
Abstract
Mineralized bone is increasingly considered as a functional substrate for regenerative applications, yet its impact on neuron–glia remodeling remains insufficiently defined. We cultured ex vivo injured rat hippocampal tissue on glass coverslips either uncoated or coated with micron-scale mouse skull bone particles and quantified neurite architecture and astrocyte morphology. Bone-particle substrates supported robust adhesion and selectively modulated process development. Axons on bone displayed a 2.8-fold increase in varicosity-like expansion size and 2.2-fold higher neurofilament-M expression relative to glass, indicating potentiated axonal sprouting. In contrast, dendrites exhibited 20% shorter mean length and 73% lower branching. Astrocytes on bone showed 21% shorter processes with 34% fewer processes per cell; total cell area and GFAP levels were unchanged. However, astrocytes displayed increased circularity, decreased roundness, and elevated solidity—morphologies consistent with a reactive, potentially chronic, state. Together, these data identify mineralized bone as a bioactive osseous substrate that enhances axonogenesis while biasing astrocytes toward reactivity. This divergence suggests design trade-offs for osseous or mineral-hybrid scaffolds aimed at central nervous system repair. Our findings provide quantitative guidance for engineering bone-derived or mineral-composite scaffolds that differentially control neuronal and glial outcomes in neural repair strategies.
Keywords
Introduction
The clinical demand for effective brain repair strategies remains significant due to limited central nervous system regenerative capacity following injury or disease. Traumatic brain injury (TBI), stroke, and neurodegenerative disorders commonly result in irreversible neuronal loss, glial scarring, and disrupted neural circuits, leading to persistent functional deficits.1,2 Current therapeutic approaches primarily focus on symptom management rather than restoration of neural tissue or circuitry. In this context, biomaterial-based scaffolds offer promising potential to bridge damaged tissue, mitigate inflammation, and support the growth or replacement of neural cells. 3 The design of scaffolds that are both biocompatible and capable of guiding organized tissue regeneration is therefore a key objective in the development of CNS repair strategies.
Tissue engineering efforts for neural repair have largely centered on soft, biocompatible scaffolds that mimic the extracellular matrix and support cell adhesion, growth, and differentiation. Numerous materials have been explored, including natural polymers such as hyaluronic acid,4–7 chitosan,8–11 alginate,12–14 gelatin,15,16 nanomaterials, 17 as well as composite organic and inorganic hybrid scaffolds incorporating stem cells. 18 Hydrogels offer distinct advantages over rigid scaffolds by absorbing water to expand and conform to irregular wound geometries while serving as delivery vehicles for trophic factors or stem cells. 19 Recent innovations in injectable hydrogels have enabled greater control over rheological properties, injectability, and biphasic permeability, facilitating the design of functionally tailored biomaterials.20–22 Such properties support critical relationships between scaffold structure and biological integration. 2 Synthetic polymers including poly-L-lactic acid,23,24 poly-D,L-lactic-co-glycolic acid,25–28 and conductive materials like polypyrroles and carbon nanotubes29–31 have also been investigated. Nevertheless, these non-native materials raise safety concerns, including potential cytotoxicity and immunogenicity, underscoring the value of natural biopolymers in clinical applications. 32
Despite their advantages, hydrogels possess limitations in neural tissue engineering. Their inherently low mechanical strength restricts their use in load-bearing or interface regions, such as the brain–skull junction.33–35 Furthermore, their degradation kinetics can be difficult to control, potentially leading to scaffold loss before tissue regeneration is complete. 36 Hydrogels can also induce local pressure changes through swelling, which is particularly problematic in the confined spaces of neural tissue. 37 Additionally, their lack of anisotropic structure reduces their capacity to guide axonal growth or support proper synaptic architecture. 38 Most hydrogels are electrically insulating and, while conductive additives can improve electrical compatibility, these additives often introduce new biocompatibility challenges. 39 Clinically, hydrogels also suffer from fragility and inconsistent in situ formation.40–42 Together, these issues suggest a need for alternative or hybrid scaffolds that combine the bioactivity of soft materials with the mechanical and structural advantages of solid scaffolds.
Solid scaffolds—including ceramics and rigid biomaterials—offer mechanical strength and architectural stability that address many of the shortcomings of soft polymers. These properties are particularly beneficial in applications requiring long-term support or load-bearing function, such as spinal cord and craniofacial repair.43,44 Solid scaffolds maintain their form under physiological conditions, providing consistent structure for tissue remodeling and cell alignment, yet choice of scaffolding materials and their stability for proper cell alignment depends on the desired outcome: directed or random alignment. For example, aligned scaffolds of electrospun polymers or poly-L-lactic acid have been successfully used to direct skeletal and cardiac muscle cells for graft development where linear tissue alignment is desired. In contrast, random scaffolds constructed from poly(lactic-co-glycolic) acid (PLGA) or random electrospun structures have been used for graft development where structural flexibility is preferred and tend to be stronger than linear aligned scaffolds. 45 Materials such as hydroxyapatite, β-tricalcium phosphate, and bioactive glass have been widely used due to their chemical similarity to natural bone, promoting osteoconduction and integration.46,47 Surface modifications—including coatings with ECM proteins, bioactive peptides, or ions—further enhance cellular adhesion and lineage-specific differentiation. 48
In neural tissue engineering, solid scaffolds—particularly when microstructured or doped with conductive materials—can guide axonal orientation, promote synaptic formation, and facilitate electrical communication between neurons. 49 Some ceramics also exhibit bioinert or bioactive behavior that supports long-term implantation with minimal chronic immune response. 50 Additionally, solid metal oxide scaffolds can be customized using advanced techniques such as 3D printing and electrospinning to create patient-specific geometries and controlled degradation profiles.51–54 Compared with hydrogels, they offer improved spatial fidelity and the ability to support complex tissue regeneration through hybrid composite systems.55,56
Bone-derived materials have long been used in tissue engineering, particularly in regenerating hard tissues like bone, cartilage, and dental structures. Owing to their natural architecture, bioactivity, and mineral content, bone-derived scaffolds support cell adhesion, proliferation, and lineage-specific differentiation in orthopedic and craniofacial contexts.57,58 Forms of bone used include autografts, allografts, decellularized bone matrices, powders, and mineral composites. 59 Mineralized bone, rich in hydroxyapatite and calcium phosphates, provides both structural strength and bioactive signaling. This makes it suitable for load-bearing environments and supports osteoconductive processes.57,60 Moreover, bone degradation rate can be tuned to support gradual tissue replacement.60,61 The surface roughness and microtopography of mineralized bone enhance cell adhesion and differentiation, especially in osteogenic and mesenchymal cell populations.61,62
Though traditionally linked to bone repair, mineralized bone is gaining attention in neural tissue engineering. The bone–nerve interface plays an essential role in various anatomical regions, including the craniofacial skeleton, spine, and peripheral nerve pathways. 63 In cases of neuro-orthopedic injuries or traumatic brain injury with skull fractures, integrating bone with neural scaffolds may be beneficial.62,64 Bone-based substrates may offer mechanical support and topographical guidance for regenerating neural cells in such hybrid environments.
However, the use of bone as a neural scaffold remains largely unexplored. Whether mineralized bone supports or impedes neural regeneration is still unknown. Investigating this is particularly important in cases where bone fragments are present in a CNS lesion, whether introduced surgically or through trauma, and can produce damage. Understanding the interactions between bone material and neural cells is crucial for evaluating its potential benefits or drawbacks in regenerative contexts.64,65
The current study explores this novel direction by examining the effects of mineralized bone on neuronal and astrocytic morphology in vitro. Specifically, we assess whether bone substrates support cellular outgrowth, migration, and structural differentiation. Our findings demonstrate that mineralized bone promotes axonal elongation and increased medium neurofilament expression, while dendritic sprouting is supported by the bone, but to a more limited extent compared with dendrites on glass substrates. Additionally, astrocytes exhibit migratory behavior on bone but also display morphological features indicative of reactive transformation. By characterizing these neural–bone interactions, we aim to provide foundational insights that may inform the design of mineral-based or hybrid scaffolds for CNS repair, particularly in applications where bone proximity is a clinical reality.
Materials and methods
Animals
Housing, care, and experimental procedures involving animal use conformed to NIH/USDA and ARRIVE 2.0 guidelines. The experiments were approved and supervised by the Institutional Animal Care and Use Committee of Ariel University and the Israel Ministry of Health (Protocol AU-IL-2112-110-1).
Bone powder preparation
Skull tissue from adult mice (Sabra-M, Inotiv, Jerusalem, Israel [strain no longer commercially available]; 40 g; 1–3 months) was excised and soaked in double-distilled water (DDW) at room temperature overnight. Soft periosteal and connective tissue residues were manually removed using fine scissors and a scalpel until no visible soft tissue remained. The cleaned skulls were rinsed in fresh DDW and autoclaved (30 min, 121°C, 207 kPa). The sterilized skulls were crushed using a mortar and pestle into grains <30 µm, mixed with DDW to a final concentration of 20 mg/mL, and 50 µL of the suspension was applied to round glass coverslips (12 mm diameter; Menzel-Gläser). Coverslips were heated to 50°C for 30 min to allow water evaporation and particle adhesion, followed by a second autoclaving step.
Given that cortical bone particles were predominantly smaller than 30 µm, with approximately 50% below 10 µm, and considering native murine cortical bone pore diameters (10–80 µm), it is highly improbable that the particles retain original porosity. Thus, the particles used are effectively non-porous, obviating the need for a porous control.
Hippocampal slice culture
A day before culture, bone-coated coverslips were coated with 20 ng/mL poly-D-lysine solution (Cat. P7280, Sigma-Aldrich, Rehovot, Israel) and incubated overnight at 4°C. The next day, the coverslips were washed twice with DDW and dried in a laminar flow hood. Cultures were made from 10 animals, each generating 32 sections (total of 300). The procedure was as follows: postnatal male rats (Sprague Dawley, Inotiv, Jerusalem, Israel; 0–3 day old) were dissected to isolate the two hippocampi. The hippocampi were then manually cut using sterile scalpel blades in both the longitudinal (rostral–caudal, 4 mm, 8 cuts) and transverse (medial–lateral, 2 mm, 4 cuts) axes, generating 32 uniform square sections of 0.5 × 0.5 mm2.The sections were then placed on the prepared coverslips and incubated overnight in “First-day medium” containing 85.1% Minimum Essential Eagle’s medium (MEM) (Cat. M2279, Sigma-Aldrich, Rehovot, Israel), 11.5% heat-inactivated fetal bovine serum (Cat. 04-007-1A, Biological Industries), 1.2% L-Glutamine (Cat. G7513, Sigma-Aldrich, Rehovot, Israel) and 2.2% D-Glucose (Cat. G8769, Sigma-Aldrich, Rehovot, Israel) at 37°C, 10% CO2. The medium was substituted the following day by a growth medium that contained 45% MEM (Cat. M2279, Sigma-Aldrich, Rehovot, Israel), 40% Dulbecco’s Modified Eagle’s medium (DMEM; Cat. D5796, Sigma-Aldrich, Rehovot, Israel), 10% Nutrient mixture F-12 Ham (Cat. N4888, Sigma-Aldrich, Rehovot, Israel), 0.25% (w/v) bovine serum albumin (BSA; Cat. A4503, Sigma-Aldrich, Rehovot, Israel), 0.75% D-glucose (Cat. G8769, Sigma-Aldrich, Rehovot, Israel), 0.25% L-Glutamine (Cat. G7513, Sigma-Aldrich, Rehovot, Israel), 0.5% B-27 supplement (Cat. 17504-044, Gibco, Rhenium Bio Science, Modi’in, Israel), 0.1% kynurenic acid (Cat. K3375, Sigma-Aldrich, Rehovot, Israel), 0.01% of 70% uridine (Cat. U3750, Sigma-Aldrich, Rehovot, Israel), and 30% 5-fluoro-2′-deoxyuridine (Cat. F0503, Sigma-Aldrich, Rehovot, Israel), and incubated again at 37°C, 10% CO2 for 13 d, without further medium replacement.
Immunofluorescence
Samples were fixed using 4% paraformaldehyde (Cat. P6148, Sigma-Aldrich, Rehovot, Israel) for 10 min, permeabilized with 0.25% TritonX100 (Cat. H5142, Promega, IMBH, Beit Haemek, Israel) for 5 min and blocked with 3% Inactivated Normal Goat Serum (Cat. 04-009-1A, Biological Industries, Sartorius, Beit Haemek, Israel) for 1 h; all the procedures above were performed at RT. The samples were then incubated overnight at 4°C with the unconjugated primary rabbit anti-neurofilament M (NFM) antibody (Cat. Ab9034, Abcam, Zotal, Tel Aviv, Israel; 0.002 mg/mL) for visualization of axons, mouse anti-Microtubule-Associated Protein 2 (MAP2) (Cat. Ab11267, Abcam, 0.002 mg/mL) for visualization of neuronal somas and dendrites, and Rabbit anti-Glial Fibrillary Acidic Protein (GFAP) (Cat. Ab7260, Abcam, Zotal, Tel Aviv, Israel; 0.002 mg/mL) for visualization of astrocytes. The next day, the samples were washed three times with Phosphate Buffered Saline 1× (Cat. BP243820, Fisher Scientific, Mishmar HaYarden, Israel) and treated with the secondary antibodies Alexa555-conjugated goat anti-rabbit (Cat. A21429, Life Technologies, 0.004 mg/mL) and Alexa488 conjugated Goat Anti-Mouse (Cat. A11001, Life Technologies, Fisher Scientific, Mishmar HaYarden, Israel; 0.004 mg/mL) for 1 h at RT. The samples were washed five times with Phosphate Buffered Saline 1× and mounted using Fluoromount (Cat. F4680, Sigma-Aldrich, Rehovot, Israel) containing 2.5% of 1,4-diazabicyclo[2.2.2]octane to prevent bleaching of the fluorophores.
Microscopy
An Olympus SZ61 stereoscopic microscope was used to display the skull scaffold before and after the grinding process. Phase contrast and fluorescent images were acquired using the Olympus IX81 inverted microscope, equipped with the Olympus UPIanFL N 10×/0.30 (∞/−/FN26.5) objective, and FITC, TxRed, DAPI filter cubes.
Image sampling and analysis
Image analyses were performed using the Fiji plugin suite of ImageJ. 66 Imaging was intensity-corrected using the following formula: Corrected total cell fluorescence (CTCF) = Integrated Density − (Selected target area × Mean fluorescence of background). RGB pixels were analyzed by weighted conversion using the following formula: Brightness value = 0.299R + 0.587G + 0.114B. Left and right hemisphere hippocampal cultures were imaged (one section per coverslip) in a single field per slide. Each coverslip contained a hippocampal slice cultured atop a centrally applied bone powder region, from which cells and cellular outgrowth extended into both bone-coated and non-bone areas. For quantitative analysis, the coverslip was segmented into bone powder-coated (“coated”) and glass, bone power-free (“uncoated”) regions, and measurements were performed separately within each region to enable comparative assessment.
Bone grain morphology (length and area) was assessed from five substrate-coated coverslips. For each coverslip, 20 individual grains were randomly selected, measuring a total of 100 grains. Length was measured along the longest axis using the segmented line tool and area was outlined using the freehand selection tool. NFM fluorescence intensity and axonal expansion area were quantified from four hippocampal slices, each cultured on a separate coverslip. For each slice, 16–23 axons per region (glass and bone) were measured. Axons were manually outlined, and the mean gray value and area were extracted. For each coverslip, average values per region were calculated, then averaged across samples. NFM intensity was expressed relative to the glass region used as the control (set as 100%). The distribution of axonal expansion areas was compiled from the same dataset.
Dendritic extension length was measured from 10 dendrites per region (glass and bone). Dendrites were traced from the root adjacent to the soma to the terminal tip using the segmented line tool. Dendritic branching density was assessed from three separate sprouting regions. A 20 μm-wide strip along the migration front was marked using the freehand tool and the number of dendrites crossing this line was counted and expressed as dendrites per 20 μm. Astrocytic process number and extension length were analyzed from five hippocampal slices. For each region (glass and bone), 10 astrocytes were outlined manually and the number of extensions per cell was counted. Process length was measured from 25 extensions on glass and 20 on bone, traced individually using the segmented line tool. Astrocyte migration distance, GFAP fluorescence intensity, and astrocyte morphology were evaluated across five hippocampal slices. For each region (glass and bone), 10 astrocytes were selected. Migration distance was measured from the slice edge to the most distal astrocyte using the segmented line tool. For GFAP intensity and cell shape analysis, astrocytes were outlined using the wand tool and analyzed for area, circularity, roundness, and solidity using the shape descriptors tool. GFAP intensity was expressed as the corrected total cellular fluorescence (CTCF), which was calculated by the formula: CTCF = Integrated Density − (Selected target area × Mean fluorescence of background). 67
Circularity, roundness, and solidity were calculated as follows: Circularity = 4π × area/perimeter 2 , where values close to 1.0 represent a perfect circle, and lower values reflect increased surface complexity, such as branching or irregular edges. Roundness = 4 × area/(π × major axis 2 ) quantifies elongation based on the length of the major axis, independent of surface texture; lower values indicate more elongated cell shapes. Solidity = area/convex hull area measures how fully the cell occupies the space defined by its outermost boundary. Lower values reflect fragmented or irregularly spread shapes, whereas higher values indicate a more unified and coherent morphology. Each parameter captures different aspects of cell geometry. While circularity is sensitive to the smoothness and complexity of the cell edge, roundness reflects the overall ratio of the cell, indicating elongation regardless of changes in the cell perimeter. Solidity indicates how fully the cell fills its convex outline, serving as a measure of structural cohesiveness. Together, these parameters provide a comprehensive profile of astrocyte morphology, enabling detection of biologically meaningful differences in cell shape that may result from substrate interaction or reactive transformation.
Statistics
Data are expressed as mean ± standard deviation. Statistical significance between samples was calculated using GraphPad Prism 10.1 software by unpaired t-test, Welch’s t-test in cases of unequal variances, or lognormal Welch’s t-test for non-Gaussian, non-homogeneous datasets (α = 0.05). Analysis of percent change in NFM intensity was performed using a one-sample ratio t-test due to lognormal distribution of bone dataset values (α = 0.05). Homogeneity of variances was assessed using an F-test. Gaussian distributions were determined by consensus results of Anderson–Darling A 2 , D’Agostino–Pearson K 2 , and Shapiro–Wilk W tests. Lognormality was assessed using Akaike Information Criterion (AIC) comparison. Image analysis was performed using ImageJ (version 1.54f; standard and Fiji plugins).
Results
We established a reliable and reproducible method of growth substrate preparation consisting of bone powder prepared from mouse skull bone tissue. Skulls of 1-3-month-old mice were removed (Figure 1(a)) and manually cleaned, as described in the methods section. Prepared bone was then manually ground into powder using a mortar and pestle (Figure 1(b)–(d)), producing grains with an average length of 24.02 ± 21.86 µm, and an average area of 296.57 ± 676.17 µm2. Grain length and width ranged between 3.18 µm and 121.18 µm (Figure 1(e)), producing areas between 4.43 µm2 and 4675.72 µm2 (Figure 1(f)). Bone grains were affixed to glass coverslips and seeded with slices of rat hippocampus (Figure 1(g)). Note that bone powder grains tended to clump into semi-organized regions with regions nearly devoid of bone particles between. Growth in devoid areas (glass) served as a control.

Culture of hippocampal slices on bone powder. A system for growing hippocampal slices on bone powder: (a) cleaned skull bone tissue (1 to 3-month-old mice), (b) bone powder grains, (c) phase contrast closeup of bone grains (10× objective), (d) digital magnification of an area in (c), (e) distribution of grain length on the longest axis, (f) distribution of grain area, and (g) representative hippocampal slice (yellow dot, 13 days in culture) cultivated on bone powder (light blue dot) coated coverslip. Red arrows: bone powder-coated regions. Dark blue dots: uncoated glass surface.
Bone powder was then tested for effects on neurite sprouting in mouse hippocampal slices by immunohistochemistry staining with anti-neurofilament M (NFM). Figure 2(a) to (c) show that axons strongly sprout from the slice, extending both toward the bone powder and the uncoated glass regions. The axons that extend from the slice onto the glass often crossed the bone powder region boundary and grew and adhered to bone particles (Figure 2(d)–(f)). Images of axons on the bone powder revealed fibrous shafts and enlarged growth expansions that were absent from axons on the glass (Figure 2(f)). Axons on powder also appeared to turn and change directions frequently, forming a complex mesh, and expressed higher levels of NFM than axons on glass (Figure 2(g) and (h); for quantification, see Figure 3 below).
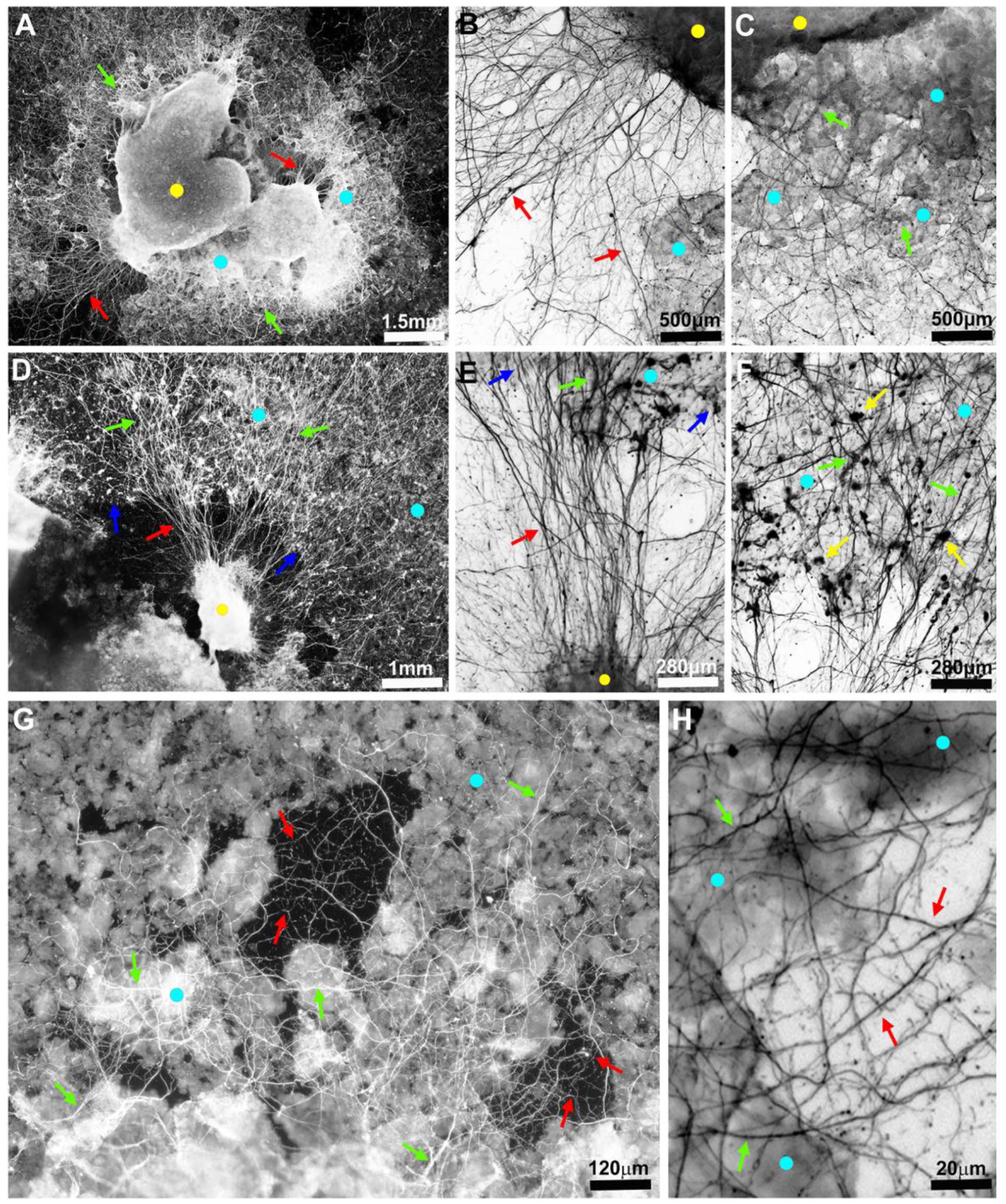
The axons sprout on bone powder. Images of fluorescent-labeled axons in 13-day-old hippocampal slice culture. In all images - yellow dots = slices; light blue dots = regions coated with bone powder; red arrows = axons on uncoated glass regions; green arrows = axons on bone powder. Images in b, c, e, f, and h are inverted: (a) sprouted axons on uncoated glass regions (red arrows) and on bone powder (green arrows), (b) high magnification of axons extending from a slice onto uncoated glass region, (c) high magnification of axons extending from a slice onto bone powder, (d) axons originating from a fraction of a slice crossing the powder layer border (blue arrows) and growth on coated and uncoated regions, (e) high magnification of the central uncoated region in (d), (f) high magnification of the central coated region in (d), (g) close up on axons sprouting and extending on bone grains and bone grain-devoid glass areas, and (h) close up of the axons grown in the bone grain-devoid areas of (g).
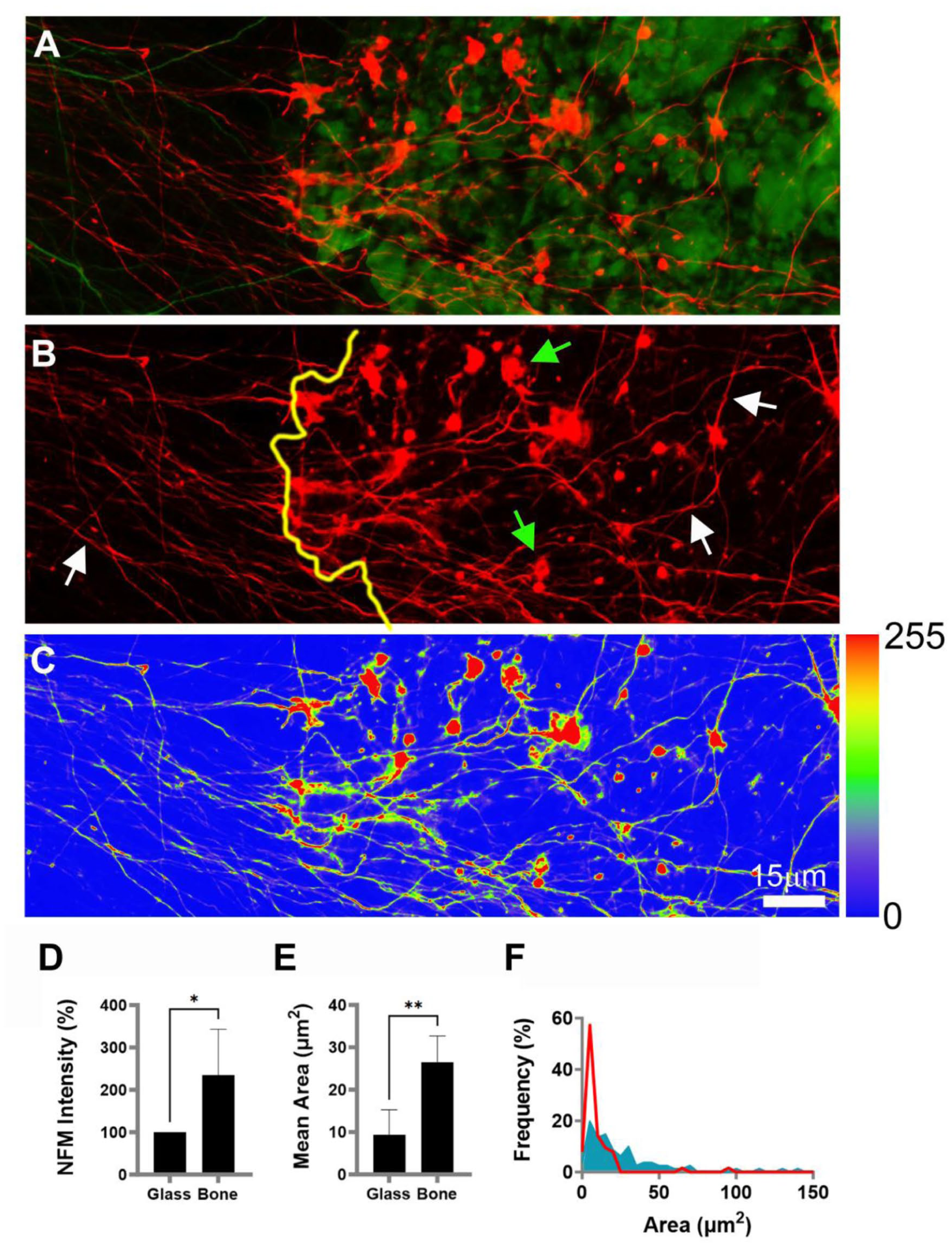
Axons enlarge and increase NFM expression on bone powder. The fluorescence images show axons (red) on bone powder (green) in a 13-day-old culture: (a, b) axons extend from left (glass) to right (bone powder, green). The yellow line indicates powder border. White arrows: axonal shafts. Green arrows: axonal expansions, (c) color-coded version of (a) with accompanying relative intensity scale (right side), (d) average NFM intensity (p = 0.0423), (e) average area of axonal expansions (p = 0.0069), (f) distribution plot of axonal expansion size; red line: bone grains; blue area, glass.
The extent of these axonal reactions to bone contact was quantified and is shown in Figure 3. Image analysis of axons crossing the glass-bone powder boundary allowed quantitative comparison of axonal growth morphology on both surface substrates (Figure 3(a)). We found that axons traversing glass (Figure 3(b), to the left of the yellow border line) are thinner, have a lower NFM level, and possess smaller arborizations than axons located on bone powder (Figure 3(b), to the right of the yellow line). In Figure 3(c), NFM staining intensity was heat-mapped and corroborated these findings, indicating a higher NFM level on bone than glass. NFM fluorescent intensity was 2.3-fold higher in axons on bone than on glass (one-sample ratio t-test, t[3] = 3.405, p = 0.0423; Figure 3(d)). Growth cone-like arborizations of axons grown on bone powder were 2.8-fold larger than those grown on glass (26.56 ± 27.6 µm2 vs 9.3 ± 14.2 µm2; t[6] = 4.029, p = 0.0069; Figure 3(e)). The plot distribution of the axonal area on bone particles (Figure 3(f)) ranged between 2 µm2 and 140.2 µm2, whereas on glass, the axonal area was between 1.44 µm2 and 95.7 µm2.
We further characterized dendritic growth features on glass and bone. Dendrites extending from slices were imaged by immunofluorescence staining with anti-microtubule associated protein 2 (MAP2) and branches projected extensively from slices (Figure 4(a)), primarily toward the glass. Dendrites extending on the glass occasionally crossed to the powder layer, but ramification on the bone was not as extensive as on the glass (Figure 4(b)). A minority of the explants grew directly on the powder (Figure 4(c) and (d)). In comparison with glass, bone produces a rate of dendritic extension of just 1 every 20 µm, whereas glass produced a rate of 4 every 20 µm (t[4] = 6.307, p = 0.0032; Figure 4(e)). Dendritic branches on glass elongated to a maximum length of 318 ± 42.66 µm, whereas on bone the branches were significantly shorter (250.7 ± 34.7 µm; t[18] = 2.559, p = 0.0197; Figure 4(f)).

Neurons sprout dendrites and migrate on bone powder. Inverted fluorescence images of neuronal cell bodies and dendrites (immunolabeled with anti-MAP2) in 13-day-old hippocampal slice culture (white dots) on bone powder: (a) extensions of dendrites from a slice (red arrows), (b) extending dendrites from the slice to the glass (red arrows) and diversion towards bone powder (yellow arrows). Green arrows: neurons that have migrated away from the slice, (c) dendrites extend from hippocampal slice to glass (red arrows) or directly onto bone powder (yellow arrows), (d) enlargement of dendritic extensions (red arrows) on bone powder, (e) mean counts of dendritic branches/20 µm of hippocampal slice border (p = 0.0032), and (f) mean length of dendritic extensions (p = 0.0197).
Immunofluorescence staining with anti-glial fibrillary acidic protein (GFAP) revealed that astrocytic behavior was influenced by contact with bone powder (Figure 5). After two weeks of cultivation, astrocytic processes extended from the tissue and grew on the bone particles without contacting the glass substrate (Figure 5(a) and (b)). In other instances, processes extended onto the glass (Figure 5(c)). When encountering bone particles, these processes either contacted and turned away or crossed over them (Figure 5(c)). Occasionally, cells migrated out of the tissue and were found on bone particles (Figure 5(a) and (d)) or on the glass (Figure 5(e)), with their processes transitioning between substrates. Quantification indicated that growth on bone affected astrocytic process ramification. Cells growing on glass had an average of 10 ± 5 primary and secondary processes per cell, whereas those on bone exhibited a 35.4% reduction (6 ± 1 processes per cell; Welch’s t-test: t[10.43] = 2.232, p = 0.0486; Figure 5(f)). Additionally, the length of processes was 19.2% shorter on bone than on glass (134.42 ± 55 μm vs 166.4 ± 42.3 μm, respectively; t[42] = 3.670, p = 0.0007; Figure 5(g)).
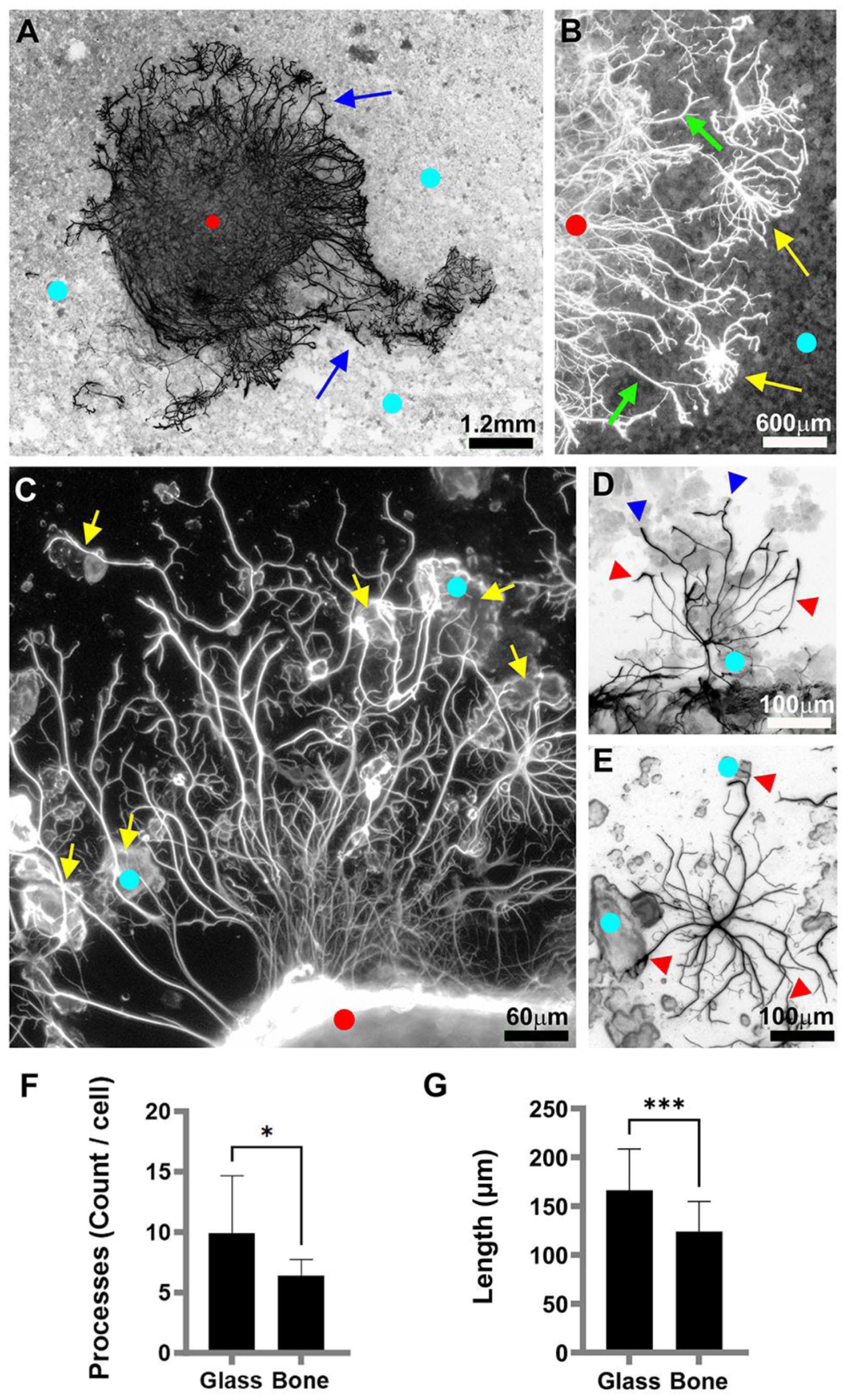
Astrocyte migration and outgrowth on bone powder. Fluorescence images (inverted, with exception to b and c) show astrocytes (immunolabeled with anti-GFAP) in a 13-day-old hippocampal slice culture (red dots). Light blue dots = bone powder coated regions: (a) astrocytes (blue arrows) extend from the slice onto bone coated regions, (b) enlarged portion of (a) shows both extension of processes from cells located within the slice (green arrows) and cells that have migrated outside the slice (yellow arrows), (c) the processes and astrocytes grow on uncoated glass and then divert to bone powder (yellow arrows), (d) representative astrocyte showing different process morphologies on uncoated glass (red arrowheads) and bone grains (blue arrowheads), (e) representative astrocyte with solitary processes growing on both the grains and the uncoated glass (red arrows), (f) mean process count/cell (p = 0.0486), and (g) mean process length (p = 0.0007).
Further, although the average area of astrocytes on bone and glass was similar (2950.6 ± 1928.7 µm2, 3361.8 ± 1424.5 µm2 for bone and glass, respectively; t[18] = 0.5423, p = 0.5943; Figure 6(a)), the morphology of the cells differed. Cellular circularity on glass was 0.01 ± 0.004, whereas on bone it was 3-fold greater at 0.03 ± 0.02 (Lognormal Welch’s t-test: t[15.48] = 3.545,p = 0.0028; Figure 6(b)). Additionally, on glass and bone, the average solidity of astrocyte morphology was 0.11 ± 0.03 versus 0.02 ± 0.086, respectively, constituting a 5.5-fold reduction on bone tissue (Lognormal Welch’s t-test: t[13.88] = 2.851,p = 0.0129; Figure 6(c)). However, the roundness of the cells did not differ significantly between the two surface matrices (0.57 ± 0.16 vs 0.47 ± 0.24 for glass and bone, respectively; t[18] = 1.113, p = 0.2803; Figure 6(d)). Figure 6(e) shows that maximal migration distances of cells were similar on glass and bone (529.5 ± 202.7 µm vs 402.43 ± 164.7 µm for glass and bone, respectively; t[11] = 1.242, p = 0.2402). Furthermore, GFAP expression in astrocytes growing on glass or bone was also similar (7.18E+07 ± 3.59E+07 vs 8.96E+07 ± 3.59E+07 for glass and bone, respectively; t[18] = 1.106, p = 0.2832; Figure 6(f)).

Bone induces morphological changes in astrocytes. Morphometrical quantification of GFAP-labeled slice cultures on bone powder (13-day-old): (a–d) shape and dimensions of astrocytes (a: p = 0.5943; b: p = 0.0028; c: p = 0.0129; d: p = 0.2803), (e) mean maximal astrocyte migration distance (p = 0.2402), and (f) mean GFAP expression level (p = 0.2832).
Discussion
In the present investigation, we have established that astroglia and neurons respond to bone substrates. Neurons are known to respond to topographical cues that engage developmental pathways and influence cell growth and morphological differentiation, especially when cultured on micropatterned ceramics that mimic extracellular matrix structures.68–70 In our study, axons displayed a marked increase in medium neurofilament (NFM) expression and a notable expansion of axonal diameter on bone grains. In contrast with the enhanced axonal features, neuronal dendrites did not exhibit significant morphological differentiation on bone, whereas glass surfaces supported a more extensive dendritic arborization. These results indicate that bone may preferentially support axonal maturation and potentially the development of intercellular connections.
In the broader field of central nervous system (CNS) tissue engineering, soft hydrogel-based scaffolds such as collagen, hyaluronic acid, and PEG have been the dominant materials due to their ability to mimic the mechanical softness of brain tissue and support neurite extension.71–75 However, increasing interest has emerged in using rigid and structured substrates that serve roles such as anchoring, providing directional guidance, or facilitating interface formation between neural and non-neural tissues.70,76,77 Materials including silicon-based implants, nanostructured carbon scaffolds, and bioceramics like hydroxyapatite and bioactive glass have demonstrated promising results in modulating neural behavior and enhancing electrical integration and axonal guidance.78–83
As noted earlier, bone-derived scaffolds are well-established in regenerative medicine, particularly for orthopedic and dental applications.84,85 Decellularized bone matrices and autologous bone grafts with retained mineral content have been used successfully to support osteogenic, mesenchymal, and endothelial cell growth in both in vitro and in vivo settings. 86 Despite this widespread application, there is a notable lack of studies evaluating the potential of mineralized bone to support neural cells such as neurons or astrocytes. Given the unique interactive properties of neural cells with their environment, our study presents a novel investigation into the potential of mineralized bone as a functional substrate for neural growth and differentiation ex vivo.
To assess this, we employed crushed osseous mineralized bone powder as a naturally-derived, bioactive substrate. While distinct from standardized scaffold types like trabecular matrices or engineered cortical sections, this form provided a biologically-derived surface likely retaining some components of native bone. The irregular micro- and nanoscale topography of the bone grains likely has influence on cellular adhesion, cytoskeletal architecture, and morphogenesis as has been demonstrated in artificial osseous bone material scaffold mimics.87,88 The manual crushing process used in preparation introduces variability in grain size that may affect cellular responses but simultaneously mimics the heterogeneous nature of real bone surfaces. This heterogeneity allows for an initial evaluation of bone compatibility with neural cell types. Future work should investigate the granule size influences to examine whether topographical effects exert more control on cell migration and morphological differentiation.
The morphological responses of axons to the bone substrate were obvious, but their interpretation is complex. The emergence of cone-like axonal expansions may indicate complex remodeling processes in response to the substrate rigidity and topography. Such expansions may reflect adaptive responses involving branching, synapse formation, or exploratory outgrowth—hallmarks of neural plasticity.89,90 Supporting this, we observed elevated NFM expression, a cytoskeletal marker associated with axonal stability and growth. Conversely, these swellings resemble varicosities associated with impaired transport and axonal stress, commonly seen in neurodegenerative or traumatic conditions.91–94 This dual interpretation highlights the importance of discerning whether such morphological features are functionally beneficial or indicative of underlying stress.
Dendritic responses to bone were more modest. Although dendritic outgrowth was observed, it was significantly reduced compared with responses on glass. Dendritic arborization is critical for synaptic integration and information processing.95–97 The rigid, mineralized surface may limit cytoskeletal flexibility required for extensive branching or may lack certain adhesive or biochemical cues present on glass or synthetic substrates.98–100 Overall, while bone appears permissive for dendritic attachment, additional modification—such as ECM protein coatings or hybrid composites—may be necessary to fully support dendritic complexity. 101
Astrocytes exhibited pronounced responses to the bone substrate, underscoring their relevance to regenerative applications. Astrocytes are central players in neural repair, maintaining ionic balance, modulating synaptic activity, and releasing trophic factors. 102 In injury contexts such as TBI, astrocytes become reactive, adopting hypertrophic morphologies and secreting matrix components103,104—a response termed reactive astrogliosis. 105 Initially protective, this process can become maladaptive, leading to glial scar formation that hinders axonal regeneration. 106
Morphological analysis of astrocytes in our study revealed increased circularity and solidity, alongside decreased roundness, when grown on bone. These features are indicative of a shift from the highly branched, stellate shape of resting astrocytes to a more reactive or even chronically activated phenotype,103,107 although the consensus thinking currently advises establishment of additional standard astrocyte activation markers and development identified activated astrocyte subtypes. 104 Increased circularity suggests the retraction of peripheral processes, while greater solidity reflects compact spreading. Decreased roundness, despite smoother contours, points toward polarization or elongation—features linked to gliotic transformation. 104 These changes raise concern that bone substrates, while supportive of cell survival and migration, may promote prolonged or excessive astrocytic morphological changes and activation.
Nonetheless, the overall support of neurite outgrowth and astrocytic morphological changes observed in our cultures indicates a potential role for bone as a neural scaffold. These results align with previous findings from micropatterned poly-lysine-coated ceramics, described as ECM-mimicking “periosteum-like” surfaces, which have promoted neuronal differentiation and growth.108,109 Otomo et al. 110 demonstrated that microgrooved ceramics coated with poly-lactic acid enhanced neurite extension and astrocyte development compared to smooth surfaces. These studies lend further credibility to our findings and suggest that the structured, mineral-rich surface of bone may be similarly beneficial.
However, translation to clinical application requires substantial preclinical validation. Animal studies examining the efficacy of scaffolds in promoting migration, synaptic integration, and behavioral recovery remain limited. Regulatory restrictions in some regions complicate TBI model experimentation, prompting the development of ex vivo organotypic models that simulate TBI lesions. 111 Currently, most in vivo studies rely on synthetic polymers, peptides, or hydrogel systems such as PLGA, collagen-heparin bigels, or chitosan blends. 112 Though promising as screening platforms, these models lack the complexity of in vivo systems.
Mineralized bone presents a distinct alternative to currently studied neuroscaffolding materials. Its use in orthopedic and craniofacial contexts is already clinically validated, and its application in CNS settings—particularly at hard-soft tissue interfaces like the brain-skull boundary—holds great promise. Our data demonstrate that bone powder can influence neural morphology, supporting further investigation into structured constructs, sintered bone, or composite systems that combine mechanical integrity with tailored biological signaling.
Importantly, our ex vivo findings must be clearly distinguished from pathological scenarios involving traumatic bone intrusion into brain tissue. In penetrating TBI, bone fragments entering the parenchyma can induce inflammation, scarring, and infection due to blood-brain barrier disruption and immune activation.65,113 In contrast, the controlled application of sterile, processed bone avoids these risks, though comprehensive safety assessments remain critical. Scaffold-associated concerns include micro-motion, immune reactivity, and microbial colonization. Addressing these issues through surface treatments, antimicrobial additives, and biomechanical optimization will be essential.
Furthermore, the apparent tendency of bone to induce chronic astrocytic and possibly neuronal reactivity requires careful monitoring. Chronic gliosis may compromise integration and limit long-term regenerative outcomes. Therefore, material optimization should aim to preserve the bioactivity of bone while minimizing cues that elicit prolonged reactivity. In vivo models assessing tissue compatibility, remodeling dynamics, and functional recovery are crucial next steps.
Autologous bone use presents an attractive clinical path, particularly for TBI repair. Using bone harvested from the patient avoids immune rejection, minimizes regulatory barriers, and supports biological integration. Compared with synthetic materials or xenografts, autografts are inherently biocompatible, non-immunogenic, and non-tumorigenic—key requirements for CNS application. If prepared under sterile, controlled conditions, bone could be reintroduced during surgical repair, serving as both a protective structure and regenerative substrate.
To promote successful repair, neurite outgrowth must outpace glial activation to avoid the formation of inhibitory scars. 114 Achieving this balance will require scaffold designs that enhance early neuronal activity while suppressing prolonged gliosis. Mineralized bone, particularly when combined with neural-permissive coatings or co-scaffolds, may meet this challenge.
Conclusions
This study demonstrates that mineralized bone supports neural cell adhesion, migration, and morphological differentiation, with differential effects on axons, dendrites, and astrocytes. Axonal elongation, thickening, and NFM upregulation were observed, alongside limited dendritic arborization and morphological signs of astrocyte activation. These findings suggest that bone scaffolds may enhance neural regeneration but must be carefully engineered to balance structural support with cellular homeostasis. Ongoing studies will focus on functional integration and behavioral recovery in in vivo models of CNS trauma.
Footnotes
Abbreviations
DAPI: 4′,6-diamidino-2-phenylindole
MAP2: Microtubule-associated protein 2
NFM: Neurofilament-M
GFAP: Glial fibrillary acidic protein
DDW: Double-distilled Milli-Q (0.22 µm) water
Author contributions
Orly Weiss: Investigation, data curation, formal analysis, writing—original draft preparation, visualization. Michael Kirby: Writing—original draft preparation, writing—review and editing, visualization. Danny Baranes: Conceptualization, resources, supervision, project administration, funding acquisition.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding was provided by the Fund for Research Collaboration from Ariel university
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
Source data is available on request from the corresponding author.*