
Abstract
The long-term success of intra-arterial stenting remains limited by in-stent restenosis (ISR). Transforming growth factor–β1 (TGF–β1) can inhibit smooth muscle cell (SMC) proliferation and migration and convert SMCs into extracellular matrix (ECM)–synthesizing cells. Here, we evaluate the effects of stent-based delivery of TGF-β1 on ISR in a rabbit model. Channeled stents loaded with TGF-β1 or control microspheres were deployed in rabbit aortas. Stented aortas were harvested at 7 and 28 d and evaluated for Ki-67–positive cells, collagenous ECM production, and intima-to-media (I/M) ratio. At 7 d, the TGF-β1 group exhibited fewer Ki-67–positive cells were found for the TGF-β1 group (17.87 ± 2.18 cells per mm2) relative to control (25.07 ± 2.65 cells per mm2, p = 0.04), but increased collagen content (31.4 ± 2.5 percentage area) compared with control (29.3 ± 1.2 percentage area, p = 0.019). The I/M ratio in the TGF-β1 group was reduced by 50% and 9.1% versus control at 7 d (0.13 ± 0.02 vs. 0.26 ± 0.02, p = 0.0001) and 28 d (1.80 ± 0.05 vs. 1.98 ± 0.08, p = 0.0038), respectively. Stent-based controlled release of TGF-β1 limits ISR and is associated with inhibition of SMC proliferation but an increase in ECM production.
Introduction
Angioplasty and stent placement are mainstays of endovascular interventions for coronary, peripheral, and carotid steno-occlusive arteriovascular diseases. However, restenosis, where the vascular lumen renarrows as a reparative response to vascular injury,1,2 limits the long-term efficacy of these revascularization procedures. The mechanisms of restenosis include neointimal hyperplasia, elastic recoil, and negative vascular remodeling.2,3 In the coronary arteries, restenosis following angioplasty occurs in 30% to 50% of patients within 6 mo. 4 Although placement of bare metal stents (BMS) provides a scaffold that prevents elastic recoil and negative remodeling, in-stent restenosis (ISR) occurs in 15% to 35% and is characterized primarily by neointimal hyperplasia.3,5–7 Drug-eluting stents (DES) designed to directly address ISR have further reduced restenosis rates to less than 10% but are associated with an increased risk of late in-stent thrombosis, and the long-term effects after complete elution of the therapeutic agents remain unclear.8–10 To further develop strategies to prevent ISR, continued elucidation of the pathobiologic processes underlying neointima formation following placement of BMS remains warranted.
The development of neointimal hyperplasia in the weeks to months after initial arterial stent deployment is temporally characterized by an early proliferative phase in which vascular smooth muscle cells (SMC) migrate from the media to the neointima and replicate.1,11–13 This process then transitions to a late synthetic phase dominated by extracellular matrix (ECM) deposition.14,15 A number of growth factors and cytokines play significant roles in regulating this complex process. 3
Transforming growth factor–β1 (TGF-β1) is a pleiotropic growth factor that modulates a wide variety of cellular processes, including cell proliferation, ECM production, and other pathways involved in wound healing.16,17 Its role in ISR is, however, controversial, with many studies implicating TGF-β1 as a promoter of neointimal hyperplasia,18–21 whereas others suggest that it may have protective effects.17,22 Central to this controversy is its bifunctional effect on SMCs: TGF-β1 can both induce and inhibit SMC proliferation and migration.19,23–26 It has been suggested that whether exogenous application or induced expression of TGF-β1 stimulates or inhibits SMC proliferation and migration depends on several factors, including the presence of other growth factors, 18 dose and duration,24,27 and whether TGF-β1 reaches the adventitia. 22 More well recognized is the significant role of TGF-β1 in regulating ECM production,14,28,29 as it has been shown to induce phenotypic conversion of proliferating vascular SMCs into ECM-depositing cells.30,31
In this study, we used a previously characterized DES platform 32 to locally deliver TGF-β1 in a rabbit model of ISR to investigate its effects on SMC proliferation, ECM production, and neointima formation.
Materials and Methods
Stent Design
The stent platform is based on the Palmaz-Schatz coronary stent. The stents are 1.6 mm in diameter and 10 mm in length, with 10 longitudinal struts evenly spaced circumferentially and balloon expandable up to 5 mm in diameter. The basic slotted tube design was modified by electro-discharge machining (Norman-Nobel, Cleveland, OH) channels 80 µm wide by 60 µm deep along the length of the abluminal (i.e., toward the wall) side of each strut. The channels were machined from end to end to allow drug delivery both to the wall and to the tissue surrounding the stent ends. Biodegradable microspheres, which store and release therapeutic agents, were then mechanically loaded into these channels and covalently anchored, as previously described. 32
Microsphere Preparation
Biodegradable poly(lactic-co-glycolic-acid)/polyethylene glycol (PLGA/PEG) microspheres were prepared through a modified version of previously described techniques.33–36 A mixture of 75:25 PLGA (Polysciences, Warrington, PA) and PEG-8000 (Fisher Scientific, Fair Lawn, NJ) was employed with the double emulsion technique to generate a final microsphere diameter of 10 µm and a degradation and drug release time of approximately 4 wk, as characterized previously.33–36 In addition, a pH buffer of 7.4 was incorporated to limit local pH changes, stabilize incorporated TGF-β1, and render the microspheres more biocompatible. 37 During the microsphere manufacturing process, recombinant TGF−β1 (Calbiochem, San Diego, CA; 0.0083 µg per week per stent) was added as an aqueous solution to the polymer solution. Control microspheres containing only polymer and buffer were also prepared.
Stent Loading
Channeled stents were pretreated with 2% 3-methacryloxypropyltrimethoxysilane (Polysciences) in 75% ethanol and then heat cured for 60 min at 105 °C to form a metal oxide-methoxysilane-monomethacrylate link to the metal-oxide layer of the stent surface. Stent channels were then manually loaded with control or TGF-β1–containing microspheres. Next, the stents were temporarily ensheathed in a silicone tube, and a gel (20% mixture of PEG-dimethacrylate [Polysciences] and PEG in a ratio of 3:2) was introduced to fill unoccupied spaces within stent channels and subsequently polymerized to form gel-stent links by exposure to UV-A overnight. The stents were then loaded on 5 mm × 2 cm angioplasty balloons (Jupiter; Cordis, Miami, FL) and lyophilized overnight.
In Vivo Stent Implantation
Age-matched adult male New Zealand White rabbits, weighing 3.8 to 4.2 kg, were used in accordance with National Institutes of Health and institutional guidelines (n = 8 per group; for each group, 5 for 7 d time point, 3 for 28 d time point). Under general anesthesia, the common femoral artery was surgically exposed, an arteriotomy was made, and a 5 Fr sheath was placed. With fluoroscopic guidance, aseptically prepared channeled stents filled with control or TGF-β1 microspheres were deployed in the infrarenal abdominal aorta. The proceduralists were blinded to the contents of the microspheres loaded onto the stents. Stents were then postdilated with a 5 mm angioplasty balloon at 8 atm to a final lumen size of 125% above baseline. Care was taken to ensure that no branches were present within the stent segment. Pre- and postdeployment digital subtraction angiograms were recorded for both control and TGF-β1 treatment groups. The rabbits were fed a 0.25% cholesterol diet post intervention.
Tissue Harvesting and Preparation
At 7 or 28 d after stent placement, animals underwent total-body perfusion-fixation with 2 L of 2% neutral buffered formalin after 2 L of normal saline was infused, as previously described. 38 The 7 d aortic specimens were excised longitudinally, and stents were removed. Each specimen underwent postfixation in 10% neutral buffered formalin for 12 to 16 h and were then embedded in paraffin for light microscopic and morphologic analysis. The 28 d aortic specimens were postfixed with stents in situ in 10% neutral buffered formalin and then embedded in a medium (PolyBed; Polysciences) for light microscopy and morphologic analysis.
Evaluation of Neointima: Intima-to-Media Ratios
For the 7 d aortic paraffin-embedded specimens, three serial 5 µm thick cross sections were obtained. An elastica van Gieson-Masson trichrome or elastica van Gieson-hematoxylin double stain was performed on each section. 36 For the 28 d specimens, the aortic segment was subdivided into five equal segments so that the full length of the stent would be represented in subsequent analyses. One section was obtained from each segment, and all were stained by modified Verhoff elastic staining. 36 High-resolution digital images at 100× magnification were then obtained of the stained 7 and 28 d specimens using a SPOT true-color camera (Diagnostic Instruments, Silver Spring, MD) with a microscope (Nikon E600 with Plan Apochromat Lenses; Nikon, Melville, NY). Using ImagePro Plus software (Media Cybernetics, Silver Spring, MD), the cross-sectional area of the intima and the media were determined by blinded observers, and the ratio of intima area to media area was subsequently tabulated for each specimen.
Evaluation of SMC Proliferation
Immunohistochemistry for Ki-67 was employed as a marker of SMC proliferation. Cross sections from 7 d aortic specimens (three sections per animal) were incubated at 37 °C overnight with a primary monoclonal antibody to Ki-67 (clone Mib-1; dilution 1:100; Dako, Carpinteria, CA) followed by incubation with appropriate secondary antibodies and chromogen. The number of Ki-67–positive cells per cross section was subsequently tabulated for each by a blinded observer. To eliminate error caused by variations in artery size, the number of proliferating cells was divided by the corresponding cross-sectional area for each and reported as the density of proliferating cells.
Evaluation of ECM Production
Quantification of vessel wall collagen content was undertaken at the 7 d time point using Masson Trichrome staining with or without Verhoeff von Gieson counterstaining of elastin to measure the production of ECM by SMCs. Digital microscopic images obtained as above were analyzed with ImagePro Plus software. Total collagen content was determined by converting acquired cross-sectional images to a floating point and thresholding at 0 to 35. The processed images were then analyzed to determine the percentage of collagen-positive pixels and confirmed by single-stained Masson Trichrome results. The results were divided by the total image area in pixels prior to processing and reported as cross-sectional percentage collagen for each group. All collagen measurements were determined by a blinded observer.
Statistical Analysis
All data were expressed as mean ± standard error. For all experiments, mean and standard error were determined for each group, with statistical comparisons made using one-factor analysis of variance repeated measures with significance evaluated at 95% using Statview (SAS Institute, Cary, NC) and SPSS 6.1 (Prentice Hall, Upper Saddle River, NJ).
Results and Discussion
TGF-β1 was first implicated to play a significant role in ISR by correlative studies that showed increased levels of TGF-β1 in human restenotic lesions.14,39 However, whether TGF-β1 contributes to or limits neointima formation remains controversial. At the core of this controversy is the ability of TGF-β1 to both promote and inhibit SMC proliferation and migration. Although several studies have applied exogenous delivery of TGF-β1 or TGF-β1 gene transfer to investigate its role in ISR development,22,40,41 the overall end effect is thought to be dependent on several factors, including dose, duration, and where the treatment reaches spatially within the layers of the vessel wall.22,24,27 Hwang et al. 24 showed that a higher dose of exogenous TGF-β1 inhibited SMC growth, whereas lower doses stimulated proliferation. The timing of treatment in the temporal sequence of neointima formation is likely another important factor.
Here, we employed a previously validated stent-based drug delivery platform for controlled and local delivery of TGF-β1 in a rabbit model of ISR. 32 Recombinant TGF-β1 was encapsulated in biodegradable PLGA/PEG microspheres that were then loaded into channels created along the abluminal surface of coronary stents. We have previously shown that therapeutic agents loaded within PLGA/PEG microspheres are released with an initial bolus over 1 wk and then followed by slowly decreasing but sustained release over the next 3 wk.36,42 We have also previously demonstrated that the buffered polymer system employed here does not enhance local inflammation. 32 Using this stent platform, which serves to simultaneously deliver drugs and model ISR, channeled stents loaded with TGF-β1–eluting microspheres or control microspheres were implanted in the abdominal aortas of New Zealand White rabbits that were then fed a high-cholesterol diet. Stented aortic segments were excised and evaluated at 7 and 28 d after stent placement to evaluate early and late effects.
SMC Proliferation
The impact of stent-based release of TGF-β1 on vascular SMC proliferation was examined relative to controls at 7 d (n = 5 per group) by staining for Ki-67, a cellular marker for proliferation. As summarized in Figure 1 , stent-based local release of TGF-β1 resulted in significantly fewer Ki-67–positive cells (17.87 ± 2.18 cells per mm2) relative to control (25.07 ± 2.65 cells per mm2, p = 0.04).
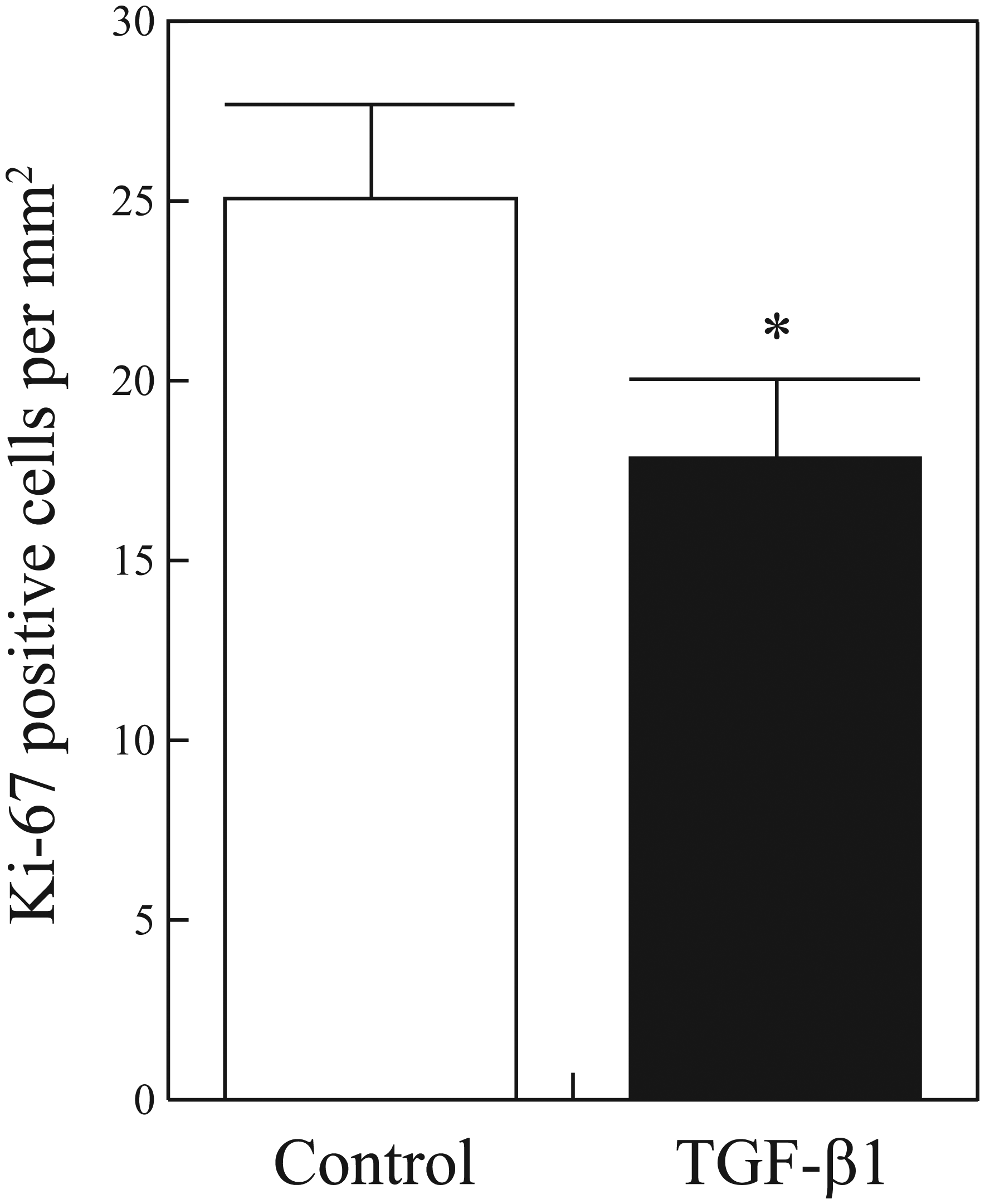
Density of proliferating cells after stent placement. Number of Ki-67–positive cells per mm2 7 d after arterial stenting and release of control or TGF-β1. Values represent mean ± standard error. *p < 0.05.
These results imply that a sufficient dose of TGF-β1 was delivered to inhibit, and not stimulate, SMC proliferation. The sustained delivery of TGF-β1 over the 7 d of implantation likely also contributed to the antiproliferative effects. The specific dose threshold above which exogenous TGF-β1 delivery inhibits SMC proliferation in vivo is unknown.
Collagen Production
Relative collagen content was evaluated as a proxy for overall ECM production. The effect of stent-based local delivery of TGF-β1 on collagen deposition in the neointima was evaluated at 7 d (n = 5 per group). As summarized in Figure 2 , collagen content was significantly increased in the TGF-β1 treatment group (31.4 ± 2.5 percentage total area) relative to the control group (29.3 ± 1.2 percentage total area, p = 0.019).
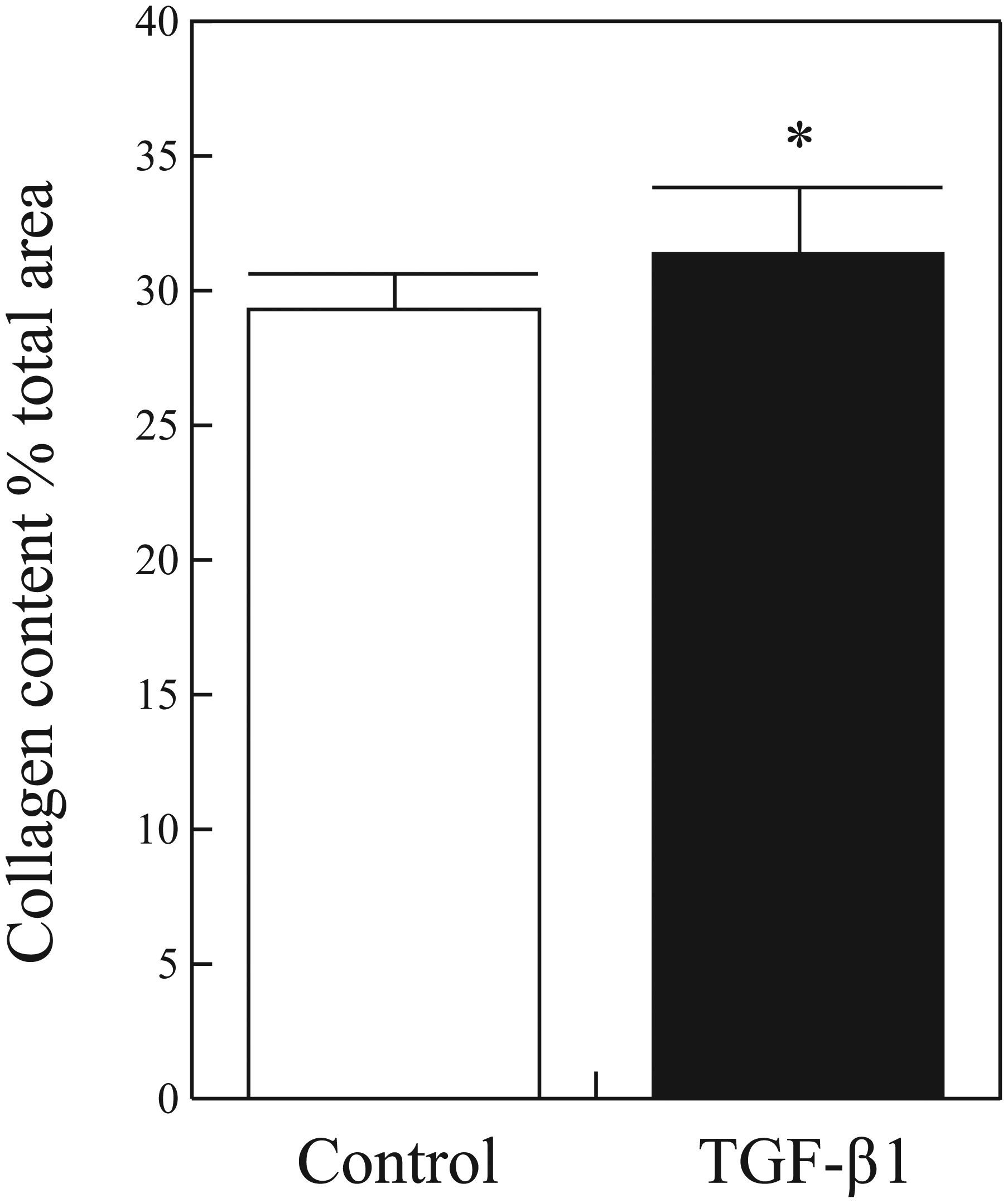
Collagen content after stent placement. Collagen content as a percentage of total cross-sectional area 7 d after in vivo stent deployment and release of control or TGF-β1. Values represent mean ± standard error. *p < 0.05.
With inhibition of SMC proliferation, SMCs are converted into ECM-producing cells.30,31 The increased collagen content and decreased SMC proliferation observed with the TGF-β1 treatment group confirm this transition, which was induced with local TGF-β1 delivery over 7 d. It is unknown whether other processes, such as local recruitment and adluminal migration of other ECM-producing cells, may also play a role in this observed effect. Of note, the timing of increased ECM deposition relative to controls is earlier than usual in the temporal sequence of neointima formation. Neointimal lesions are typically minimally cellular and consist mostly of ECM only after 1 to 2 mo from initial vascular injury,14,43 not after 7 d. Collagen accounts for a significant component of neointimal ECM. 43 With stent-based delivery of TGF-β1, an ECM-dominant neointimal morphology was seen only 7 d after initial stent placement.
Neointima Formation
Although inhibition of SMC proliferation limits ISR, ECM production and deposition contribute to neointima volume. The overall end effect of stent-based TGF-β1 release on ISR relative to control was evaluated at both 7 and 28 d, with results summarized in Figure 3 . At 7 d ( Fig. 3a ), the intima-to-media (I/M) ratio of the TGF-β1 treatment group (0.13 ± 0.02) was half that of the control group (0.26 ± 0.02, p = 0.0001). At 28 d ( Fig. 3b ), the I/M ratio was reduced by 9.1% in the TGF-β1–treated group (1.80 ± 0.05) relative to control (1.98 ± 0.08, p = 0.0038). Stent-based local treatment with TGF-β1 thus resulted in a significant reduction of in-stent neointima formation relative to control at both 7 and 28 d. There was no evidence of intraluminal thrombi in either group or time point.
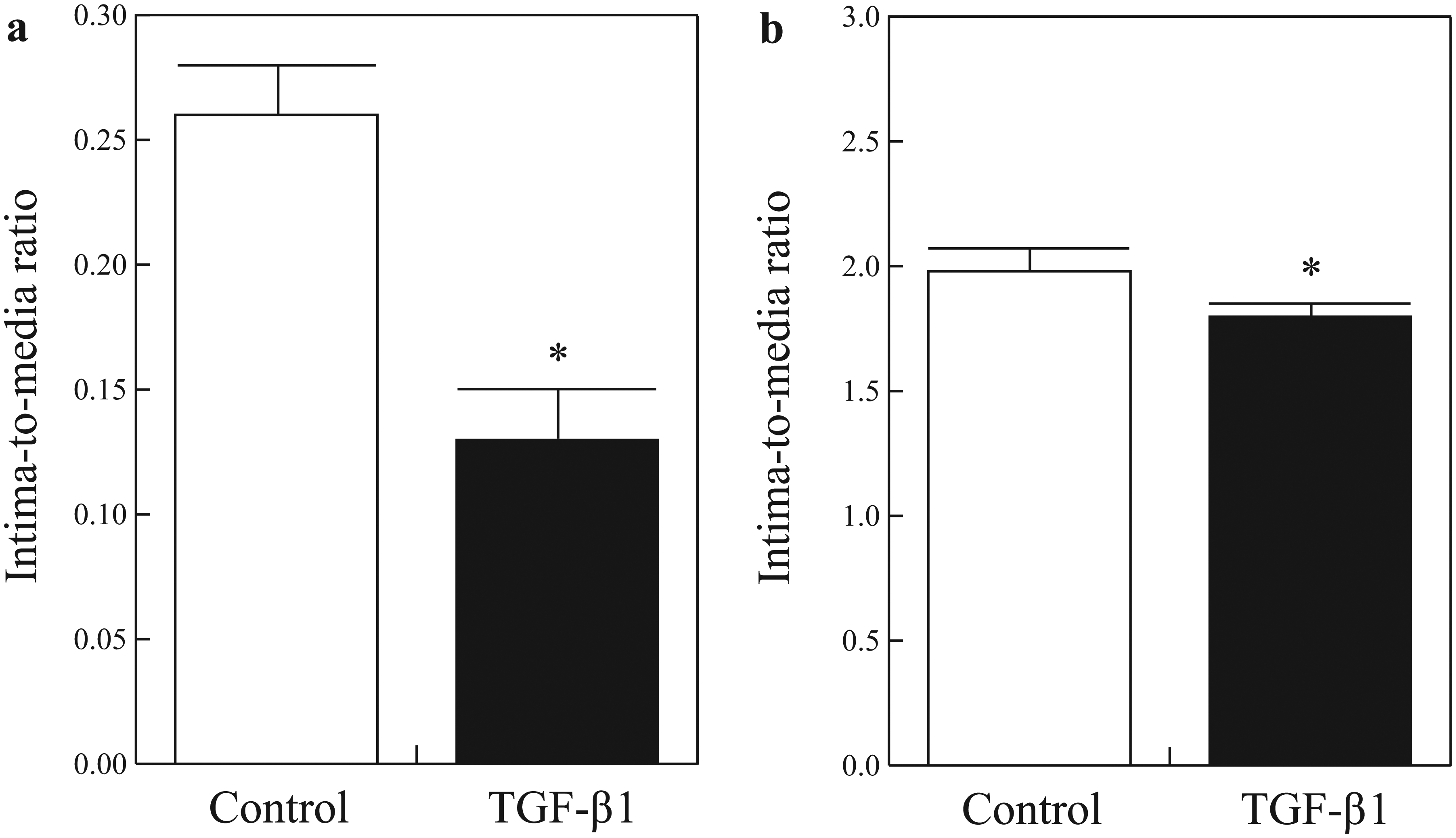
Intima-to-media ratio after stent placement. The ratio of intima area to media area at (
Although we found statistically significant reductions in neointima size by stent-based TGF-β1 release at both early and late time points, the magnitude of reduction was more remarkable at 7 d than at 28 d. Because the neointima is composed mostly of ECM in the late phase, whereas early neointima is composed almost entirely of proliferating cells, the Ki67 and collagen content data suggest that the more significant relative reduction in neointima size at 7 d reflects early termination of the proliferative phase of neointima formation with concomitant early transition into the ECM-synthesizing phase. Assuming that stent-based TGF-β1 local delivery inhibited both SMC proliferation and migration, this would result in fewer cells in the neointima to produce and deposit collagen and other ECM proteins and thus less neointima. However, it is possible that the attenuated but continued release of exogenous TGF-β1 between days 7 and 28 further stimulated ECM production relative to the control group, resulting in an overall smaller difference between the two groups at 28 d.
Although the implantation of stents that elute antiproliferative drugs has significantly reduced ISR rates in the coronary arteries, ISR is a complex response to injury involving several cellular processes. An antiproliferative approach alone may be insufficient to resolve ISR. Here, we investigated TGF-β1 as a potential key factor in neointima formation. A stent-based drug delivery system was employed to study the effects of local and sustained release of exogenous TGF-β1 on arterial ISR. In this model system, TGF-β1 treatment resulted in decreased cell proliferation but increased collagenous ECM. Exogenous TGF-β1 delivery thus functioned as a phenotypic switch, converting SMCs that would otherwise proliferate and migrate into the neointima after injury into cells that produce ECM proteins. In doing so, the temporal sequence of neointima formation was accelerated by early termination of the proliferative phase and immediate transition into the ECM synthesis phase. The overall end effect observed was reduced neointima formation at both early and late time points.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.