknowledgments
We thank Beate Geyer and Melanie Krug for excellent technical assistance.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Federal Ministry of Education and Research (FKZ 13N13454, “AutoLoop”) and the Novartis Pharma GmbH.