
Abstract
Multiple myeloma (MM) is a type of cancer characterized by abnormal proliferation of clonal cells; it is the very dangerous and highly prevalent disease. Although significant progress has been made in clinical research, especially with novel drugs such as bortezomib, lenalidomide, and carfilzomib, most of the patients with MM still suffer from often fetal relapses due to drug resistance. In this study, we aimed to develop immune cells that could specifically target and destroy MM cells. Chimeric antigen receptor–modified NK-92 (CAR-NK92) cells have been very effective against B-cell acute lymphoblastic leukemia (B-ALL); as MM shows high expression of CD138, we constructed CD138-directed CAR-NK-92MI cells (CAR-CD138). It 2is reported that there is a small subset of CD138–/CD19+ MM cells showing, to some extent, stem cell qualities. We therefore generated the CD19-directed CAR-NK-92MI cells (CAR-CD19) as well. These two CAR-NK cells showed strong in vitro biological activity in specifically killing target tumor cells. Thus, the concomitant use of these CAR-NK cells may achieve excellent results in vivo.
Introduction
Multiple myeloma (MM), the second most prevalent hematologic tumor, is a highly severe plasma cell malignancy, with a high morbidity rate and about 10% occupation rate among hematologic tumors. 1 In recent years, although some novel therapeutic methods have been developed, most patients with MM suffer from often fetal relapses.
Human natural killer (NK) cells can kill malignant cells in vivo; 2 furthermore, NK cells have been shown to be cytotoxic to MM cells.3,4 However, NK cells have certain drawbacks, which are not suitable for immune therapy, such as difficulty of proliferating, 5 which restrict their role in adoptive immunotherapy. NK-92 cells, derived from NK cells, are confirmed to be safe for clinical use in vivo. 6 Moreover, the proliferation of NK-92 cells is easy, compared to T cells that require a complex operation. 7 NK-92MI cells transduced with the cDNA of human interleukin (IL)-2 show smooth and fast proliferation, independent of additional IL-2. 8 A recent research study has shown that chimeric antigen receptor (CAR)-modified NK-92MI cells could kill MM cells effectively, with some success. 9
CD138 (syndecan-1), a specific marker of MM cells, is highly expressed in MM cell lines and plasma samples of patients with MM.10,11 Being a key marker associated with the growth and proliferation of MM cells, 12 CD138 is a very promising marker for the treatment of MM. CD138-directed first-generation CARs have been shown to be active in vitro; 9 furthermore, second-generation CARs have also been used. However, MM continued to show refractoriness, with patients suffering from relapses; this may be attributed to a small subset of CD138–/CD19+ MM cells that are similar to stem cells. Being low in proportion, the cells have not been verified to be stem cells as there are currently no feasible methods to perform this analysis. 13 So far, MM cell lines and patient samples have been found to have CD138– cells with low differentiation rates.14–16 These cells have been reported to be CD19+; they have been shown to have the properties of B cells and be associated with tolerance of drugs. 17 Although it is still not clear whether the CD138–/CD19+ cells are stem cells, there is a definite relationship between the proliferation of MM cells and the CD138–/CD19+ cells. 18 We constructed two kinds of CAR-NK-92MI cells to specifically target CD138 and CD19, respectively, and analyzed their cytotoxicity levels in vitro.
Materials and methods
Cell culture
Human mantle cell lymphoma cell lines MAVER-1 and JeKo-1, myeloma-derived cell lines RPMI8226 and U266, NK-92MI cells, and human erythroleukemic cell line, K562, were purchased from the American Type Culture Collection (Manassas, VA). NK-92MI cells were incubated in an alpha modification of Eagle’s minimum essential medium (MEM-α; Invitrogen, Carlsbad, CA) containing 2 mM L-glutamine and 1.5 g L−1 sodium bicarbonate. RPMI8226, U266, MAVER-1, JeKo-1, and K562 cells were cultured in complete media comprising RPMI 1640 (HyClone), 10% fetal bovine serum (Biowest, FRA), 100 U mL−1 penicillin, and 100 μg mL−1 streptomycin. HEK293T cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM; HyClone) containing 10% fetal bovine serum. All cells were cultured at 37°C in a humidified atmosphere with 5% CO2.
Construction of CD138-CAR and CD19-CAR
The CAR-CD138 and CAR-CD19 cells were constructed as second-generation CARs. Specifically, anti-CD138 scFv or anti-CD19 scFv were inserted into the lentiviral vector pCDH-CMV-MCS-EF1-CopPuro. Subsequently, the Fc domain, costimulatory CD28 cytoplasmic domain, and the CD3ζ region were attached in sequence to form the desired second-generation CAR.
Lentivirus production and NK-92MI cell transduction
Lentivirus production was performed as described previously. 19 NK-92MI cells (1 × 106) were co-cultured with lentiviral supernatants (multiplicity of infection (MOI) = 15) for 6 h in a 48-well culture plate. The supernatant was removed and the cells were placed in a 10-cm dish followed by adding 7 mL MEM-α. After 72 h, the NK-92MI cells stained with allophycocyanin (APC)-conjugated anti-Fc mAb were assessed using flow cytometry (FACSCalibur; BD Biosciences, NJ). The Fc-positive NK-92MI cells were sorted with FACSAriaTM III Cell Sorter (BD Biosciences, NJ) and puromycin (Thermo Fisher Scientific, MA).
Construction of K562-CD138 and K562-CD19 cells
To validate the specificity of these two CAR-NK cells to their targets, we constructed K562 cells expressing the CD138 antigen (K562-CD138) and CD19 antigen (K562-CD19), as described previously. 19 The CD138 (NM_002997) and CD19 (AH005421.2) sequences were retrieved from National Center for Biotechnology Information (NCBI), processed using the SignalP 4.1 Server and UniProt, respectively, inserted into pCDH-CMV-MCS-EF1-CopGFP vectors and packaged lentiviruses. After transduction for 6 h and subsequent culturing, the K562 cells were stained with their corresponding antibodies and sorted using the FACSAriaTM III cell sorter (BD Biosciences) to obtain a positive rate of 100%.
Flow cytometry
The RPMI8226, U266, and K562-CD138 cells were stained with FITC-conjugated anti-CD138 antibodies (BD Biosciences), while the MAVER-1, JeKo-1, and K562-CD19 cells were stained with FITC-conjugated anti-CD19 antibodies. Specifically, the cells were incubated at 37°C for 15 min, washed three times with phosphate-buffered saline (PBS), and then analyzed using a FACSCalibur flow cytometer.
In vitro killing experiment
The in vitro killing experiment was performed by measuring lactate dehydrogenase (LDH) released by target cells using the CytoTox 96® Non-Radioactive Cytotoxicity Assay (Promega, Madison, USA) in strict adherence to the manufacturer’s instructions. We used four groups with the E:T ratios of 0.25:1, 0.5:1, 1:1, and 2:1. Specifically, 4 × 105 target cells were incubated with the corresponding amount of CAR-NK cells for 6 h in a 24-well plate, and the volume of the final culture system was 1.5 mL. In total, 50 µL of supernatant from each group was placed in a 96-well plate, and 50 μL of CytoTox 96 reagent was then added to each well. After 30 min, 50 μL of stop solution was added to each well and the reaction time was 1 h. The absorbance was recorded for each sample at 490 nm using a microplate reader. The killing rate was calculated using the following formula

To verify the results of this experiment, we used the second method to test the cytotoxicity of CAR-CD138 to MM cells, as described previously. 19 The target cells were resuspended in PBS and treated with 1 μL carboxyfluorescein succinimidyl ester (CFSE) at 37°C for 10 min. The target cells (4 × 105) were then placed in a 24-well plate, the corresponding number of CAR-NK cells were added to each plate, and the contents were mixed thoroughly. The final volume of the culture system was 1.5 mL. After reacting for 6 h, the supernatant was removed, the cells were resuspended in PBS and 1 μL 7-aminoactinomycin D (7-AAD) was added. The killing rates were then analyzed using a FACSCalibur flow cytometer. The CFSE-positive cells were the target cells, and the percentage of 7-AAD-positive cells was the death rate of the target cells.
Data analysis
For the in vitro experiments, the data were processed using GraphPad Prism 5 and Social Sciences 23.0 (SPSS Inc., Chicago, IL, USA). The experiments were repeated at least three times for each group, and statistical analysis was performed using Student’s t test, with P values < 0.05 indicating significance.
Results
Construction of CAR-NKs and detection of CD138 and CD19 antigens on the target cell surface
First, we constructed CAR-NK-92MI cells (CAR-CD138 and CAR-CD19), the second-generation CARs, to target the CD138 and CD19 antigens, respectively (Figure 1(a)). The cDNA sequences of CD138 and CD19, obtained from the PersonGen BioTherapeutics (Suzhou) Co., Ltd, consisted of heavy and light chains, with a signal peptide sequence (MLLLVTSLLLCELPHPAFLLIP). The scFv regions of anti-CD138 and anti-CD19 were inserted into a lentiviral vector, pCDH-CMV-MCS-EF1-CopPuro, along with the Fc domain, the CD28 costimulatory region, and the CD3ζ region. After transduction, positive CAR-NK cells were sorted and the positive rate was verified to be above 99% (Figure 1(b)).

Construction of CAR-NKs and detection of antigens. (a) CAR-CD138 and CAR-CD19 were second-generation CARs, which were inserted into the lentiviral vector pCDH-CMV-MCS-EF1-CopPuro, with the restriction enzyme cutting sites XbaI and NotI. (b) The detection of positive rate of CAR-NKs. The transduced NK-92MI cells were collected and stained with antibodies and then sorted. Several days later, we detected their positive rates. (c) To test the expression level of CD138 antigen in MM cell lines RPMI8226 and U266 cells and that of CD19 antigen in MCL MAVER-1 and JeKo-1 cells. (d) To test the expression level of CD138 antigen on the surface of K562-CD138 cells and that of CD19 antigen on the surface of K562-CD19 cells, as well as the antigen of parent K562 cells.
We also detected the expression of antigens on the surface of MM and lymphoma cells. Both RPMI8226 and U266 cells substantially expressed the CD138 antigen, with the high expression levels of 96% and 99%, respectively. Because of the extremely low expression of CD19 on the surface of MM cells, we selected two lymphomas, MAVER-1 and JeKo-1, as the targets to verify the cytotoxicity of CAR-CD19 cells. The expression levels of MAVER-1 and JeKo-1 were 92% and 80% (Figure 1(c)), respectively, demonstrating that they were suitable to be used as the target cells.
Furthermore, we constructed K562 cells stably expressing CD138 and CD19 antigens (K562-CD138 and K562-CD19, respectively), to assay the specificity of the killing ability of CAR-CD138 and CAR-CD19 cells. The parent K562 cells are CD138–/CD19– (Figure 1(d)), and the expression levels of these two antigen-modified K562 cells were 96% and 97%, respectively.
CAR-NKs show enhanced cytotoxicity to tumor cells
The target cells of CAR-CD138 cells were RPMI8226 and U266 cells. CAR-CD138 cells showed obviously higher cytotoxicity to RPMI8226 cells than the parent NK-92MI cells. The death rate in the experimental group was about twice that in the control group when the E:T ratio was 0.25:1. When the E:T ratio was 2:1, the death rate in the experimental group was 80%, while that in the control group was 50% (Figure 2(a)). For U266 cells, the killing rate by CAR-CD138 was much higher than by NK-92MI cells (Figure 2(b)). The target cells of CAR-CD19 were MAVER-1 and JeKo-1 cells. For MAVER-1 cells, the killing rate by CAR-CD19 was several times than that by the control group, with all four E:T ratios. Especially with an E:T ratio of 0.25:1, the death rate in the experimental group was five times more than that in the control group (Figure 2(c)). For an E:T ratio of 0.5:1, the killing rate of JeKo-1 cells by CAR-CD19 cells was five times more than that by NK-92MI cells (Figure 2(d)).

The killing ability of CAR-NKs toward tumor cells. (a) 4 × 105 target cells were collected, followed by the addition of CAR-CD138 cells, and 6 h later the supernatant was collected to test the killing ability toward RPMI8226 (a) and U266 (b) cells using Cytometric Bead Array (CBA) by flow cytometry. (c) The killing detection of CAR-CD19 against mantle cell lymphoma MAVER-1 (c) and JeKo-1 (d) was performed. Data presented are the mean ± SD of the three separate experiments. *P < 0.05, **P < 0.01, ***P < 0.001 compared with NK-92MI cells at the same E:T ratio.
To further assay the specificity of the killing ability of CAR-NKs toward CD138+ and CD19+ cells, we constructed the K562-CD138 and K562-CD19 cells, respectively. The time of incubation and groups were retained as in the previous experiment. The results indicated that the killing ability of CAR-CD138 cells toward K562-CD138 cells was evidently higher than that of NK-92MI cells, while the death rates induced by CAR-CD138 were almost the same as that of the controls for the parent K562 cells (Figure 3(a) and (c)), demonstrating that the constructed CAR-NK cells were highly specific toward their target cells. Similar results were obtained for the CAR-CD19 cells with the K562-CD19 cells (Figure 3(b) and (d)).

Specificity of CAR-CD138 and CAR-CD19 cells to CD138 and CD19 antigens, respectively. (a) To test the killing ability of CAR-CD138 cells toward K562-CD138 (a) and parent K562 (c) cells. (b) To test the killing ability of CAR-CD19 cells toward K562-CD19 (b) and parent K562 (d) cells. 4 × 105 target cells were collected, followed by the addition of CAR-NK cells, and then the LDH released by target cells were tested. Data presented are the mean ± SD of the three separate experiments. *P < 0.05, **P < 0.01, ***P < 0.001 compared with NK-92MI cells at the same E:T ratio.
CAR-CD138 cells assuredly kill MM cells
Next, we verified the cytotoxicity of CAR-CD138 cells toward MM cells using a FACSCalibur flow cytometer. Target cells stained by CFSE were co-cultured with CAR-CD138 cells for 6 h and then treated with 7-AAD. The results revealed that the death rate of MM cells was higher than that of the control group, for RPMI8226 and U266 cells. Although the difference was not very large between the experimental and control groups, it was significant (Figure 4(a) and (b)). For example, the death rate was 90% in the experimental group and 60% in the control group for the U266 dells, with an E:T ratio of 1:1.
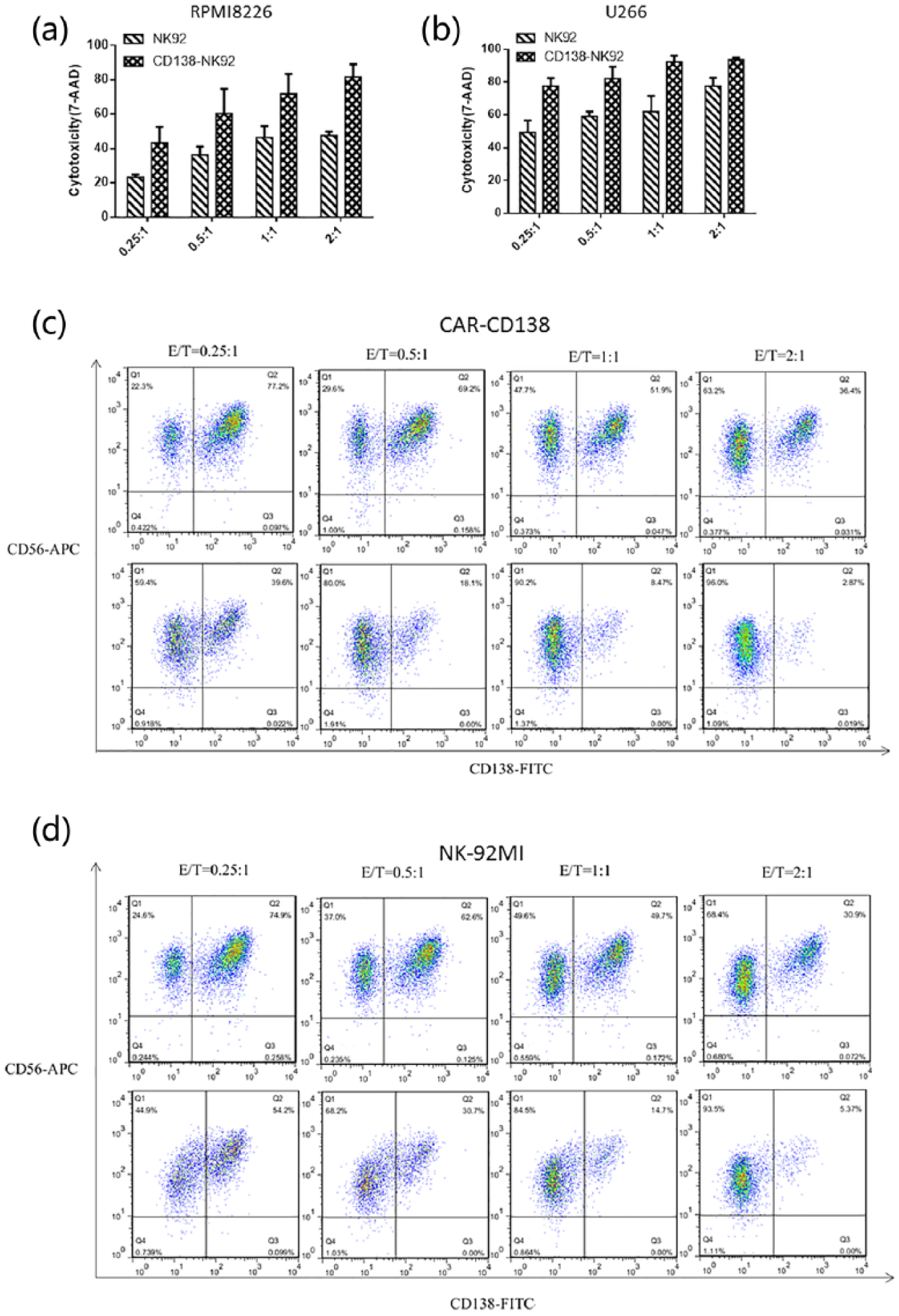
Verification of the cytotoxicity of CAR-CD138 toward MM cells. (a) 4 × 105 target cells stained by CFSE were collected, followed by the addition of CAR-CD138 cells, and 6 h later 7-AAD was added and the detection of the quantity of dead RPMI8226 (a) and U266 (b) was performed by flow cytometry. Data presented are the mean ± SD of the three separate experiments. (C) The killing rates of CAR-CD138 (c) and NK-92MI (d) toward RPMI8226 cells were analyzed individually. RPMI8226 cells were mixed with effector cells followed by staining anti-CD56 and anti-CD138 antibodies, and 6 h later the detection was performed by flow cytometry.
We then mixed RPMI8226 cells with CAR-CD138 and NK-92MI cells in their corresponding proportions and stained with FITC-conjugated anti-CD138 antibodies and APC-conjugated anti-CD56 antibodies, respectively. Before co-culture, we analyzed the proportion of these cells on a flow cytometer. After 6 h, we analyzed their proportions again. The results indicated that, for CAR-CD138 cells, RPMI8226 cells reduced by 48.7%, 73.8%, 87.5%, and 92.1% with the E:T ratios of 0.25:1, 0.5:1, 1:1, and 2:1 (Figure 4(c)), while the target cells reduced by 27.5%, 50.9%, 70.0%, and 81.5%, respectively, for the NK-92MI cells (Figure 4(d)).
Thus, CAR-CD138 indeed was highly cytotoxic toward MM cells.
Discussion
MM is a prevalent hematological malignancy characterized by the release of abnormal immunoglobulins. Traditional chemotherapy and radiotherapy treatment have great drawbacks, while hematopoietic stem cell transplantation can easily trigger graft-versus-host disease. Although some immunomodulators have been recently developed, tolerance remains a challenging obstacle. CAR-T, a type of emerging immunotherapy, is a promising method for overcoming tumor cells. The scFv of CAR can recognize tumor-associated antigen, independent of MHCs; furthermore, the antigen is diverse.20,21 CAR-T cell therapeutic product, CTL019, produced by Novartis (USA), the first gene-modified cells, showed significant success in preclinical studies. It has also been clinically approved targeting CD19 on the surface of hematologic tumors. In this study, we adopted a new strategy using CAR-NK technology as a therapy against MM cells. We constructed CAR-CD138 and CAR-CD19 cells and used them concomitantly to target CD138 and CD19 antigens on the surface of MM cells. MM cell lines and the bone marrow plasma cells of patients with MM are known to highly express CD138 antigen.10,11 Another marker, CD38, which is highly expressed on MM cells, may have more potential; however, it is also expressed in T and B cells, making it difficult for CAR-NK cell therapy to distinguish MM cells from T and B cells.22,23 As CD138 is target associated with the growth and proliferation of MM cells and can promote tumor angiogenesis, it is a more suitable target.
An experimental constraint we encountered in this study was a lack of CD138–/CD19+ MM cells. The fraction of CD19+ cells among the primary MM cells is extremely low, 11 and their detection through routine method is very difficult. Lin et al. 11 analyzed bone marrow aspirates from hundreds of patients with MM using flow cytometry and found that 1% of the MM cells were CD19+. Gupta et al. 24 analyzed bone marrow aspirates of 103 MM patients and found that 3.7% of the MM cells were CD19+. Therefore, to verify the activity of our CAR-CD19 cells, we used MAVER-1 and JeKo-1 cells, which are CD19+ mantle cell lymphoma. Another limitation of this study is that we did not perform an in vivo experiment of the concomitant use of these two CAR-NKs.
Our results reveal a promising method of therapy against MM, and further studies should focus on testing both these in vitro and in vivo. Our study takes us a step closer to obtaining an efficient treatment for MM.
Footnotes
Acknowledgements
S.Z. performed the experiments, acquired and interpreted the data, and wrote the manuscript. Z.H. was involved in the construction of plasmid. C.J., H.M., and G.A. cultured some cells. L.Y. designed the experiments and gave an elaborate guidance for this project.
Declaration of conflicting interests
L.Y. is a co-founder of PersonGen BioTherapeutics Co., Ltd. which focuses on R&D of CAR-T and CAR-NK technologies and translating them to clinics. There is no other potential conflict of interest to disclose.
Funding
This work was supported by the Priority Academic Program Development of Jiangsu Higher Education Institutions, the National Natural Science Foundation of China (Grant Nos 31471283 and 81201861), the Collaborative Innovation Major Project (Grant No. XYXT2015304), the Six Talent Peaks Project in Jiangsu Province (No. SWYY-CXTD-010), and Shandong Provincial Natural Science Foundation (Grant No. ZR2015PH029).