
Abstract
The PNPLA3 single nucleotide polymorphism, rs738409, is the strongest known genetic risk factor for metabolic dysfunction-associated steatotic liver disease; thus, targeting the minor allele with a GalNAc-conjugated siRNA is an attractive strategy to treat patients carrying the genetic variant. To enable translational safety assessment of a GalNAc-conjugated siRNA that specifically targets the rs738409 sequence of PNPLA3, a transgenic human PNPLA3I148M knock-in mouse (huPNPLA3I148M) was utilized. This model showed no significant genotype-related phenotypic differences to wild-type mice in a phenotype characterization study when maintained on standard rodent chow. Additionally, a repeat-dose toxicology study using a GalNAc-conjugated siRNA specific for rs738409 resulted in comparable findings between genotypes (i.e., liver enzyme and histopathology changes), indicating the findings were due to the siRNA therapeutic and not a result of target knockdown in huPNPLA3I148M mice. Overall, these data demonstrate the huPNPLA3I148M mouse is suitable for repeat-dose toxicology studies, suggesting this approach could be applied to other siRNA programs lacking a pharmacologically relevant nonclinical species to support translational safety assessments during drug development.
Introduction
The missense single nucleotide polymorphism (SNP) rs738409 (I148M) of the patatin-like phospholipase domain containing 3 gene (PNPLA3I148M) is the most powerful common genetic driver across the metabolic dysfunction-associated steatotic liver disease (MASLD) spectrum.1–3 Roughly half of all people with MASLD carry at least one PNPLA3I148M allele, and with global MASLD prevalence now above 30% of adults, the variant has become a prime therapeutic target.3,4 Furthermore, rs738409 is in near-perfect linkage disequilibrium (r2 ≈ 0.99) with the synonymous rs738408 variant three bases downstream, forming a distinctive minor-allele haplotype that can be exploited for allele-selective RNA-interference (RNAi). 5
Functional work increasingly supports the idea that the PNPLA3I148M variant is a neomorphic mutation that acquires novel pathogenic activities, rather than a simple loss- or gain-of-lipase variant, underscoring the value of allele-specific silencing strategies.6–8 Thus, allele-specific small interfering ribonucleic acids (siRNAs) that silence only the PNPLA3I148M transcript, while sparing the PNPLA3 reference allele (whose endogenous role remains unclear), offers a precision medicine-based approach for both homozygous and heterozygous carriers with reduced on-target safety risk. 9
The six-nucleotide haplotype encompassing rs738409/rs738408 is human-specific, allele-specific siRNAs targeted to this sequence are inactive in standard nonclinical species (i.e., mouse, rat, dog, and non-human primate). Therefore, alternative development strategies such as surrogate oligonucleotides that are pharmacologically active in the selected species or genetically modified models such as transgenic knock-in (KI) mice, which express the human target gene, need to be considered. In addition, new approach methodologies (NAMs) may be considered,10,11 but current drug development guidance under which siRNAs are regulated requires general toxicity assessment in both rodent and non-rodent species. 12
To support translational safety assessment and drug development of an siRNA targeting the rs738409/rs738408 sequence, we generated a human PNPLA3I148M KI mouse (huPNPLA3I148M). This KI mouse, like the original model described by Smagris et al., 13 is phenotypically normal on standard rodent chow but can develop steatosis and progress to steatohepatitis and early fibrosis on a Western diet (high fat and high sugar). Here we show that on a standard rodent diet the huPNPLA3I148M KI mouse provides a pharmacologically relevant species for siRNA safety assessment.
Materials and Methods
Synthesis and In Vitro Characterization of siRNA-1
siRNA-1 sequence identification, synthesis, and N-acetyl galactosamine (GalNAc) conjugation were performed using methods previously described by Murray et al., 2021. 9
Characterization of siRNA-1 was performed in Hep3B cells (homozygous for the human PNPLA3 reference allele) and HepG2 cells (homozygous for the human PNPLA3I148M variant) using the QuantiGene® ViewRNA HC Screening Assay (Thermo Fisher Scientific). siRNA-1 with a 10-point concentration range was reverse transfected into Hep3B or HepG2 cells with Lipofectamine RNAiMAX. After 72 h incubation, plates were processed on Amgen’s automated platform according to manufactural protocol. Images captured on the Opera Phenix (PerkinElmer) were analyzed in Columbus software to quantify mean PNPLA3 messenger ribonucleic acid (mRNA) spot counts per cell. Counts were normalized to phosphate buffered saline controls (100%) and “no probe” wells (0%). Dose-response curves were fitted with a four-parameter sigmoidal model in Genedata Screener.
Generation of huPNPLA3I148M homozygous knock-in mouse
The huPNPLA3I148M homozygous (HO; huPNPLA3I148M/huPNPLA3I148M) KI mouse model was generated on a wild-type (WT) C57BL/6J background using genetic editing at Inotiv (USA; https://www.inotiv.com/genetically-engineered-models-and-services). In brief, the donor deoxyribonucleic acid (DNA) containing the human PNPLA3 coding sequence with rs738408 and rs738409 was targeted to the start codon of the mouse Pnpla3 sequence, resulting in expression of the donor DNA under normal control of the mouse promoter while eliminating expression of the mouse Pnpla3. Four founder animals were verified to be positive for the targeted integration, and two were selected for backcrossing onto WT mice to generate the first filial generation and then backcrossed for at least 10 generations before use in translational safety studies. The genotyping strategy and results used to confirm successful donor integration and screen for random integration are provided in Supplementary Data S1.
In vivo studies
All animal experimental protocols involving the care and use of animals in this article were reviewed and approved by the Charles River Laboratories Institutional Care and Use Committee and were conducted in accordance with the guidelines set by the Association for Assessment and Accreditation of Laboratory Animal Care.
Phenotyping study
This study phenotypically characterized 21–22-week-old male and female huPNPLA3I148M heterozygous (HE; mouse Pnpla3/huPNPLA3I148M) and HO (huPNPLA3I148M/huPNPLA3I148M) KI mice in relation to WT (mouse Pnpla3/Pnpla3) littermates through clinical observations, body weight evaluation, neurobehavioral assessment (functional observational battery [FOB]) including quantitative motor activity, clinical pathology, organ weights, and gross and histopathology evaluations. All mice were maintained on a standard rodent chow of Lab Diet Certified CR Rodent Diet 5CR4 and housed in polycarbonate cages containing appropriate bedding equipped with an automatic watering valve. Full methodology is available in Supplementary Data S2.
Non-Good Laboratory Practice (GLP) 29-day repeat-dose toxicology study
This study evaluated male and female huPNPLA3I148M HO KI or WT mice (7 weeks of age) in a non-GLP repeat-dose toxicology study using a GalNAc-conjugated siRNA (siRNA-1). All mice were maintained on a standard rodent chow of Lab Diet Certified CR Rodent Diet 5CR4 and housed in polycarbonate cages containing appropriate bedding equipped with an automatic watering valve. Following randomization, animals were group housed (up to three animals of the same sex and same dosing group together). Prior to randomization, animals may have been individually housed. siRNA-1 was administered by subcutaneous injection to huPNPLA3I148M HO KI or WT mice (n = 6/sex) as 3 doses of (control), 30, or 300 mg/kg on days 1, 15, and 29. The study evaluated clinical observations, body weights, neurobehavioral assessment (FOB) including quantitative motor activity, ophthalmology, clinical pathology, organ weights, gross findings, and histopathology. Serum and liver drug concentrations and liver huPNPLA3I148M mRNA expression knockdown were assessed in a separate cohort of female huPNPLA3I148M HO KI mice (n = 6). Detailed methodology is available in Supplementary Data S3 under the sections “Serum and Liver Bioanalysis” and “Liver mRNA Analysis.”
Results and Discussion
The phenotyping study was designed to evaluate genotype-related differences in huPNPLA3I148M HE KI mice that express both the mouse Pnpla3 allele and the huPNPLA3I148M variant, and HO KI mice that only express the huPNPLA3I148M variant. Both the huPNPLA3I148M HE and HO mice were compared to WT littermates. All mice were provided with standard rodent chow. By providing standard rodent chow, instead of a Western diet or another altered diet, the study avoids confounding genotype-mediated phenotypes with findings that may be a result of an altered diet. 14 Therefore, in the context of this study, the huPNPLA3I148M KI mice are simply a transgenic model rather than a disease model of MASLD used by others to interrogate PNPLA3I148M variant function and pathophysiology.13,15,16
There were no findings attributed to genotype during the phenotype characterization study (Fig. 1; full results in Supplementary Data S2). Identified differences among the genotypes were of small magnitude, considered incidental and unrelated to genotype, or attributed to biological variation. Moreover, no evidence of steatosis or further progression to steatohepatitis was observed given all three genotypes were on standard rodent chow and confirms that this model is phenotypically normal on standard rodent chow. Collectively, these results indicated that huPNPLA3I148M HE and HO KI mice do not exhibit unexpected physiological properties, were similar to WT littermates, and are appropriate for use in translational safety assessments when on standard rodent chow.
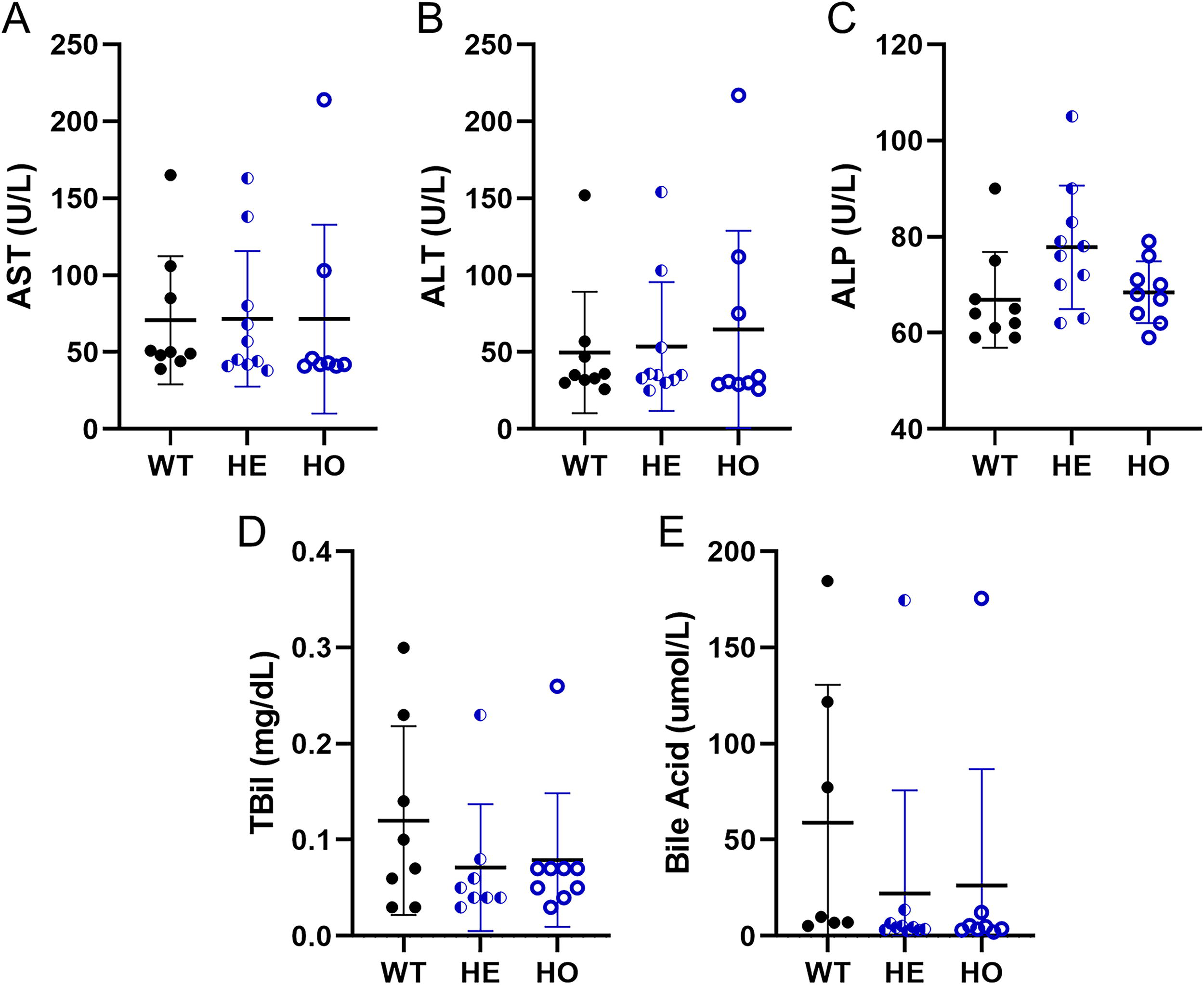
Select clinical chemistry parameters from the phenotyping study using huPNPLA3I148M homozygous, huPNPLA3I148M heterozygous, and wild-type mice. Clinical chemistry parameters were assessed from huPNPLA3I148M homozygous, huPNPLA3I148M heterozygous, and wild-type littermates prior to necropsy. No genotype-related differences were identified in
To further assess the utility of the huPNPLA3I148M HO KI mice for use in translational safety assessments, a GalNAc-conjugated siRNA (siRNA-1) that specifically silences the human sequence associated with the SNP rs738409 location was generated for use in a repeat-dose toxicology study. The specificity of siRNA-1 for the rs738409 sequence was demonstrated by robust mRNA knockdown of the PNPLA3I148M minor allele in HepG2 cells and the lack of PNPLA3 reference allele mRNA knockdown in Hep3B cells with IC50 values of 5.25 nM and >500 nM respectively (Fig. 2A).
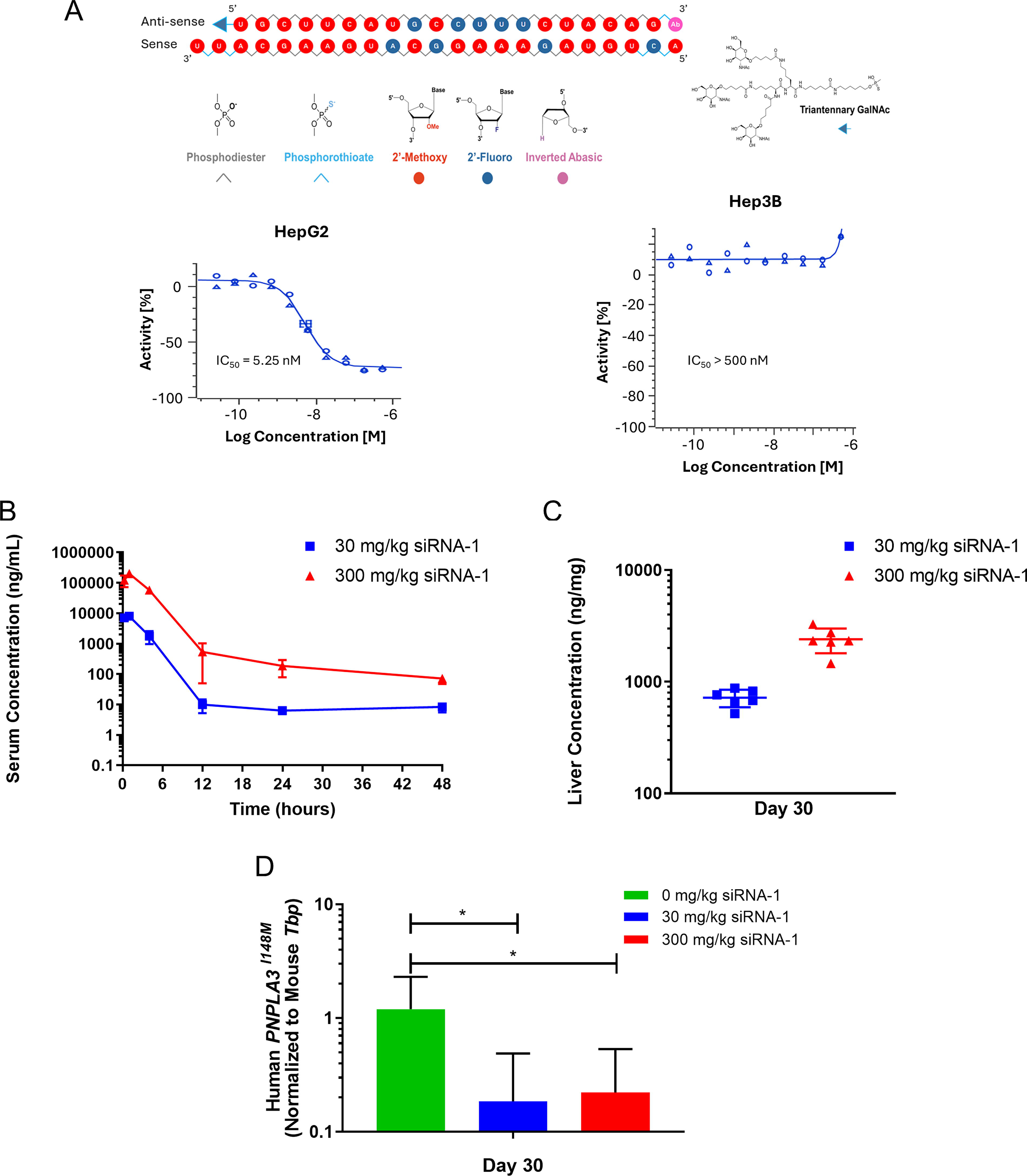
siRNA-1 and associated serum toxicokinetic profile, liver bioanalysis, and liver huPNPLA3I148M mRNA knockdown observed in non-GLP 29-day repeat-dose toxicology study using female huPNPLA3I148M homozygous knock-in mice. Female huPNPLA3I148M homozygous knock-in mice were administered 0 (control), 30, or 300 mg/kg siRNA-1 subcutaneously on days 1, 15, and 29.
siRNA-1 was administered subcutaneously on days 1, 15, and 29 at dose levels of 0 (control; no treatment), 30, or 300 mg/kg to both huPNPLA3I148M HO KI mice and WT mice. The mice were maintained on standard rodent chow for the duration of the study to enable assessment of siRNA-1-mediated effects and avoid potential confounding results associated with an altered diet and disease formation and progression.
On day 1, after the first subcutaneous administration of siRNA-1 to huPNPLA3I148M HO KI mice, the serum antisense concentration was 7.96 and 200 μg/mL for Cmax and 28.6 and 762 h*μg/mL for AUClast at 30 and 300 mg/kg, respectively (Fig. 2B). Average siRNA-1 antisense liver concentrations on day 30 at 30 and 300 mg/kg were 721 ng/mg and 2410 ng/mg, respectively (Fig. 2C). siRNA-1 antisense concentrations were below the limit of quantitation in the 0 mg/kg serum and liver samples. huPNPLA3I148M mRNA levels were significantly decreased in the liver compared to control animals at both 30 and 300 mg/kg by 0.185-fold (81.5% knockdown) and 0.221-fold (77.9% knockdown), respectively (Fig. 2D). siRNA-1 serum and liver concentrations were not assessed in WT mice nor was mouse Pnpla3 liver mRNA expression in this study.
During in-life study, there were no treatment-related unscheduled deaths, clinical or gross findings, ophthalmical findings, or changes in FOB parameters (Supplementary Data S3). In general, after study termination on day 29, there were comparable increases at 300 mg/kg in both WT and huPNPLA3I148M HO KI mice serum aspartate aminotransferase, alanine aminotransferase, alkaline phosphatase, total bilirubin, and bile acids levels (Fig. 3). Similar increases in absolute and relative liver weights up to 12.2% were observed at 300 mg/kg in female WT and huPNPLA3I148M HO KI mice (Supplementary Data S3). Histopathology findings in both genotypes were limited to the liver and injection site, with dose-dependent vacuolation of macrophages in the subcutaneous tissue in both WT and huPNPLA3I148M HO KI mice (Supplementary Data S3). Liver findings were similar in both genotypes and included generally dose-dependent hepatocellular single-cell necrosis, increased mitoses, and vacuolation primarily affecting centrilobular hepatocytes (Supplementary Data S3). Hepatocellular vacuolation, single-cell necrosis with increased mitoses, and injection-site macrophage vacuolation are known platform-related effects of GalNAc-conjugated siRNA therapeutics in rodents. 17
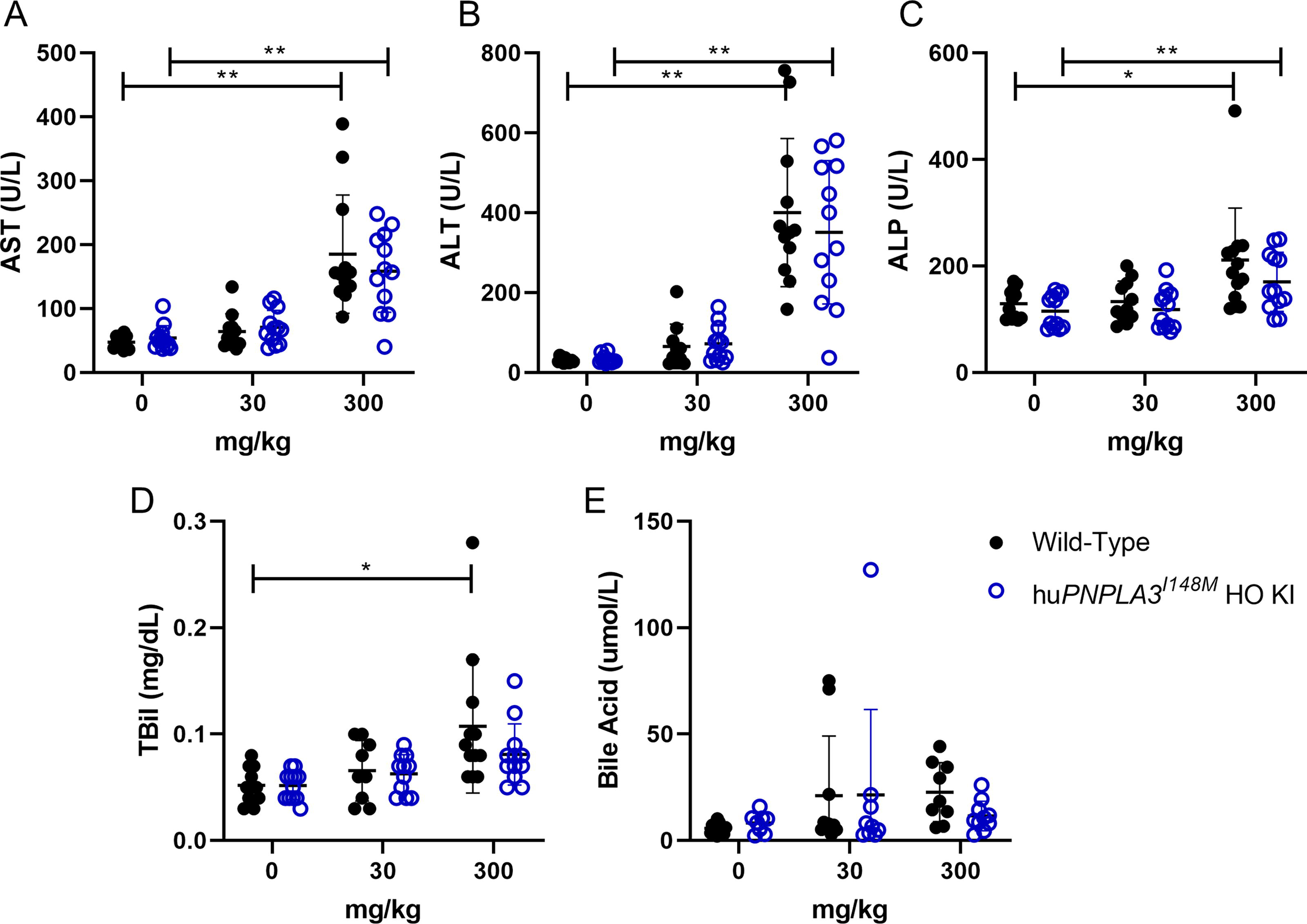
Select clinical chemistry parameters from the non-GLP 29-day repeat-dose toxicology study using huPNPLA3I148M homozygous knock-in and wild-type mice. Clinical chemistry parameters were assessed on day 30 (necropsy) after three subcutaneous dose administrations at either 0 (control), 30, or 300 mg/kg on days 1, 15, and 29 to female huPNPLA3I148M homozygous knock-in mice or wild-type mice. No genotype-related differences were identified in
Overall, the non-GLP 29-day repeat-dose toxicology study findings were generally similar between WT and huPNPLA3I148M HO KI mice treated with siRNA-1 indicating that the identified changes can be attributed to siRNA administration but not huPNPLA3I148M silencing. Moreover, the use of appropriately aged mice for repeat-dose toxicology studies (7 weeks of age) did not identify unique genotype findings consistent with the results of the phenotyping study.
Since the mice were maintained on a standard rodent chow rather than a Western diet or other altered diet, no findings of steatosis or similar progressed disease associated with PNPLA3I148M were identified in either huPNPLA3I148M HO KI mice or WT mice. Under conditions of the repeat-dose toxicology study, the putative benefits of PNPLA3I148M silencing in MASLD cannot be assessed since disease formation does not occur on standard rodent chow in this model. However, for the purposes of translational safety assessment and characterization of an siRNA for first-in-human trials this model on a standard chow diet enables a robust assessment without confounding data that may arise from disease models.
In summary, here we demonstrate that the huPNPLA3I148M HO KI mouse is a suitable model for repeat-dose translational safety assessments of siRNA therapeutics targeting the PNPLA3I148M sequence. Beyond general repeat-dose toxicology studies, this model may also be used for developmental and reproductive toxicity, safety pharmacology, or carcinogenicity assessments. On the other hand, the use of this model comes with certain limitations that warrant further discussion. First, while this model enables a clear assessment of siRNA-related effects without interference from background disease, it does not capture the full spectrum of phenotypes associated with MASLD, particularly in the context of diet-induced progression. Second, the translatability of toxicological findings from this mouse model to human patients remains to be fully validated, especially since rodent responses to oligonucleotides may differ in subtle ways from primate or human physiology. Third, generating and maintaining this KI line involves complex breeding strategies and may not be scalable or feasible across multiple targets or programs. Finally, the ethical considerations of introducing humanized SNPs into animal models for every variant of interest must be weighed against the scientific benefits, especially when in the future in vitro alternatives or NAMs may eventually provide equally robust translational safety data.
Footnotes
Acknowledgments
The authors would like to thank the CRL staff, scientists, pathologists, and study directors for their partnership and collaboration with the in vivo mouse studies described in this article. The authors would also like to thank Bill Siska for his interpretation and guidance with the clinical pathology data.
Author Disclosure Statement
All authors are current or former employees of Amgen Inc. and may own stock in Amgen, Inc.
Funding Information
Amgen, Inc. provided funding to perform the studies and generate the data described in the publication.
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.