
Abstract
Introduction:
Pyogenic granuloma (PG) is a reactive inflammatory vascular lesion of the skin and mucous membranes, characterised by the presence of enlarged venules and seamed and seamless capillaries with plump endothelial cells (EC), and numerous macrophages. EC activation upregulates the synthesis of galectins and induces their translocation to the EC surface promoting angiogenesis and lymphangiogenesis, particularly galectin-1 (Gal-1), Gal-3 and Gal-8. However, the presence and distribution of Gal-1, -3 and -8, as well as their implications in the pathogenesis of PG, has not been considered.
Materials and methods:
Eight biopsies from patients diagnosed with PG were selected. The presence of PECAM-1/CD31, IL-1β, VEGF-C, VEGFR-2, VEGFR-3, integrin β1, CD44, fibronectin and Gal-1, -3 and -8 was assessed by immunofluorescence staining using confocal laser scanning microscopy.
Results and Discussion:
Immunostaining revealed that these molecules were present in the enlarged venules with plump ECs, in some macrophages and other immune cells. We propose that macrophages release VEGF-A and VEGF-C inducing VEGFR-2/VEGFR-3 expression and activation, leading macrophages to transdifferentiate into plump ECs that might integrate into pre-existing venules, contributing to the formation of enlarged venules with transluminal bridges and capillaries. EC activation, induced by certain cytokines, has been shown to stimulate galectin expression and changes in the cellular localisation through association and activation of specific EC surface glycoproteins. Therefore, it is plausible that Gal-1, -3 and -8, acting in a concerted manner, could be mediating the transdifferentiation of macrophages into plump ECs and facilitating their migration and incorporation into the new vessels.
Lay Summary
In this study, immunostaining of pyogenic granuloma (PG) tissue sections showed immunoreactivity for PECAM-1/CD31, IL-1β, VEGF-C, VEGFR-2 and VEGFR-3, and galectin-1, -3 and -8 in enlarged venules with plump endothelial cells (EC), as well as in some macrophages and other immune cells. Interestingly, enlarged and thin-walled transient vessels lined by PECAM-1/CD31 and VEGFR-2 immunopositive ECs that form from pre-existing normal venules in response to VEGF-A (called ‘mother’ vessels [MV]) and that undergo intraluminal bridging evolving into various types of capillaries (called ‘daughter’ vessels [DV]) have been observed in benign and malignant tumours, in physiological and pathological angiogenesis as well as in vascular malformations, suggesting an important role for VEGF-A and VEGFR-2 in such a process. However, it is not only the mechanisms by which the MVs evolve in different types of DVs that remains to be elucidated, but also whether the cells that form intraluminal bridges proceed from locally activated ECs or whether they are derived from bone marrow precursors or from resident macrophages.
Given that the formation of homodimers by Gal-1 and Gal-8 and pentamers by Gal-3 to generate gal–glycan lattices at the cell surface and in the extracellular space has been shown, it is possible that in PG tissue Gal-1, -3 and -8, through their binding partners, form a supramolecular structure at the surface of ECs and plump ECs, macrophages and in the extracellular space that might be mediating the transdifferentiation of macrophages into plump ECs and facilitating the migration and incorporation of these cells into the pre-existing venules, thus contributing to the formation of MVs and DVs.
Introduction
Pyogenic granuloma (PG), also referred to as lobular capillary haemangioma, is a reactive inflammatory vascular lesion of the skin and mucous membranes that can equally affect both sexes and occurs at all ages. There is controversy concerning its nature. Some authors consider PG as a benign vascular tumour, while others define it as a reactive inflammatory lesion.1–5 PG is histologically characterised by the presence of enlarged venules with plump endothelial cells (ECs) or immature ECs, seamed and seamless capillaries arranged in a lobular pattern in the dermis, and numerous macrophages, which are frequently associated with angiogenesis.1,3–6 With respect to the presence of plump ECs and seamed and seamless capillaries, previous studies have shown an increased expression of vascular endothelial growth factor (VEGF) and bFGF limited to plump ECs with no lumen formation, many of them expressing PECAM-1/CD31.7–9 The same studies point to macrophages as the major source of synthesis and secretion of both angiogenic factors7–9 and that vascular growth observed would be associated with the activation of the vascular endothelial growth factor receptor-3 (VEGFR-3/Flt4).7–9 Equally well established is the important role during the early phase of inflammation and wound healing of activated macrophages as an important source of pro-inflammatory cytokines and growth factors such as IL-1β, IL-1α, IL-6. IL-12, TNFα, VEGF-A, VEGF-C and bFGF,9,10 and that overexpression of VEGF-C, the main ligand for VEGFR-3, would be involved in the recruitment, accumulation and transdifferentiation of macrophages into endothelial-like cells during inflammation.11,12 Other studies have suggested an important contribution of macrophage as endothelial progenitor during the lymphangiogenesis process.13–18 Although the plump ECs have been identified in PG, only its presence and localisation have been examined, while its progenitor and possible contribution to the formation of enlarged venules and capillaries have received little attention.
It has further been reported that the activation of ECs by inflammatory cytokines and growth factors secreted by different immune cell types, including macrophages, contributes in the pathogenesis of many skin diseases frequently associated with the enlargement and formation of new capillaries from the pre-existing vasculature (angiogenesis and lymphangiogenesis) and that the major regulator of the angiogenesis cascade and wound repair released during inflammation is the VEGF-A, whose functions are mediated by two receptors tyrosine kinases, VEGFR-1 (Flt-1) and VEGFR-2 (Flk-1),19,20 while in inflammatory lymphangiogenesis VEGF-C is the main ligand that activates VEGFR-3 (Flt-4).8,21–26
Equally, previous reports have suggested that EC activation upregulates the synthesis of galectins and induces their translocation to the EC surface promoting angiogenesis and lymphangiogenesis via association and activation of specific EC surface glycoproteins, particularly galectin-1 (Gal-1), Gal-3 and Gal-8.27–34
Gal-1 is a prototypic member of the galectin family with a conserved carbohydrate recognition domain (CRD) that has affinity for multiple N-acetyllactosamine (LacNAc) disaccharides (Gal-GlNAc) present in N- and O-linked glycans.35–38 By association of their monomers, it can form homodimers through its C-terminal domain on the cell surface and in the extracellular space, which results in the formation of molecular complexes.35–38 This lectin participates in signalling pathways, regulating a variety of important biological responses including cell growth, proliferation, cell–cell and cell–matrix adhesion, cell differentiation, motility and cell death, and plays a critical role in the initiation and resolution of the inflammatory response, angiogenesis, fibrosis, and tumour development.35–38 Gal-3 is a chimera type galectin that is composed of a C-terminal domain that binds to specific N- and O-glycan ligands, and an N-terminal domain which facilitates its multidimerisation and generation of galectin–glycan (gal–glycan) lattices on the cell membrane and in the extracellular space controlling several cellular processes through specific signalling pathways; those processes include cell–cell adhesion, cell–matrix adhesion, proliferation, growth, differentiation, migration, inflammation, immune response, apoptosis, angiogenesis and tumour development.39–45 It is predominantly localised in the cytoplasm but can also be detected in the nucleus, intracellular and extracellular sides of the cell membrane, and found in the extracellular space.28–32,43,45 Gal-8 is a tandem-repeat-type galectin that has two CRDs joined by a linker peptide. It appears to be one of the most prominent galectins detected in ECs.28,30,33 It plays an important role in the control of EC migration, capillary tube formation and in vivo angiogenesis33,46–50 and tumour growth. 51 In addition, Gal-8 modulates lymphangiogenesis induced by VEGF-C in vivo and in vitro.28,30,33 Nevertheless, to the best of our knowledge, the presence and distribution of Gal-1, -3 and -8, as well as their implications in the pathogenesis of PG, has not been considered.
In view of the above considerations, the aim of this study was to examine not only the presence and localisation of the plump ECs in PG, but also establish its origin and contribution to the formation of enlarged venules and new capillaries.
Given that in human skin some galectins play a crucial role in diverse cutaneous disorders,38,43,52–54 we sought to determine whether Gal-1, -3 and -8 were present in PG. We also examined whether these galectins could generate gal–glycan lattices able to regulate the behaviour of the plump ECs through the interactions with platelet endothelial cell adhesion molecule-1 (PECAM-1/CD31), VEGFR-2, VEGFR-3, CD44, integrin β1 and fibronectin (FN). All of them were considered as targets for these galectins as they contain N- and/or O-linked glycans in the extracellular domains.
Materials and methods
Skin biopsies
Eight biopsies from patients clinically and histopathologically (haematoxylin and eosin [H&E] staining) diagnosed with PG were retrieved from the files of the Section of Dermatopathology at the ASIB. Data regarding gender, age and site of lesion appear listed in Table 1. For the identification of collagen bundles, some histological sections were stained following Masson’s trichrome protocol. This study was approved by the Ethics Committee of the SAIB and performed according to the Declaration of Helsinki.
Patients diagnosed with PG included in the study.

F, female; M, male; PG, pyogenic granuloma.
Indirect immunofluorescence staining of tissue sections
For immunohistochemistry purposes, paraffin-embedded skin tissue blocks were cut into sections each with a thickness of 4 µm. Paraffin sections were then set onto silanised slides (DakoCytomation, Denmark), dewaxed in xylene, dehydrated through graded alcohol series (from 100 to 70%), then rehydrated in distilled water and equilibrated in phosphate-buffered saline (PBS) for 10 min. Before incubation with the primary antibodies, sections were blocked for 1 h in a humid chamber with PBS containing 2% BSA and 0.1% Tween 20, in order to prevent non-specific staining. Sections were then incubated with the primary antibodies diluted in PBS, 2% BSA, 0.1% Tween 20 (Table 2) overnight at 4 °C followed by incubation with anti-rabbit IgG Cruz Fluor 594 (Santa Cruz Biotechnology, Inc., Dallas, TX, USA) or anti-mouse IgG Cruz Fluor 488 (Santa Cruz Biotechnology, Inc.) diluted with PBS containing 2% BSA and 0.1% Tween20 for 30 min in a humid chamber in the dark. Cell nuclei were counterstained with DAPI (4’,6-Diamidino-2-phenylindole) (Molecular Thermo-Fisher, Waltham, MA, USA) for 15 min in the dark. Finally, the sections were washed in PBS and coverslipped with mounting medium (IMMU-MOUNT, Shandon, Pittsburgh, PA, USA). All images were captured using a 1X81 Olympus inverted microscope with the Fluo View Confocal Laser Scanning configuration (CLSM) (Olympus America) equipped with software program FV10.ASW version 02.01.01.04 (Olympus Corporation). Image J software (NHI, Washington, DC, USA) was used for the processing of contrast and brightness.
Antibodies.
Results
Histopathological analysis of the PG tissues by Masson’s trichrome staining showed the presence of enlarged, thin-walled venules with some plump ECs in the dermis, some of these cells forming intraluminal bridges that divide the lumen into small channels and capillaries (Figure 1). Staining also revealed the presence of numerous capillaries lined by some plump ECs localised close to the enlarged venules, as well as the presence of some vascular structures known as glomeruloid bodies, numerous macrophages and other immune cells (Figure 1). They were all embedded in a collagen matrix (Figure 1).

Masson trichrome staining from two biopsies of PG tissues. Biopsy 1, lesion located on upper lip. Biopsy 2, lesion located on thorax. Note the presence of enlarged venules with some plump ECs (V, arrowheads), small channels (asterisks), capillaries (C) and macrophages (arrows). They were all embedded in a collagen matrix. Yellow dotted line shows glomeruloid body. Enlargements show macrophages. Scale bars: 20 µm; 10 µm. EC, endothelial cell; PG, pyogenic granuloma.
In vivo PECAM-1/CD31 immunostaining
Platelet endothelial cell adhesion molecule-1 (PECAM-1/CD31) is a transmembrane glycoprotein with proinflammatory and proangiogenic activities that is highly expressed on the surface of ECs and lower levels on platelets and immune cells including monocytes, macrophages, and T and B cells.55–59 It contains seven N-linked glycans and interacts with VEGFR-2, VEGFR-3 and integrins. 60 For these reasons, we examined the presence and distribution of this glycoprotein on the ECs of vessels of PG tissues. Immunolabelling analysed by CLSM showed that PECAM-1 was present in the enlarged venules with plump ECs, small channels and capillaries (Figure 2a). Interestingly, a strong PECAM-1 immunoreactivity was appreciable in the plump ECs that were close to and incorporated into the enlarged venules, forming intraluminal bridges, as well as in some plump ECs of small channels and capillaries that were mostly situated close to enlarged venules. Likewise, PECAM-1 immunoreactivity was present in some macrophages and other immune cells (Figure 2a).

Immunostaining of PECAM-1/CD31, IL-1β, VEGF-C, VEGFR-2, VEGFR-3, Gal-1 and Gal-3 in the dermis reticular of one biopsy of PG tissue (lower lip). Serial sections were stained for PECAM-1/CD31 (a), IL-1β (b), VEGF-C (c), VEGFR-2 (d), VEGFR-3 (e), Gal-1 (f) and Gal-3 (g). Enlargements show the corresponding immunoreactivities in the venules (V), small channels (asterisks), capillaries (C), plump ECs (arrowheads), macrophages (arrows) and immune cells (stars). Blue, DAPI. Scale bars: 30 µm; 15 µm. EC, endothelial cell; PG, pyogenic granuloma.
In vivo IL-1β and VEGF-C immunostaining
Because IL-1β promotes the EC activation during inflammation and angiogenesis by increasing expression of adhesion molecules on the surface of ECs61–63 and it regulates the expression of VEGF-C, 64 we examined the presence and distribution of IL-1β and VEGF-C in the dermis of the PG tissues selected. Examination of the dermis revealed IL-1β and VEGF-C immunoreactivities localised in the enlarged venules with plump ECs and small channels and capillaries, as well as in some macrophages and other immune cells (Figure 2b and c).
In vivo VEGFR-2 and VEGFR-3 immunostaining
Considering that VEGF-C is a secreted glycoprotein that has an affinity for VEGFR-2 and VEGFR-323,64 and that both tyrosine kinase receptors contain N-linked glycans,65,66 we also examined the presence and distribution of VEGFR-2 and VEGFR-3 in the dermis of PG tissues. Immunostaining showed that both receptors were localised in the enlarged venules with plump ECs, small channels and capillaries, as well as in some macrophages and other immune cells (Figure 2d and e). Like PECAM-1, strong VEGFR-2 and VEGFR-3 immunoreactivity was detected in the plump ECs that were close to and incorporated into the enlarged venules, as well as in the plump ECs that were forming transluminal bridges, small channels and capillaries, and some macrophages and other immune cells (Figure 2d and e).
In vivo Gal-1, Gal-3 and Gal-8 immunostaining
To determine whether these galectins were present in the dermis of PG, tissue sections were also examined by immunofluorescence staining. Immunolocalisation revealed that Gal-1, -3 and -8 were present in the enlarged venules with plump ECs and small channels and capillaries, as well as in some macrophages and other immune cells (Figures 2f, 2g, 3a and 4a–c). Notably, an intensive cytoplasmic and cell membrane immunoreactivity for Gal-1, -3 and -8 was detected in the plump ECs that were close to and incorporated into the enlarged venules, as well as in the plump ECs, forming intraluminal bridges, small channels and capillaries, and some macrophages and immune cells (Figures 2f. 2g, 3a and 4a–c). In addition, a moderate immunoreactivity for these galectins was detected in fibrils and thin bundles of collagen and some elongated fibroblasts (Figure 4a–c). No immunoreactivity was detected when the primary antibodies were omitted or replaced with PBS (not shown).
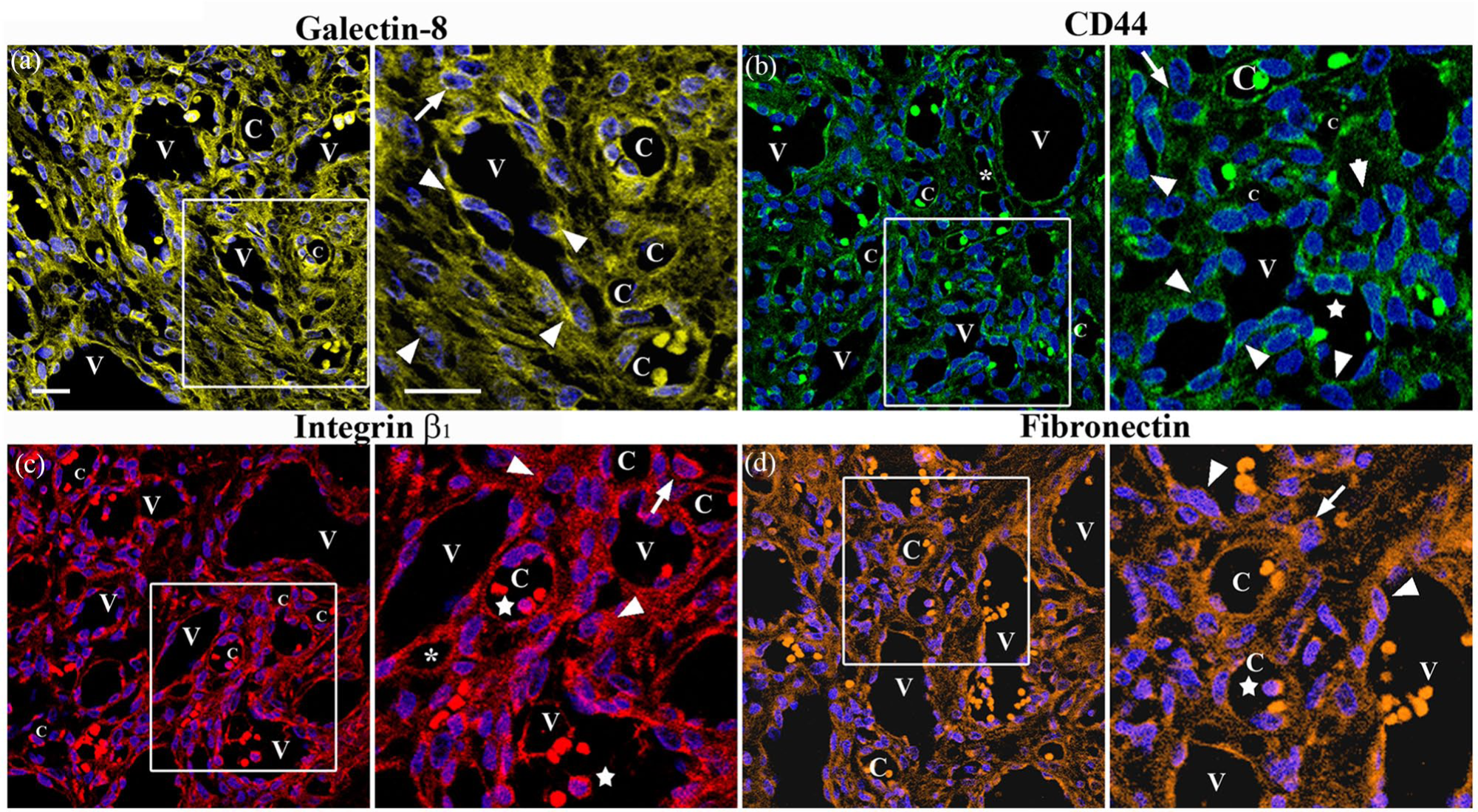
Immunolocalisation of Gal-8, CD44, integrin β1 and FN in the reticular dermis of the same biopsy. Serial sections were stained for Gal-8 (a), CD44 (b), integrin β1 (c) and FN (d). Enlargements show the corresponding immunoreactivities in the venules (V), small channels (asterisks), capillaries (C), plump ECs (arrowheads), macrophages (arrows) and immune cells (stars). Blue, DAPI. Scale bars: 30 µm; 15 µm. EC, endothelial cell; FN, fibronectin; PG, pyogenic granuloma.
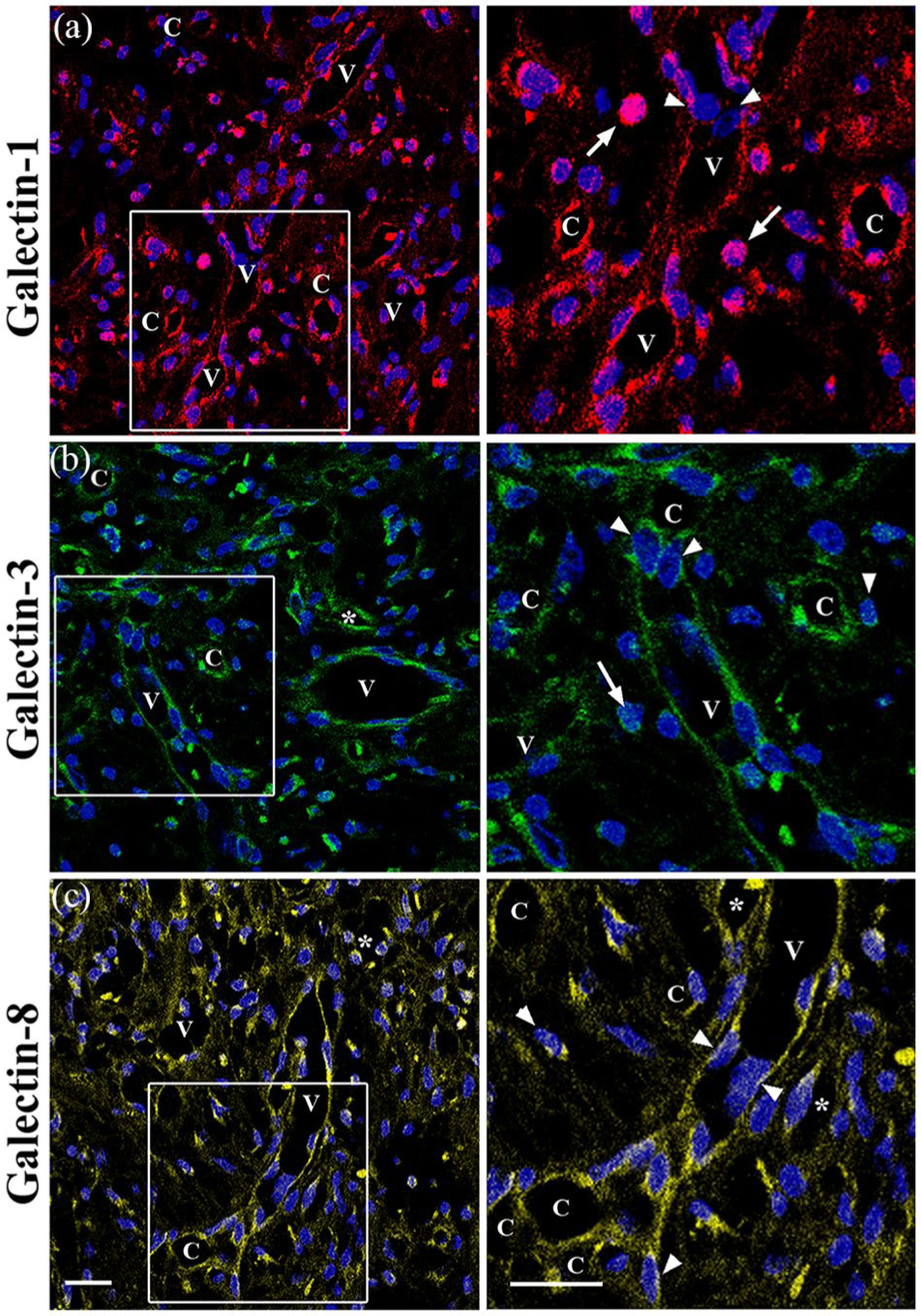
A series of representative CLSM fluorescence images of Gal-1, -3 and -8 in one biopsy of the PG (finger). Sections were stained for Gal-1 (a), Gal-3 (b) and Gal-8 (c). Enlargements show the corresponding immunoreactivities in the venules (V), small channels (asterisks), capillaries (C), plump ECs (arrowheads), macrophages (arrows) and immune cells (stars). Blue, DAPI. Scale bars: 20 µm; 10 µm. EC, endothelial cell; PG, pyogenic granuloma.
In vivo CD44 immunostaining
Since CD44 is a glycosylated cell-surface receptor that has an N-terminal that contains N- and O-linked glycans67–69 and can interact with Gal-8 contributing to the resolution of inflammation, 70 we also examined the presence and distribution of CD44 in the dermis of PG tissues selected. CD44 was observed in the enlarged venules with plump ECs, small channels and capillaries as well as in some macrophages and other immune cells and some fibroblasts and fibrils and collagen bands (Figure 3b).
In vivo integrin β1 immunostaining
Because integrin β1 contains N-linked glycans71,73 and is considered as binding partner for Gal-1,35,73 Gal-3,47,74–76 and Gal-8,47,49,77,78 we investigated the presence and distribution of integrin β1 in the dermis of PG tissues. Immunostaining showed that this subunit was localised in the enlarged venules with plump ECs, small channels and capillaries (Figures 3c and 4a–c), as well as in some macrophages, immune cells, fibroblasts and collagen bands (Figure 3c).
In vivo FN immunostaining
We next examined the presence and distribution of FN, a glycoprotein that contains N-linked glycans 79 and associates with Gal-1, 35 Gal-3 76 and Gal-8.47,80 Examination of the dermis revealed that FN was localised in the enlarged venules with plump ECs, small channels and capillaries (Figure 3d). Moderate immunoreactivity was also observed in some fibroblasts and thin bundles of collagen and in the extracellular space (Figure 3d).
Discussion
In the present study, immunostaining of PG tissue sections showed immunoreactivity for PECAM-1/CD31, IL-1β, VEGF-C, VEGFR-2 and VEGFR-3 in enlarged venules with plump ECs, including those cells that were forming transluminal bridges and dividing the lumen into small channels and capillaries, as well as in some macrophages and other immune cells. It is important to stress that this immunoreactivity increased in the plump ECs. Remarkably, studies conducted by Caramelo’s group in PG had revealed the presence of EC precursors or plump ECs without lumen formation that displayed increased expression of VEGF and immunoreactivity for PECAM-1/CD31, suggesting that early VEGF expression might act as angiogenic mediator and that the presence of local inflammatory cytokines such as IL-1β and TNFα in PG would explain the induction of VEGF in these cells. 6 Other studies in PG report the presence of aggregates of capillaries and venules lined by ECs and plump ECs positive to CD31, CD34, CD105; ICAM-1 and VCAM-1.1–5,81 Of particular interest, studies in PG have also shown VEGF and bFGF upregulation, considering the macrophages as the major source of synthesis and secretion of both angiogenic factors.2,4,7 It is noteworthy that the angiogenesis observed in PG has been associated to the activation of Flt4/VEGFR3 and the nitric oxide signalling pathway. 8 Likewise, overexpression of VEGF-C has been shown to induce enlargement of veins and venules positive for PECAM-1/VEGFR-2 in skin and mucous membranes. 9 Interestingly, enlarged and thin-walled transient vessels lined by PECAM-1/CD31 and VEGFR-2 immunopositive ECs that form from pre-existing normal venules in response to VEGF-A (called ‘mother’ vessels [MV]) and that undergo intraluminal bridging evolving into various types of capillaries (called ‘daughter’ vessels [DV]), have been observed in benign and malignant tumours, in physiological and pathological angiogenesis as well as in vascular malformations, suggesting an important role for VEGF-A and VEGFR-2 in such a process.81–85 However, it is not only the mechanisms by which the MVs evolve in different types of DVs that remains to be elucidated, but also whether the cells that form intraluminal bridges proceed from locally activated ECs or whether they are derived from bone marrow precursors or from resident macrophages.83,84 Of relevance, studies in vitro and in vivo have shown that overexpression of VEGF induces macrophages to transdifferentiate into EC progenitors or endothelial-like cells characterised by expression of VE-cadherin, CD31, CD105 and eNOS, which are able to incorporate in the newly formed vessels.11,12 In this context, other studies report that lymphatic EC progenitors derived from macrophages integrate into lymphatic endothelium. The same studies propose that the VEGF-C/VEGFR-3 signalling pathway could play a critical role in the early phase of transdifferentiation of macrophages and that the progenitors of the lymphatic ECs may originate from macrophages derived from bone marrow cells or resident macrophages that have been activated in situ.13–18 In line with these reports, contribution of lymphatic EC progenitors to lymphangiogenesis in human renal transplants has been shown, suggesting that these EC progenitors derive from macrophages expressing VEGFR-3 and incorporate into the newly growing lymphatic vessel.18,86
Based on the above evidence and our observations, it is tempting to hypothesise that in response to inflammatory stimuli, for example upregulation of IL-1β, macrophage release VEGF-A and VEGF-C inducing VEGFR-2/VEGFR-3 expression, and that VEGF-C upregulation would provoke an increase in the activation of VEGFR-2 and VEGFR-3, generating an autocrine loop that would lead macrophages to transdifferentiate into plump ECs characterised by the expression of PECAM-1/CD31, VEGFR-2 and VEGFR-3. These plump ECs which might integrate into pre-existing venules contributing to the formation of enlarged venules with transluminal bridges (MVs) and capillaries (DVs) (Figure 5).

Macrophage transdifferentiate into plump endothelial cells that integrate into pre-existing venules contributing to the formation of enlarged venules with transluminal bridges (mother vessels) and capillaries (daughter vessels).
Immunostaining also revealed that Gal-1, -3 and -8 were present in the ECs and plump ECs of the MVs and DVs, in some macrophages and other type of immune cells. Consistent with this, several in vitro and in vivo studies have shown that ECs synthesise Gal-1, -3 and -8 and that EC activation, induced by certain cytokines that are released from inflammatory and ECs at the sites of inflammation, stimulates galectin expression and induces changes in the cellular localisation through association and activation of specific EC surface glycoproteins.28,30,33,34 Similarly, studies on galectins and angiogenesis have suggested that Gal-1 can be synthesised or taken by activated ECs to promote angiogenesis in an autocrine or paracrine manner34–38 and that Gal-3 modulates angiogenesis mediated by VEGF and bFGF.39–45 Thus, the presence of Gal-1, -3 and -8, possibly stimulated by EC activation that has been induced by IL-1β,VEGF-A and VEGF-C, could be contributing to the generation of MVs and DVs with plump ECs in PG tissues.
Regarding the presence of Gal-1, -3 and -8 in some macrophages, it is well known that Gal-1 and Gal-3 are abundantly expressed and secreted by human monocytes and macrophages regulating its phenotype and functioning during acute and chronic inflammation16,29,33,39,45 and that Gal-3 modulates the production of some cytokines such as IL-1β and IL-5 in macrophages and increases the migration of monocytes and macrophages in the presence of FN, suggesting that Gal-3 can act as chemoattractant for monocytes, macrophages and ECs.45,87 Therefore, it is plausible that Gal-1, -3 and -8, acting in a concerted manner, could be mediating the transdifferentiation of macrophages into plump ECs and, in turn, facilitating the migration and incorporation of these cells into the new vessels, i.e. into MVs and DVs. Interestingly, previous studies have indicated that Gal-8, together with Gal-1 and Gal-3, modulates the invasive and migratory capacities of malignant cells.88,89
With respect to the immunolocalisation of Gal-1, -3 and -8 in the cell membrane of the ECs and plump ECs as well as in some macrophages, several in vitro and in vivo studies have shown that EC activation, in addition to upregulating the synthesis of Gal-1, -3 and -8, also induces changes in its cellular localisation and distribution through its interaction with the N- and/or O-glycans residues of glycosylated molecules that are present on the surface of ECs or in the extracellular space, modulating the cell–cell and cell–matrix interactions through specific signalling pathways. In keeping with these observations, Gal-1 and Gal-3 interact with the N-linked glycans of VEGFR-2, while Gal-8 interacts with the N-glycans of VEGFR-2 and VGFR-3, preventing the internalisation of both tyrosine kinase receptors and modulating their signalling pathways.33,78,87–89 Likewise, Gal-1, -3 and -8 have been reported to be able to interact with the N- and/or O-linked glycans of the cell surface glycoproteins CD44 and integrin β1 and with the extracellular matrix components collagen and FN, regulating cell attachment, spreading and migration.48,70 Importantly, many of the aforementioned molecules are recognised as binding partners of Gal-1, -3 and -8.
Given that Gal-1, -3 and -8, VEGFR-2, VEGFR-3, CD44, integrin β1 and PECAM-1/CD31 were immunolocalised in the reticular dermis, and that the formation of homodimers by Gal-1 and Gal-8 and pentamers by Gal-3 to generate gal–glycan lattices at the cell surface and in the extracellular space has been shown, it is possible that in PG tissue, Gal-1, -3 and -8, through their binding partners, form a supramolecular structure at the surface of ECs and plump ECs, macrophages and in the extracellular space that might be mediating the transdifferentiation of macrophages into plump ECs and facilitating the migration and incorporation of these cells into the pre-existing venules, thus contributing to the formation of MVs and DVs.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.