
Abstract
To investigate the effects of total flavonoids from Oxytropis falcata Bunge on the inflammatory signaling pathway suppressor of cytokine signaling (SOCS)/Janus kinase (JAK)/signal transducer and activator of transcription (STAT) in diabetic nephropathy KK-Ay mice. KK-Ay mice were used to establish a diabetic nephropathy model. The general condition of the mice treated with different concentrations of total flavonoids from O. falcata was monitored, respectively. Body weight, blood glucose, 24-h urinary albumin (UAlb), serum creatinine (Cre), blood urea nitrogen (BUN), and uric acid (UA) levels were measured at different time points. Hematoxylin and eosin staining quantitative reverse transcription-polymerase chain reaction and western blotting were used to detect changes in renal tissues and glomerular mesangial cells. Four weeks after model establishment, body weight, blood glucose, and 24 h UAlb significantly increased in KK-Ay mice compared with that in control C57BL/6j mice (P < 0.05). Compared with non-treated model mice, mice treated with total flavonoids from O. falcata for 4 weeks had significantly decreased serum Cre, BUN, and UA; monocyte chemoattractant protein-1(MCP-1), nuclear factor(NF)-κB, interleukin(IL)-6, and transforming growth factor(TGF)-β1, JAK 1, STAT 3 and STAT 4 mRNA levels; and p-JAK2 and p-STAT1 protein levels and significantly increased SOCS-1 and SOCS-3 protein levels in the kidneys. The treatment effects were dose-dependent and same to in vitro. Our results reflected that total flavonoids from O. falcata relieved renal tissue inflammation in diabetic mice by reducing blood glucose levels and inhibiting JAK/STAT signaling, thereby protecting against the development of diabetic nephropathy.
Introduction
Diabetic nephropathy (DN) is one of the most severe chronic microvascular complications of diabetes. It is a major cause of death in diabetes patients and the major cause of end-stage renal disease. The mechanisms underlying DN development are complex. Reported changes involved in DN development include epigenetic changes,1,2 glucose metabolism disorders,3,4 changes in renal hemodynamics,5,6 insulin resistance,7,8 and abnormalities in inflammatory responses and cytokines.9,10 In recent years, the phenomenon of inflammation has received increasing interest in the field, with suggestions that DN is an immune-mediated inflammatory disease. Studies have revealed that the inflammatory cytokines angiotensin (Ang) II, monocyte chemoattractant protein (MCP)-1, intracellular adhesion molecule (ICAM)-1, nuclear factor-(NF)-κB, vascular cell adhesion molecule (VCAM)-1, interleukin (IL)-6, IL-8, and transforming growth factor (TGF)-β1 are closely related to DN.11–13 The majority of these inflammatory cytokines are primarily secreted by immune cells such as monocytes/macrophages and lymphocytes. 14 These cytokines induce fibroblast activation and the secretion of extracellular matrix (ECM) components, such as collagen, fibronectin, and laminin, which are important factors implicated in DN development and progression.
Furthermore, these inflammatory factors interact with multiple signaling pathways in the kidneys to form a complex network that promotes the development of DN. Among the various signaling pathways, the Janus kinase (JAK)/signal transducer and activator of transcription (STAT) pathway plays a dual role in signal transduction and activation of gene transcription. It exerts biological effects by mediating intracellular signal transduction processes of multiple cytokines and activating the corresponding target genes. JAK/STAT is the key signaling pathway involved in the development of DN. In vivo studies have confirmed that JAK/STAT signaling activation results in AngII-mediated mesangial cell proliferation and stimulation of TGF-β production, leading to a further increase in ECM secretion, and aggravates the pathology of DN.15–17 Therefore, inhibiting the JAK/STAT pathway can suppress intracellular signal transduction processes of inflammatory cytokines, thereby attenuating DN development. The suppressors of cytokine signaling (SOCS) family of proteins are negative regulators of the JAK/STAT pathway. They are involved in the regulation of the wound-healing process in various diseases that may occur in various diseases such as rheumatoid arthritis and other immune diseases, cerebral hypoxia, heart failure, and tumors.18,19 Recently, the SOCS family was found to relieve the damaging effect of DN by suppressing JAK/STAT signaling. 19
Numerous studies have shown that plant flavonoids demonstrate significant efficacy in improving diabetes-related complications.20,21 Their effects include the lowering of blood sugar and blood lipids, improvement of renal function, and inhibition of glomerular hypertrophy and mesangial matrix expansion. Some of these effects have been observed clinically.22–24 Recent studies have shown that total flavonoids from Oxytropis falcata Bunge improve insulin resistance, inhibit human renal tubular epithelial cell proliferation induced by TGF-β1 in vitro, reduce ECM secretion, and exert a certain regulatory effect on fibrogenic cytokines.25–27 Based on these findings, we hypothesized that total flavonoids from O. falcata would exert a beneficial effect on DN. Therefore, using a spontaneous KK-Ay mouse model of type 2 diabetes and focusing on SOCS/JAK/STAT as the target pathway, the present study investigated the functions and mechanisms of total flavonoids from O. falcata in the prevention and treatment of DN by evaluating their effects on the expression of the inflammatory factors MCP-1, NF-κB, IL-6, and TGF-β1. The results of the present study could provide new perspectives and methodologies for preventing and treating DN.
Materials and methods
Experimental preparation
O. falcata originates from Qinghai, China. Total flavonoids were extracted at the Gansu Province Academy of Chinese Medicine, at 80% purity.
Experimental animals
Fifty 8-week-old male KK-Ay mice and ten 8-week-old male C57BL/6j mice were purchased from Beijing HFK Bioscience (Beijing, China), and all of these are specific-pathogen-free (SPF) animals.
Reagents
Hematoxylin and eosin (H&E) stain kit (C0105; Beyotime), horseradish peroxidase (HRP)-labeled goat anti-rabbit secondary antibody (A0208; Beyotime), radioimmunoprecipitation assay (RIPA) lysis buffer (P0013C; Beyotime), TRIzol (9109; Takara), PrimeScript RT Reagent Kit with gDNA Eraser (RR047A; Takara), real-time PCR (qPCR) kit (RR820A; Takara), rabbit anti-SOCS-1 polyclonal antibody (GTX100443; Genetex), rabbit anti-SOCS-3 polyclonal antibody (GTX100034; Genetex), rabbit anti-phosphorylated (p)-JAK2 polyclonal antibody (GTX41286; Genetex), rabbit anti-p-STAT1 polyclonal antibody (GTX37694; Genetex), anti-JAK2 (ab108596; Abcam), anti-STAT1 (ab31369; Abcam), SOCS-2 (Abcam; ab3692), total-STAT3 (Abcam; ab119352), p-STAT3 (Abcam; ab76315), TGF-β1(Abcam; ab92486), fibronectin(Abcam; ab2413); laminin(Abcam; ab11575), collagen type IV(Abcam; ab6586).
Experimental methods
Model establishment
Eight-week-old KK-Ay mice were used as the model control group, and C57BL/6J mice were used as the non-treat control group. The mice in the model and control groups were fed a high-fat and a normal diet, respectively. After 4 weeks of the diet, blood glucose was measured; the DN model was considered to be successfully established if the blood glucose stability was ⩾16.7 mmol/L compared with the control group.
Grouping and dosing
Total flavonoids from O. falcata were intragastrically administered at 100 mg/kg (low dose), 200 mg/kg (medium dose), and 400 mg/kg (high dose) for 4 weeks as the treatment groups. Non-treat control and model control groups were given the same volume of saline. A positive control group was treated with 10 mg/kg pioglitazone. All of mice were treated once a day for four consecutive weeks.
Sample collection
At the end of treatment week 4, the mice were fasted but allowed free access to water. After, 24 h urine samples were collected and stored at −20°C for further testing. Blood was collected from the tail vein; the serum was separated and stored at −20°C. After blood collection, the mice were euthanized. The kidneys were separated and dissected longitudinally into several sections, which were stored in 4% paraformaldehyde or liquid nitrogen based on the detection method used in subsequent analyses.
Assessment of general conditions and body weight
The general condition of the mice, amount of water and food intake, body weight, and random blood glucose levels were monitored. The body weights were recorded before and 0, 2, and 4 weeks after model establishment.
Detection of blood glucose and urinary albumin (UAlb)
Twenty-four-hour urine was collected using metal metabolic cages. The urine volume was recorded. Blood glucose was measured in blood sampled from the tail vein. UAlb and blood glucose levels were measured before and at 0, 2, and 4 weeks after the model was established.
Serum biochemical parameters
Changes in serum creatinine (Cre), blood urea nitrogen (BUN), and uric acid (UA) levels were measured after 4 weeks of treatment using an automatic biochemical analyzer.
Glomerular tissue morphology and evaluation
H&E staining was used to observe pathological changes in the renal tissues. Briefly, the renal tissue samples were fixed in 4% paraformaldehyde for 1 week, dehydrated using an alcohol gradient, embedded in wax, and cut into 4 μm slices after drying. The tissue slices were mounted and hydrated by reversing the aforementioned dehydration steps. The sections were then stained with hematoxylin at room temperature for 5 min, followed by washing and color separation with a hydrochloric acid–alcohol solution. The tissue sections were washed, stained with eosin for 30 s, and dehydrated according to the aforementioned procedure until transparent. Finally, the sections were sealed for observation. The histological was evaluated according to Lu et al. 28 The degree of damage in each glomerulus was assessed using a semi-quantitative scoring method as follows: grade 0, normal glomeruli (no damage); grade 1, mesangial expansion area, up to 25% (minimal damage); grade 2, 26%–50% expansion (moderate damage); grade 3, 51%–75% expansion (moderate-to-severe damage); and grade 4, 76%–100% expansion (severe damage). The glomerular matrix expansion index (GMI) was then calculated by the following formula: GMI = (1 × n1 + 2 × n2 + 3 × n3 + 4 × n4)/(n0 + n1 + n2 + n3 + n4).
Reverse transcription-qPCR
RNA was extracted from kidney tissues stored in liquid nitrogen using TRIzol the manufacturer’s instructions. After removal of genomic DNA and RNA quantification and quality check, the RNA was reverse transcribed into cDNA. The cDNA was used as a template for qPCRs. The thermal cycles were as follows: pre-denaturation at 95°C for 10 min, 40 cycles of 95°C for 15 s, 60°C for 15 s, and 72°C for 30 s. Relative target gene expression was calculated as 2–ΔΔCt. Primer sequences are shown in Table 1.
Primer sequences.

MCP-1: monocyte chemoattractant protein-1; NF-κB: nuclear factor; IL-6: interleukin-6; TGF-β1: transforming growth factor; JAK: Janus kinase; STAT: signal transducer and activator of transcription; SOCS: suppressor of cytokine signaling.
Western blotting
Fresh renal tissue was collected, washed with phosphate-buffered saline (PBS), and cold RIPA buffer was added to the tissue on ice. After lysis, the sample was transferred to a new centrifuge tube and the lysis reaction was continued for 30 min. Then, the tube was centrifuged at 13,000 r/min, the supernatant was collected, and the protein concentration was determined using bicinchoninic acid kit. Proteins were resolved using 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The samples were subjected to the following procedures: electrophoresis, membrane transfer, blocking, overnight incubation with primary antibody, washing with PBS, secondary antibody incubation at room temperature for 1.5 h, washing with PBS, visualization using enhanced chemiluminescence, X-ray film development, and image capturing.
Cell culture experiment in vitro
Glomerular mesangial cells were cultured in RPMI-1640 medium, penicillin and streptomycin were added to make their final concentration of 100 U/mL. The cells were purchased from China Center for Type Culture Collection (CCTCC) and stored in our laboratory. According to the experimental requirements, prepare a normal culture medium containing 5 mmol/L glucose, and 30 mmol/L high-glucose medium for use. MC was cultured at 5 mmol/L and 30 mmol/L, respectively. The high-glucose group was divided into five groups, model, pioglitazone (10µg/mL), low dose flavonoids of oxytropis falcata (LFOF) (10 µg/mL), medium dose flavonoids of oxytropis falcata (MFOF) (20 µg/mL), and high dose flavonoids of oxytropis falcata (HFOF) (40 µg/mL). Mycoplasma was detected before the experiment and the result was negative.
Assessment of ECM accumulation
The experimental method was based on Pugliese et al. 29 and was further improved. Media were collected and 10% carbonate–bicarbonate buffer, pH 9.6, protease inhibitors was added to prevent protein degradation, whereas cells were extracted with 0.5 N NaOH containing 2 mM phenylmethanesulfonyl fluoride (PMSF) and sonicated. The NaOH extracts referred to as cells actually included cells, intracellular spaces, and pericellular regions. Media and cell extracts were stored at −20°C until assayed C-IV, FN, and laminin.
Statistical methods
The Statistical Package for the Social Sciences (SPSS) 21.0 statistical software package was used for statistical analyses. The results are presented as the mean ± standard deviation. One-way analysis of variance was used to compare means among multiple groups, while a least significant difference t-test was used to compare two means of two groups. P < 0.05 was deemed statistically significant.
Results
Effect of total flavonoids from O. falcata on general conditions and changes in body weight in DN mice
Compared with the non-treat control group, after establishment of the diabetes model, the KK-Ay mice showed increases in food intake, water intake, urine output, and body weight. In control C57BL/6J mice, body weight increased steadily and the magnitude of the increase remained constant over the 4-week experimental period. Mice treated with total flavonoids from O. falcata gained weight at a significantly slower rate than model control mice. The mice administered the high dose gained weight more gradually than those given the low dose. The increase in body weight was the lowest in the pioglitazone-treated positive control group (Figure 1(a)). The results show that the effect of total flavonoids from O. falcata on weight control was dose dependent. The blood glucose and 24 h UAlb levels in the C57BL/6J control group were maintained at normal levels throughout the 4-week experimental period, whereas those in the DN model group were significantly increased significantly compared with this control group. Following treatment with total flavonoids from O. falcata, the increase in the blood glucose level induced by diabetes was suppressed in a dose-dependent manner (Figure 1(b)); the positive control was lower than the others. The increase in 24 h UAlb was also suppressed by treatment with total flavonoids from O. falcata, and the effect tended to be dose dependent. The increase in 24 h UAlb was the smallest in the pioglitazone-treated positive control group (Figure 1(c)).

Effects of total flavonoids from O. falcata on general conditions and changes in body weight, blood glucose, and UAlb. (a)The changes of body weight. (b) The changes of blood glucose. (c)The changes of UAlb.
Effects of total flavonoids from O. falcata on serum Cre, BUN, and UA in DN mice
Serum Cre, BUN, and UA levels in C57BL/6J control mice remained at normal levels over the 4 week experiment. Compared with the control group, the serum levels of Cre, BUN, and UA in the model mice were significantly (P < 0.01) increased 4 weeks after the model was established. Treatment with total flavonoids from O. falcata suppressed the increases in Cre, BUN, and UA, and the effects were significant (P < 0.05) in mice treated with the medium and high doses (Table 2).
Effects of total flavonoids from O. falcata on serum Cre, BUN, and UA.
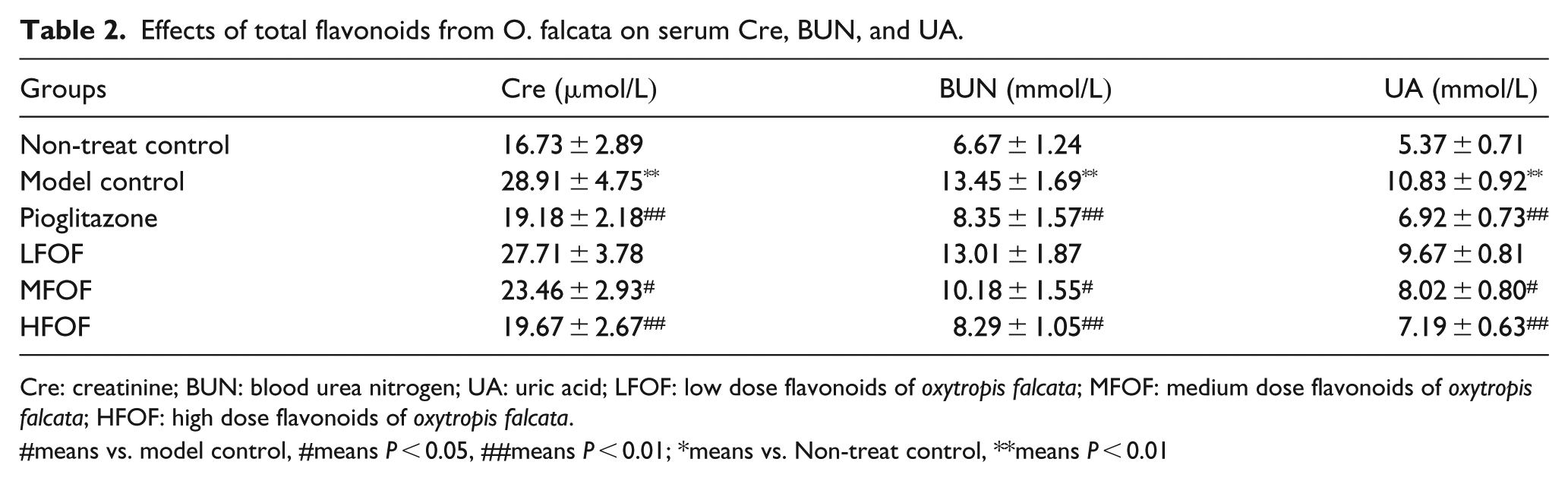
Cre: creatinine; BUN: blood urea nitrogen; UA: uric acid; LFOF: low dose flavonoids of oxytropis falcata; MFOF: medium dose flavonoids of oxytropis falcata; HFOF: high dose flavonoids of oxytropis falcata.
#means vs. model control, #means P < 0.05, ##means P < 0.01; *means vs. Non-treat control, **means P < 0.01
Effects of total flavonoids from O. falcata on glomerular histomorphology in DN mice
Compared with that in C57BL/6J control mice, the number of glomeruli in the kidney tissues of the model mice significantly decreased 4 weeks after model establishment, whereas pyknosis, glomerular cell number, and ECM significantly increased. Compared with that in the non-treated model mice, the number of glomeruli in the kidneys significantly increased in the pioglitazone-treated positive control group and the group treated with the high dose of total flavonoids from O. falcata for 4 weeks, whereas the number of glomerular cells and ECM significantly decreased in these two groups. Compared with the positive control group, there were no significant differences in the numbers of glomeruli and glomerular cells, and ECM in the renal tissues of the group treated with the high dose of total flavonoids from O. falcata (Figure 2). The glomeruli of the model group showed obvious vascular matrix dilation. With the increase of total flavonoids concentration, vascular matrix dilation decreased, and HFOF was not significantly different from the control group. The GMI score of the treatment groups was significantly lower than that of the model group.

Effects of total flavonoids from O. falcata on glomerular histomorphology. (a) H&E staining about renal tissues. (b) The analyse about glomerular histom.
Effects of total flavonoids from O. falcata on mRNA expression in the kidneys of DN mice and Glomerular mesangial cells
Compared with that in C57BL/6J control mice, renal mRNA levels of MCP-1, NF-κB, IL-6, and TGF-β1 significantly (P < 0.05 for all genes tested) increased in KK-Ay mice 4 weeks after model establishment. Compared with the non-treated model group, the increases in renal mRNA levels of all four genes tested were significantly (P < 0.05 for all genes tested) suppressed in mice treated with total flavonoids from O. falcata at week 4, at all doses tested (Figure 3(a)). In vitro experiments (Figure 3(b)), the results showed that there was no significant difference between the model group and positive group, but significantly when compared with blank control (P < 0.01). With the increase of the total flavonoids, the expression of JAK, STAT3, STAT4 decreased gradually, and the differences between the group were significant. However, SOCS-7 shows an opposite trend.
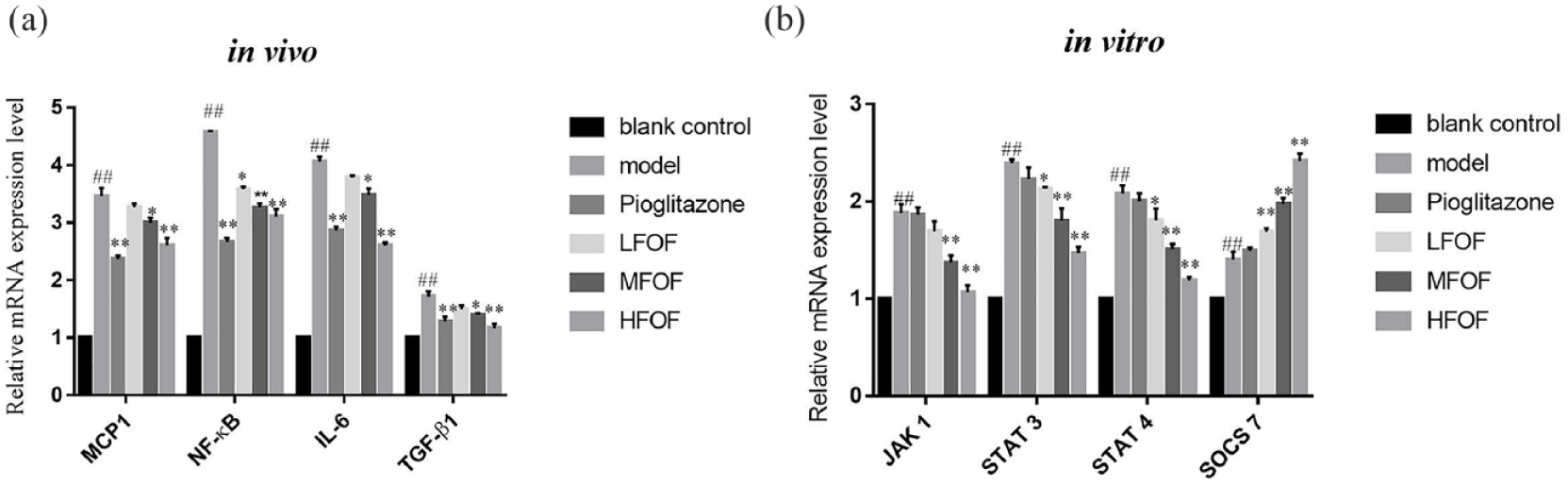
Effects of total flavonoids from O. falcata on mRNA expression. (a) The RNA espression level of cytokines in vivo. (b) Effect of total flavonoids on signaling pathway in vitro.
Effects of total flavonoids from O. falcata on the protein levels in the kidneys of DN mice and Glomerular mesangial cells
Compared with C57BL/6J control mice, p-JAK 2 and p-STAT 1 levels in the kidneys of the model group significantly increased 4 weeks after model establishment, whereas the SOCS-1 and SOCS-3 protein levels were significantly reduced. Compared with the model control mice, these responses were significantly suppressed in model mice treated with medium and high doses of total flavonoids from O. falcata at week 4, whereas the low dose had no significantly effects (Figure 4(a)). In cell experiments, TGF-β1, STAT3, p-STAT3 expression levels were the highest in the model group, and no significant difference with the positive control, and then decreased. Then, the expression of SOCS-2 was the least in the model group, and it increased in LFOF, MFOF, HFOF gradually (P < 0.01) (Figure 4(b)).
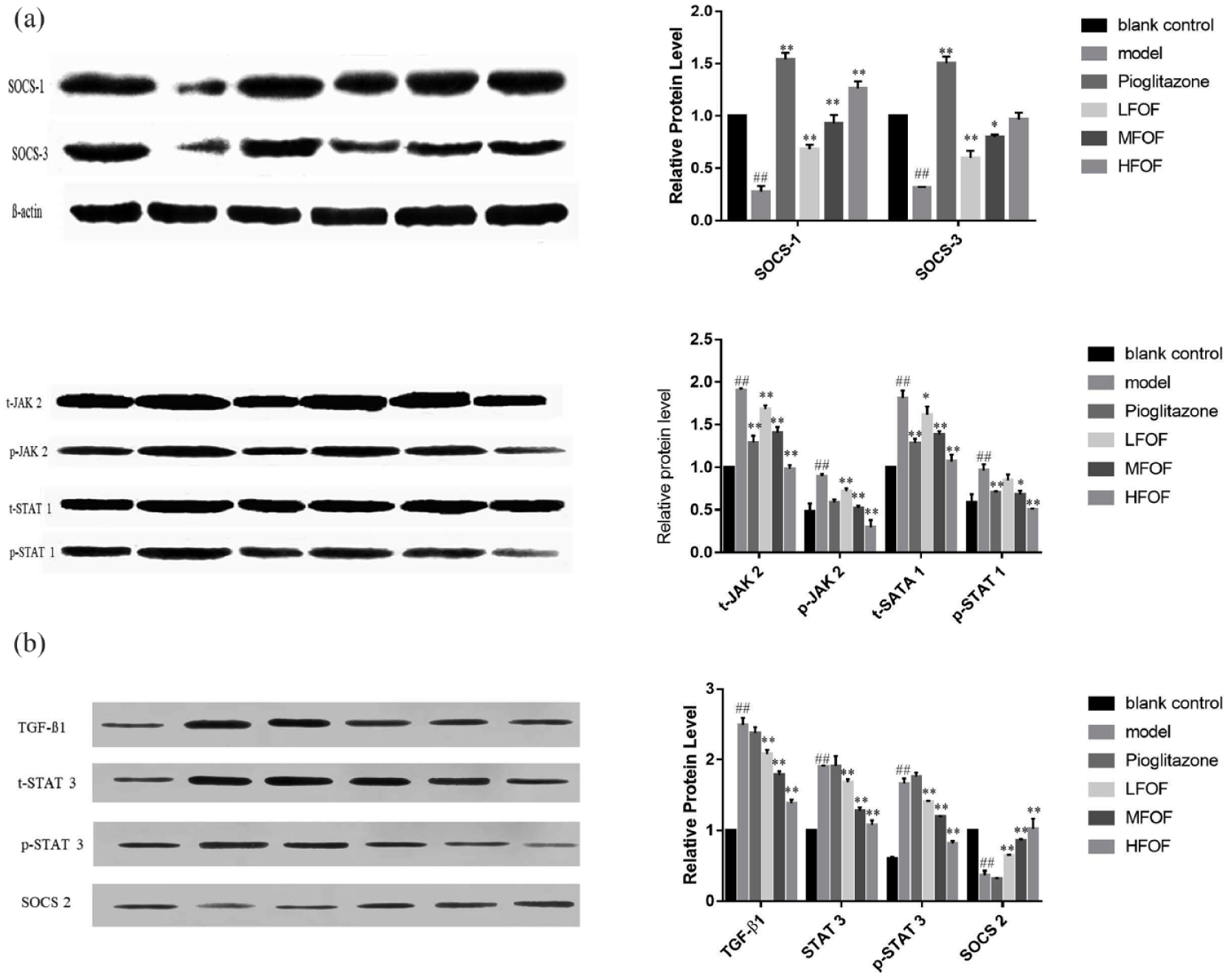
Effects of total flavonoids from O. falcata on the protein levels. (a) and (b) Efffect of total flavonoids on protein levels in kidneys. (c) Effect of total flavonoids on protein expression in glomerular mesangial cells.
The effect of total flavonoids from O. falcata on ECM accumulation in vitro
Compared with control group, model group of fibronectin, laminin, and collagen type IV expression on protein and RNA levels was significantly increased; positive control was no significant difference with model group. With the increase of total flavonoids dosage, the degree of renal mesangial cell fibrosis gradually decreased. Total flavonoids can reduce the accumulation of ECM, and it is speculated that total flavonoids act directly on renal cells to protect the kidney (Figure 5).

The effect of total flavonoid on ECM accumulation. (a) The western blot of Fibronectin, laminin and Collagen type. (b) The RNA expression level of Fibronectin, laminin and Collagen type.
Discussion
This study investigated the protective effect of total flavonoids from O. falcata on DN and the underlying molecular mechanisms using a KK-Ay mouse model. KK-Ay mice are Ay gene deficient, which renders them susceptible to obesity and gradual development of type 2 diabetes when they are fed a high-fat and high-glucose diet during growth. 30 In the present study, 8-week-old KK-Ay mice were fed a high-fat diet for 4 weeks. In C57BL/6 J control mice, blood glucose levels were normal throughout the experiment. Their body weight increased slightly over time but did not reach the criterium for obesity. In contrast, fasting blood glucose levels in KK-Ay mice were ⩾16.7 mmol/L, and their body weight increased significantly over 4 weeks of high-fat diet, suggesting that type 2 diabetes was successfully established.
Most patients with type 2 diabetes die from complications as their disease progresses, and DN, which is a key cause of death in these patients, is the major cause of end-stage renal disease. DN is a chronic microvascular disorder of the kidneys induced by high glucose. Pathological changes induced by DN include an increased glomerular filtration rate accompanied by mesangial proliferation, ECM thickening, glomerulosclerosis, and interstitial fibrosis of renal tubules. As DN progresses, the glomerular filtration rate continues to gradually decrease, accompanied by diffuse thickening of the glomerular basement membrane and matrix proliferation, and the formation of typical K-W nodules. This is followed by renal tubular atrophy and renal dysfunction and, eventually, end-stage renal disease.31,32
In the present study, 24 h UAlb levels in the KK-Ay type 2 DN model mice increased over time under high-fat diet. Blood levels of Cre, BUN, and UA, which are important indicators of renal functions,33,34 significantly increased at the end of week 4 after DN model establishment. Increases in UAlb, Cre, BUN, and UA indicate that the molecular sieve that serves as a barrier for glomerular filtration is disrupted. The negative charge in the glomerular filtration membrane is reduced, and the glomerular charge barrier and mechanical barrier are impaired. 35 In the present study, as the duration of high-fat and high-glucose diet feeding increased, the renal function of mice with type 2 diabetes gradually deteriorated, eventually leading to DN. Pathological changes in the renal tissues, including glomerular atrophy and ECM thickening, confirmed the development of DN.
Treatment of mice with type 2 diabetes with total flavonoids from O. falcata gradually decreased the fasting blood glucose with treatment duration, and the effect was dose dependent. The increases in body weight and 24 h UAlb gradually slowed down over time and in a dose-dependent manner. At the end of week 4 after model establishment, the blood concentrations of Cre, BUN, and UA in the treated groups decreased compared with those in the non-treated model group, and the treatment effect was dose dependent. Together, these results suggest that the total flavonoids from O. falcata can reduce blood glucose in mice with type 2 diabetes. The flavonoids had a protective effect on nephropathy caused by type 2 diabetes; they delayed the development and relieved the symptoms of DN. The protective effect of total flavonoids from O. falcata on the development of DN was confirmed by an improvement in the pathological changes observed in the renal tissues of the DN model mice.
Hyperglycemia is the underlying cause of DN, and direct control of the aberrant increase in blood glucose can effectively limit DN development. This may explain how total flavonoids from O. falcata can delay the onset of DN. A previous study showed that total flavonoids from O. falcata have a significant anti-inflammatory effect in addition to its ability to lower blood glucose, and inflammation is closely related to DN. 36 Therefore, we further investigated the effects of total flavonoids from O. falcata on the inflammatory factors and responses in the kidneys of the DN model mice. The results showed that total flavonoids from O. falcata reduced aberrant increases in kidney MCP-1, NF-κB, IL-6, and TGF-β1 mRNA levels induced by diabetes nephropathy. These four inflammatory factors reportedly play important roles in DN development and progression.11–13 They interact with multiple signaling pathways in the kidneys to form a complex network that promotes the development of DN. Among these, the JAK/STAT pathway is the main inflammation-related signaling pathway involved in DN development and progression.15–17 SOCS family proteins negatively regulate the JAK/STAT signaling pathway and inhibit its activation. 37 The present study revealed that total flavonoids from O. falcata reduced aberrantly elevated JAK2 and STAT1 protein phosphorylation and promoted aberrantly reduced SOCS-1 and SOCS-3 protein expression induced by diabetes nephropathy in the kidneys. This likely inhibited JAK/STAT pathway activation. Thus, total flavonoids from O. falcata inhibited the abnormal activation of the JAK/STAT pathway in the kidneys of the diabetic mice, thereby relieving inflammation. At the same time, to rule out of the protective effect through hypoglycemic effect of total flavonoids on renal, we conducted in vitro experiments and found that total flavonoids can inhibit the JAK/STAT signaling pathway. It may be due to the anti-inflammatory activity of flavonoids, it can be used as a drug to treatment inflammatory by inhibiting the inflammatory pathways.38,39 Activation of the JAK/STAT signaling cascade can stimulate excessive proliferation and growth of glomerular mesangial cells, contribute importantly to production of the cytokine as well as the ECM proteins collagen IV and fibronectin, leading to DN. 40 Thus, the modulation of this pathway may prevent or alleviate the progression of DN.
In conclusion, total flavonoids from O. falcata relieved inflammation in the kidneys in type 2 diabetes by reducing blood glucose and inhibiting JAK/STAT signaling, thereby exerting a protective effect on the development of DN. It provides a theoretical basis for the follow-up research on diabetes drugs. Furthermore, the results can provide some perspectives for future research.
Footnotes
Acknowledgements
Thanks to National natural science foundation (81560764), Gansu Longyuan Youth Innovation and Entrepreneurship Talent Project, Tibetan Medicine Regional Collaborative Innovation Center Project (2017XTCX013), and thanks to everyone who contributed to this article project.
Author’s Note
Lili Wu, Cuiyan Lv and Tonghua Liu are also affiliated with Tibetan University of Tibetan Traditional Medicine, Lhasa.
Author contributions
LY and JX are first co-authors; they contributed equally to the article. LW and CL are the corresponding authors; all authors read and approved the final manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval and consent to participate
This study was approved by the animal experiment ethics of Gansu University of Traditional Chinese Medicine. Experimental animals were killed and their bodies were disposed of: the cervical vertebra was dislocated after anesthesia with 3% pentobarbital sodium intraperitoneal injection. The carcasses were delivered to the animal center of Gansu University of Traditional Chinese Medicine for innocuous treatment at the hazardous waste disposal center of Gansu province (no. 2016-165).
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the National natural science foundation (81560764), Gansu Longyuan youth innovation and entrepreneurship talent project, Tibetan medicine regional collaborative innovation center project (2017XTCX013).