
Abstract
The present study aimed to examine the immunomodulatory effect of ethanolic extract of Typhonium flagelliforme (Lodd) Blume in cyclophosphamide-treated rats. The immunomodulatory effects were determined by lymphocytes proliferation, phagocytic activity of macrophages, plasma cytokines of tumor necrosis factor-α, interleukin-1α, interleukin-10 levels, and killer T cells (CD8+ T cells) counts. The results showed that the administration of ethanolic extract of T flagelliforme reduced immunosupessive effect on lymphocyte proliferation, increase the number and phagocytic activity of macrophages in cyclophosphamide-treated rats. Moreover, the ethanolic extract of T flagelliforme also significantly (P < .05) improved the immune system activities especially the proliferation of CD8+T cells and reduced the suppressive effects on cytokines such as tumor necrosis factor-α and interleukin-1α. In conclusion, the ethanolic extract of T flagelliforme has immunomodulatory properties in cyclophosphamide-treated rats. The results suggest that T flagelliforme can reduce immunosuppresive effect caused by a chemotherapeutic agent.
Typhonium flagelliforme (Lodd) Blume, known as rodent tuber, is a medicinal herbal plant belongs to the family of Aracaceae. Rodent tuber is often used as a traditional remedy and widely used for alternative cancer therapy, including leukemia, by various ethnic population. 1 Tuber of T flagelliforme has been used as a health care supplement to cure many types of cancer, including breast, lung, rectum, liver, prostate, pancreas and cervical cancer and leukemia. 2
Some chemical constituents have been identified from T flagelliforme. The nonpolar extract was identified containing saturated hydrocarbons and aliphatic acids, while the ethyl acetate extract was found to contain aromatic fatty acids. 3,4 The previous study has shown that the hexane fraction of rodent tuber are cytotoxic against P388 murine leukemia. 5 Some fractions of hexane and dichloromethane were capable of inhibiting the growth of NCI-H23 lung cancer significantly and found that the fraction D/F21 was an active fraction for the inhibition of cancer growth. 6 The anticancer activity of T flagelliforme, however, had only been investigated in vitro on 2 murine cancer cell lines, P388 lymphocytic leukemia and a lymphoid cell line for which the cell line designation was not mentioned. 1,5 Both studies indicated that the nonpolar extracts of T flagelliforme possessed a significant antiproliferative activity. The study on rodent tuber as an anticancer agent has not been much done, although its use as an anticancer already quite extensive as products of traditional medicine or herbal medicine combine with chemotherapy for the treatment of cancer.
In recent years, natural products have become a vital part of complementary and alternative medicine. Natural products that show antitumor activity have been gaining popularity among those faced with the deleterious side effects of treatment using chemotherapeutic drugs or synthetic compounds for cancer treatment. The resulting immunosuppression induced by chemotherapeutic has been suggested to be a major contributing factor to exacerbate in cancer mortality. The recovery from immunodeficiency induced by chemotherapy remains a major clinical problem in cancer patients. Typically, hematological recovery is medically considered to have occurred when the number of neutrophils and platelets have been restored to the normal level. However, molecular modulations and cellular interactions in lymphoid and cytokine system are required for recovery of the immune system function and should also be considered. 7 Therefore, it is interesting to design therapeutic approaches to improving anticancer treatments in order to reduce the side effects.
In the present study, the immunomodulatory effects of ethanolic extract of T flagelliforme were evaluated in cyclophosphamide-treated rats, based on changes in lymphocyte proliferation, phagocytic activity of macrophage, serum level of tumor necrosis factor (TNF)-α, interleukin (IL)-1α, and IL-10, and the killer T cells (CD8+T cells) counts.
Materials and Methods
Plant Materials
Fresh tuber of T flagelliforme were collected from Tawangmangu Botanical Garden, Central Java of Indonesia and the taxonomist of the Botanical Garden authenticated the sample. A voucher specimen has been preserved in the Taxonomic Division of the Tawangmangu Botanical Garden of Indonesia.
Plant Preparation and Ethanolic Extraction
Fresh plants were harvested and washed thoroughly with running tap water and then with distilled water. The tubers were air dried and then oven dried under reduced temperature. The fully dried plants were powdered and weighed before cold maceration. The powder of T flagelliforme was macerated with ethanol 96%. The extracts were filtered before evaporated to dry under reduced pressure at 35°C with a rotary evaporator (Heidolph Instruments GmbH & Co, Schwabach, Germany). The ethanolic extract was further lyophilized by a freeze dryer VirTis BTK (SP Scientific, Gardiner, NY, USA).
Animals
Healthy Wistar rats were obtained from the Animal Experimental Unit, Animal Research and Development Centre, Gadjah Mada University, Yogyakarta. The animals were housed and maintained under the standard conditions of 12-hour light/dark cycle, 25° ± 2°C and 60% to 70% humidity and were fed with standard rat chow and water ad libitum. The experimental protocol was conducted in accordance with the Guideline for Care and Use of Animals Laboratory and approved by the Institutional Animal Ethics Committee of Universitas Gadjah Mada.
Experimental Animals
Thirty-six normal rats were divided randomly into 6 groups of 6 rats each group and used in the experiments. Group I served as normal rats received vehicle (carboxymethyl cellulose Na 0.5%), group II served as cyclophosphamide-treated rats (150 and 110 mg/kg body weight on day 1 and 4) and were administered carboxymethyl cellulose Na 0.5% orally once daily for 7 consecutive days; groups III, IV, and V received variation of doses of 250, 500, and 1000 mg/kg body weight of the ethanolic extract of T flagelliforme, respectively. The rats were administered ethanolic extract of T flagelliforme once daily for 7 consecutive days and cyclophosphamide doses of 150 mg/kg body weight on day 1 and 110 mg/kg body weight on day 4. Group VI received only ethanolic extract of T flagelliforme 1000 mg/kg body weight without cyclophosphamide treatment.
Proliferation of Lymphocytes
On day 7, the rats were sacrificed and a surgery was performed to take out the spleen and centrifuged (1200 rpm, 4°C, 10 minutes). The pellets obtained were suspended in 0.01 M Tris-buffered ammonium chloride, pH 7.65, and allowed to stand at room temperature for 2 minutes. Then, on the bottom of the tube was added fetal bovine serum (Gibco BRL, Grand Island, NY, USA). The suspension was then centrifuged (1200 rpm, 4°C, 5 minutes), and the supernatant was discarded. The pellets were washed with RPMI (Roswell Park Memorial Institute) medium twice and centrifuged. Supernatant was discarded and the cell resuspended in a complete medium. Cells were cultured with a density of 106 cells/mL in a complete medium and then incubated in 5% CO2 incubator at 37°C for 48 hours. Proliferative activity of culture supernatant was measured by a MTT (3-(4, 5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide) test assay.
Phagocytic Activity of Macrophages
On day 7, rats were sacrificed with a light anesthetic. The abdomen was opened and cleared from the peritoneal sheath with alcohol 70% (v/v) and cold RPMI-1640 (Sigma Chem, St Louis, MO, USA) was injected into the peritoneal cavity. The animals were allowed to stand for 3 minutes until macrophages attached to the peritoneal cavity and around the intestine can be detached and suspended in the RPMI medium. Peritoneal fluid was removed from the peritoneal cavity and aspirated with a syringe. Peritoneal fluid was aspirated and centrifuged at 1200 rpm, 40°C for 10 minutes. Supernatant was removed and then added 3 mL of complete RPMI medium containing fetal bovine serum.
The phagocytic activity was performed in vitro according to the previous method using a diameter of 3.0 μm latex beads (Sigma Chem). 8 The beads were resuspended in phosphate-buffered saline (Sigma Chem) in order to get concentrations of 2.5 × 107/mL. Peritoneal macrophages were cultured previously washed twice with RPMI-1640, and add 200 mL of suspension of beads in each wells and incubated in a 5% CO2 incubator, 37°C for 60 minutes. The cells then washed with phosphate-buffered saline 3 times, then dried at room temperature and fixed with methanol. Methanol was then removed and allowed to stand until dry cover slips. After drying, cover slips were stained with Giemsa (Merck, Darmstadt, Germany) 20% (v/v) for 30 minutes and washed with distilled water, removed from culture wells and dried at room temperature. The macrophage cells (100 cells) were observed using a light microscope and counted the number of phagocytic activity of macrophages to latex bead particles and the number of latex beads that uptake by macrophages.
Determination of CD8 Profiles by Flow Cytometry
Blood samples were collected from treated rats on days 0 and 7 under anesthetized condition and kept in a vacutainer containing ethylenediamine tetra-acetic acid. Sample preparation was performed by mixing 5 μL of whole blood and 10 μL was rat antibody CD3 antigen fluorescein isothiocyanate and PE anti-rat CD8a and then was vortexed gently and allowed to stand in a dark room for 15 minutes. For dilution, lysing reagent was added, then allowed to stand in a dark room for 15 minutes. After immunolabeling, cells were analyzed on a FACS Calibur Flow Cytometer and CellQuest Pro IVD software (Becton Dickenson, Mountain View, CA, USA).
Detection of Cytokines Levels
Tumor necrosis factor-α, IL-1α, and IL-10 levels were measured by enzyme-lnked immunosorbent assay using the commercially available kits listed (Table 1), as previously described. 9 One hundred milliliters of standard (diluted in the lysis buffer) or each properly diluted sample of plasma was analyzed in triplicate, and a portion of the sample was assayed for protein content. Data were expressed as pg/mg of protein.
Primary Antisera Used in This Study.

Abbreviations: ELISA, enzyme-linked immunosorbent assay; FITC, fluorescien isothiocyanate; IL, interleukin; TNF-α, tumor necrosis factor-α.
aAntisera were raised in rat.
Statistical Analysis
Data from all experiments were presented as mean values with ± standard error of mean (SEM). Statistical analysis was performed by 1-way analysis of variance with Bonferroni post hoc test (2-tailed). P < .05 was considered statistically significant.
Results
Proliferation of Lymphocytes and Phagocytic Activity of Macrofages
The ethanolic extract of T flagelliforme administration (250-1000 mg/kg body weight) restored the level of lymphocyte proliferation (Table 2) after cyclophosphamide induction, while that of 250 mg/kg body weight was the optimum dose to increase the proliferation. Cyclophosphamide also suppressed the number of macrophages and phagosytic activity of macrophages (P < .05; Table 2). The administration of ethanolic extract of T flagelliforme (250-1000) mg/kg body weight was found to significantly increase the number and activity of macrophages in phagocytic uptake of latex beads (P < .05). The optimum dose of the extract of T flagelliforme was 250 mg/kg body weight, while the subsequent dose increase even reduced the phagocytic activity of macrophages (Table 3).
Effect of Ethanolic Extract of Thyphonium flagelliforme on Proliferation of Lymphocytes in Cyclophosphamide-Treated Rats.a
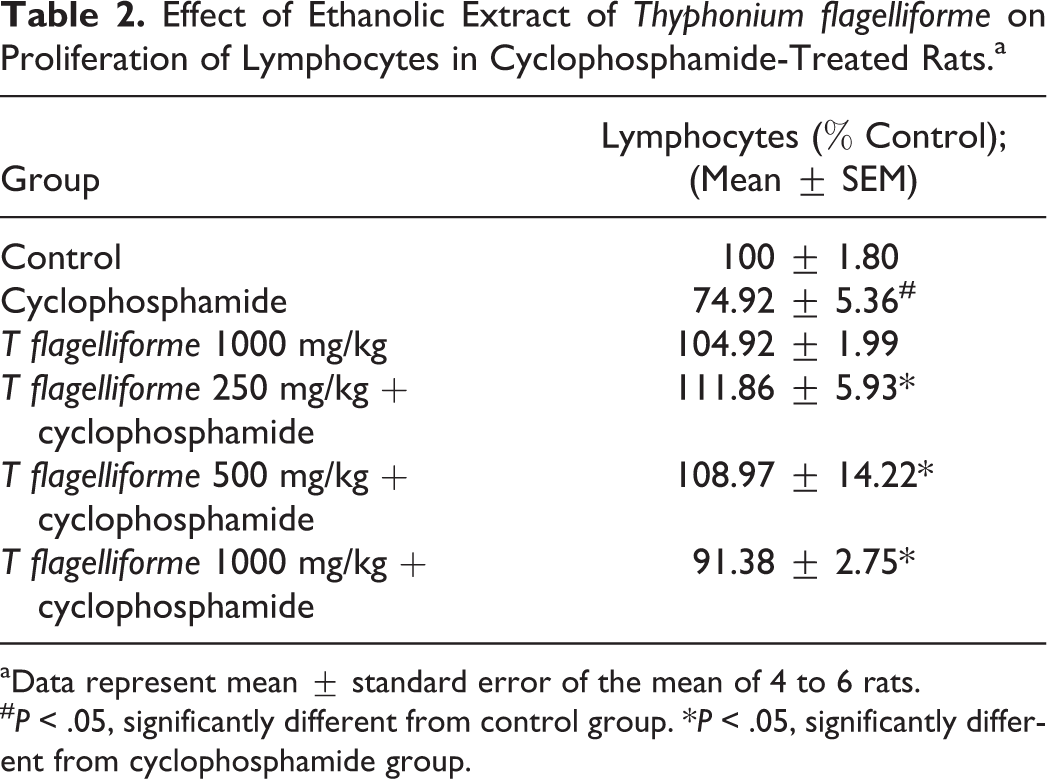
aData represent mean ± standard error of the mean of 4 to 6 rats.
# P < .05, significantly different from control group. *P < .05, significantly different from cyclophosphamide group.
Effect of Ethanolic Extract of Thyphonium flagelliforme on Phagocytic of Latex Beads by Rat Peritoneal Macrophages in Cyclophosphamide-Treated Rats.

aData represent mean ± standard error of the mean of 4 to 6 rats.
# P < .05, significantly different from control group. *P < .05, significantly different from cyclophosphamide group.
Determination of CD8+T cells by Flow Cytometry
The results demonstrated that ethanolic extract of T flagelliforme administration (250-1000 mg/kg body weight) also restored the level of CD8+ T cells in rats induced by cyclophosphamide (Table 4, Figure 1). The results showed that the administration of cyclophosphamide significantly suppressed CD8+ T cells and the ethanolic extract of T flagelliforme of 250 to 1000 mg/kg body weight significantly improved the rats CD8+ T cells (P < .05). The optimum dose of the ethanolic extract occurred at 500 mg/kg body weight (Table 4). The administration of ethanolic extract of T flagelliforme alone was increased CD8+ T cells compared with the baseline value (Table 4, Figure 1).
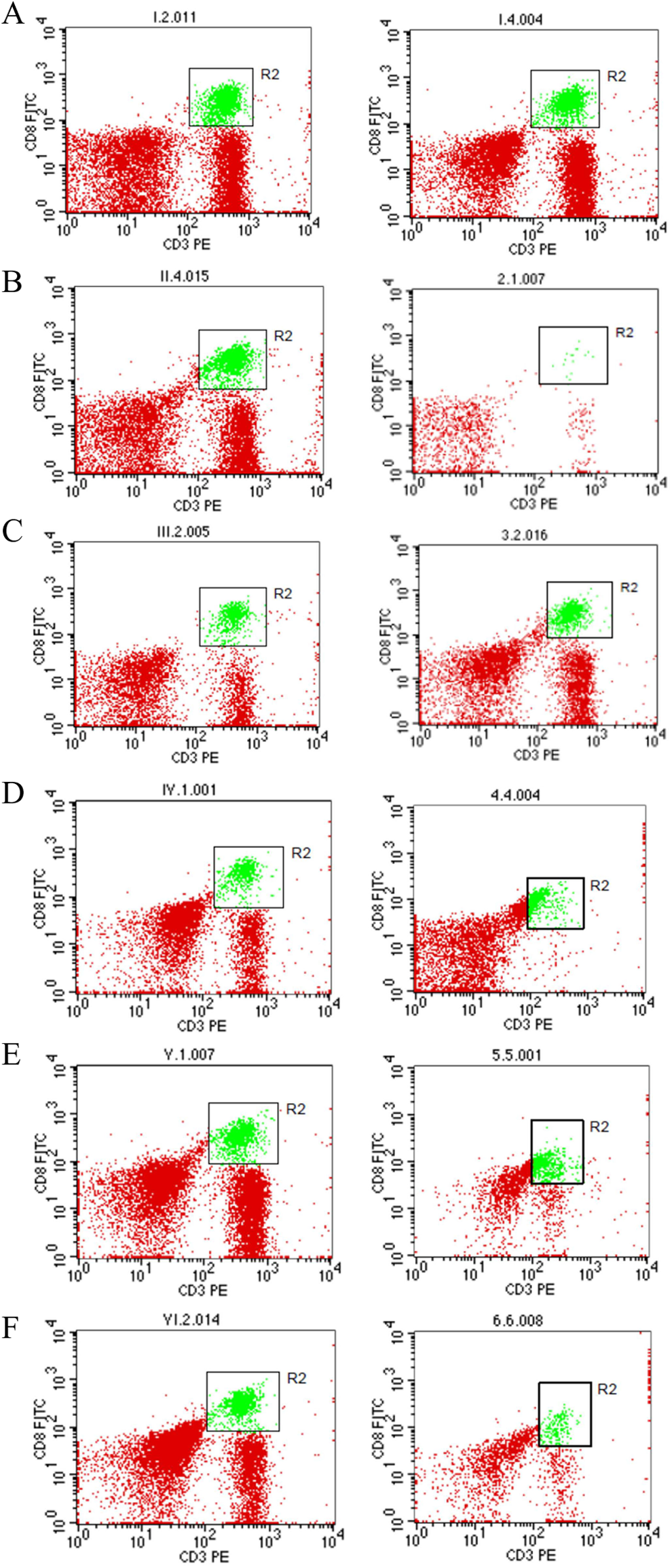
Effect of ethanolic extract of Thyphonium flagelliforme on CD8+ T cells in rats induced by cyclophosphamide. Measurement of CD8+ counts performed on day 0 (pretreatment) and day 7 (posttreatment). (A) control group; (B) cyclophosphamide-treated group; (C) ethanolic extract of T flagelliforme 1000 mg/kg body weight group; (D) ethanolic extract of T flagelliforme 250 mg/kg body weight + cyclophosphamide group; (E) ethanolic extract of T flagelliforme 500 mg/kg body weight + cyclophosphamide group; (F) ethanolic extract of T flagelliforme 1000 mg/kg body weight + cyclophosphamide group. Profile figure is one of the sample CD8+ T cells of each treatment group from 4 to 6 rats.
Effect of Ethanolic Extract of Thyphonium flagelliforme on CD8+ T Cells in Rats Induced by Cyclophosphamide.a

aData of CD8+ T cells were measured on day 0 (pretreatment) and day 7 (posttreatment), which represent mean ± standard error of the mean of 4 to 6 rats.
# P < .05, significantly different from control group. *P < .05, significantly different from cyclophosphamide group.
Detection of Cytokines
Furthermore, immunomodulatory effect of ethanolic extract of T flagelliforme on tissue cytokine levels was also measured. The results showed that the administration of cyclophosphamide also increased significantly the levels of TNF-α and IL-1α, but not the IL-10 (P < .05). The ethanolic extract of T flagelliforme dose of 250-1000 mg/kg body weight was shown to significantly reduce the suppressive effect of cyclophosphamide on cytokine levels (Table 5) with the optimum dose of 250 mg/kg body weight (P < .05).
Effect of Ethanolic Extract of Thyphonium flagelliforme on the Cytokines (TNF-α, IL-1α, and IL-10) Level in Rats Induced by Cyclophosphamide.

Abbreviations: IL, interleukin; TNF-α, tumor necrosis factor-α.
aData represent mean ± standard error of the mean of 4 to 6 rats.
# P < .05, significantly different from control group. *P < .05, significantly different from cyclophosphamide group.
Discussion
We have demonstrated in the present study that the oral administration of ethanolic extract of T flagelliforme has significant immunomodulatory effects in rats induced by cyclophosphamide. The ethanolic extract of T flagelliforme was able to restore the lymphocyte proliferation, phagocytic activity of macrophages, CD8+ T cells profile, as well as the levels of cytokines TNF-α, IL-1α and IL-10.
Cyclophosphamide is an antineoplastic drug that is widely used, in single or combination with other products. 10 Cyclophosphamide, is known to be biologically active after biotransformed by microsomal enzymes, leading to the production of a number of active metabolites capable of alkylating nucleic acid and chromosome damage through the generation of free radicals, thus promoting mutations. 11,12 Previous reports suggests that the cyclophosphamide can be used as an immunosupressive agent. Several lines of evidence demonstrated that cyclophosphamide was able to induce immunosuppression and or mutagenicity in mice and has been used as a valuable animal model to detect the effects of immunomodulatory or antimutagenic. 13,14
Lymphocytes have the ability to distinguish foreign objects from its own network, as it has on the cell surface receptor, Toll cell receptor. T lymphocytes (T cells) function in helping B cells produce antibodies, which recognize and destroy infected cells, activate macrophages in phagocytosis and the control threshold and the quality of the immune system. Macrophages are one of the effector cells to eliminate the parasite through the mechanism of phagocytosis in the immune system is not specific. The activity of macrophages can be enhanced by immunostimulant agents, either vaccine or chemical compounds, including compounds from natural products. T cell antigen receptors are always membrane bound and noncovalently associated with a set of four invariant glycoproteins collectively called CD3. Thus, CD3 has been regarded as a marker for T cells. Generally, T helper cells are also known as CD4+ T cells, cytotoxic T cells are known as CD8+ T cells. 15 Cytotoxic T cells CD8+ acts as a factor of antitumor adaptive immune system of the host and become an important prognostic factor in cancer treatment, for example, in cases of colorectal cancer. 16,17
Tumor necrosis factor-α may enhance the immune response in vivo to enhance proliferation of B and T cells and stimulate the formation of cytotoxic T cells. In addition, TNF-α increases IL-2 level, which induces the production of immunoglobulins and an increase in IL-2-stimulated natural killer cell activity and proliferation of monocytes. 18 IL-1 is another cytokine released by various cell types including macrophages, dendritic cells, lymphocytes, endothelial cells, fibroblasts, and keratocytes. Interleukin exists in 2 forms, IL-1α and IL-1β, both 17-kDa glycoproteins, the latter is secreted by cells and membrane bound forms of the former. IL-1 is required for the successful initiation of some forms of immune response. 19 Forms of other cytokines, IL-10 is an immunosuppressive glycoprotein 19 to 21 kDa in size that is secreted by Th2 cells, some B cells, and macrophages are activated. Now, it is clear that IL-10 primarily acts on activated macrophages to suppress the secretion of IL-1, IL-12, TNF-α, and reactive oxygen radicals. 18
In the present study, we found that treatment with ethanolic extract of T flagelliforme effectively reduced decreases in T cell subsets and the level of TNF-α. Therefore, we suggest that ethanolic extract of T flagelliforme can promote the proliferation of immune cells or directly inhibit cyclophosphamide-induced immunosuppressive activities. The observed increases in TNF-α levels following ethanolic extract of T flagelliforme treatment are considered to be responses secondary to the increases in T-cell numbers. However, we do not exclude the possibility of direct stimulation of the secretion of TNF-α. The increase in TNF-α, IL-1α, and IL-10 levels contents detected in the present study is considered to be evidence that ethanolic extract of T flagelliforme can promote the activity of cells producing these cytokines. However, further studies are necessary to elucidate the detail mechanism of the immunomodulatory effects of ethanolic extract of T flagelliforme.
In conclusion, immunosuppressive effect induced by cyclophosphamide in rats can be effectively reduced by the oral administration of ethanolic extract of T flagelliforme. The immunomodulatory effect of ethanolic extract of T flagelliforme on cyclophosphamide-induced rats, at least in part act through the recruitment of T cells and cytokines or enhance their activity. The results of this study indicate that ethanol extract of T flagelliforme has an immunomodulatory effect on cyclophosphamide-induced rats. This results suggest that the ethanolic extract of T flagelliforme can reduce the side effects of chemotherapy for cancer treatment.
Footnotes
Author Contributions
AN, MI, and RM were involved in plant collection, processing, and carrying out the experimental work. IPS and AEN supervised the overall study. AN and MI drafted the article. All authors read and approved the final version of the article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We are thankful to the Ministry of Education of Indonesia for providing funding for the study via the National Strategic Research Competition Grant (Grant Letter No. 389/SP2H/PL/Dit.Litabmas/IV/2011).
Ethical Approval
The experimental protocol was conducted in accordance with the Guideline for Care and Use of Animals Laboratory and approved by the Institutional Animal Ethics Committee (IAEC) of Universitas Gadjah Mada, Yogyakarta, Indonesia.