
Abstract
The aim of this article is to identify the relationship between the nuclear factor κB (NF-κB) signal pathway expression and the anastomotic stricture. It particularly focuses on the expression of p65, bcl-2, and cIAP-1 in the anastomotic stricture formation after radical resection of esophageal cancer. A total of 82 esophageal squamous carcinoma patients who had undergone esophagectomy by Ivor Lewis procedure were enrolled in the study. Tissues were collected from the patients who developed the anastomotic stricture after the operation, while para anastomotic tissues served as the control. The protein expression of p65, bcl-2, and cIAP-1 was analyzed by immunohistochemistry and western blot analyses, while the messenger RNA (mRNA) levels of p65, bcl-2, and cIAP-1 were evaluated by reverse transcription polymerase chain reaction (RT-PCR). The results showed that lower expression of p65, bcl-2, and cIAP-1 proteins was observed in the para anastomotic tissue; in the esophageal stricture’s tissue, the expression of these proteins was significantly higher (P < 0.05). The mRNA levels of P56, bcl-2, and cIAP-1 in the stricture tissue were remarkably increased (P < 0.05) compared with the para anastomotic tissues, and the mRNA levels in the sample of grade 3 dysphagia were higher (P < 0.05) than the levels of grade 1 and 2. In the normal esophageal epithelial cell of stricture patients was upregulated compared with that of no stricture patients. We can confirm that the anastomotic stricture has the relationship with the NF-κB signal pathway. The bcl-2 and cIAP-1, located downstream of NF-κB signal pathway, could be a new preventive and therapeutic target for the anastomotic stricture.
Introduction
The incidence of esophageal carcinoma has grown remarkably over the past 50 years, and the squamous cell carcinoma is the most common type in China. Surgical operation achieves the aim of complete excision of the lesion and regional lymph node and remains the preferred therapy for esophageal cancer. However, surgery is the double-edged sword of the treatment due to the high morbidity and mortality rates.1,2 Anastomotic stricture is one of the primary complications in the esophagectomy.3,4 Most benign strictures occur within the first few months after the operation, and dysphagia again is often the first symptom indicating the presence of the stenosis, which will bring tremendous pain to the patients. 5 Although with the development of the surgical technique, the incidence of the anastomotic stricture has declined; it is still about 20%–30%, much higher than expected. 3 Once the anastomotic stricture occurs, there are not very effective treatment procedures. Dilation and drug therapy are the most frequent intervention, but recurrence rate remains high.1,2 Apparently, mastering the whole mechanism of the stenosis is extremely important, which, however, is unclear so far.
Nuclear factor κB (NF-κB) is a transcriptional factor first identified by Sen and Baltimore in 1986. NF-κB’s activation relates to the pathogenesis of various diseases,6,7 especially to the tumor development and progression. The signal pathway of NF-κB has the effect of anti-apoptosis and can be activated by inflammatory, autoimmune diseases, infection, and cancer. 8 When the patient undergoes esophagectomy, the anastomosis would expose to the acid, inflammatory, and other factors, thus activating the NF-κB signal pathway. As we know, fibroblast plays a very crucial role in the anastomotic repairment. Messadi et al.’s 9 paper has reported that the expression levels of activated NF-κB in the keloid fibroblasts are apparently higher than those in healthy skin.
To our knowledge, few papers investigate the effects of NF-κB expression in the process of anastomotic stricture after the radical resection of esophageal carcinoma. So, this study aims to find the relationship between the NF-κB signal pathway expression and the anastomotic stricture, particularly focusing on the p65, bcl-2, and cIAP-1. It may provide a basis for the therapies of esophageal anastomotic stricture.
Materials and methods
Study population
This study was approved by the institutional review board of the ethics committee of The First Hospital of Jilin University, and all authors had access to the study data. Between September 2014 and December 2015, 82 esophageal squamous carcinoma patients (78 males and 4 females) who had underwent esophagectomy by Ivor Lewis procedure at The First Hospital of Jilin University were enrolled in the study. All of them were first treated by surgery in our hospital and did not develop the anastomotic leakage or receive the radiotherapy after the operation. The average age of the subjects was 59.3 ± 6.7 years. In these patients, 51 cases had mid-esophageal cancer and 31 had lower-esophageal cancer. About 32 cases were found with recurrent benign anastomotic strictures. Severity of dysphagia was assessed according to Stooler’s score system. Grade 1—the participant could have soft food, grade 2—the participant was able to eat semi-liquid food, grade 3—the participant was able to eat liquid food, and grade 4—the participant could not take food by themselves. In all, 17 patients were in grade 2 (13 mid-esophagus and 4 lower esophagus), 12 patients were in grade 3 (9 mid-esophagus and 3 lower esophagus), and 3 patients in grade 4 (all were mid-esophagus). All of them were diagnosed by endoscopy examination. Meanwhile, 50 participants without dysphagia after esophagectomy were enrolled in the control group. We also recorded the pathological stage of the patients. The basic features of the samples are shown in Table 1.
Characteristics of patients in this study.

Tissue samples
We obtained the written consent forms from all patients in this study. Before the operation, we collected the normal and abnormal mucosal tissues when the patient received the gastroscopy examination for diagnosing the cancer. After the surgery, if the patient developed dysphagia, we performed Gastrografin esophagography first to diagnose the anastomotic stricture and then checked the gastroscopy examination to identify the diagnosis. After that, we collected the tissue samples in the anastomotic stricture area and para anastomosis by biopsy forceps, with the size of about a grain of rice. After fixing all the specimens with 40-g/L neutral buffered formalin, we dehydrated and embedded the samples into paraffin. Meanwhile, a part of each sample was snap-frozen and stored in the liquid nitrogen for subsequent quantitative reverse transcription polymerase chain reaction (RT-PCR) analysis.
Immunohistochemistry
Immunohistochemistry (IHC) staining was uses to test the levels of P56, bcl-2, and cIAP-1 proteins in tissue samples. Briefly, we deparaffinized paraffin-embedded tissue sections in xylene and rehydrated tissue section in a series of ethanol solutions (100% to 50%). Then, we incubated the tissue sections in 0.3% H2O2 for 15 min to block endogenous peroxidase activity. Next, the samples were washed three times in the phosphate-buffered saline (PBS) buffer for 5 min. Antigen retrieval was done by boiling in a Tris/ethylenediaminetetraacetic acid (EDTA) (pH 9.0) solution for 20 min. The mixtures were incubated in normal serum at 26°C for 30 min. The primary antibodies against P56 (Cell Signaling Technology, Danvers, MA, USA; 1:400 dilution in PBS), BCL-2 (Cell Signaling Technology; 1:500 dilution in PBS), and cIAP-1 (Cell Signaling Technology; 1:200 dilution in PBS) were added and incubated at 4°C overnight. The next morning, after the tissues were washed three times with PBS buffer, the secondary antibody through the PV-9001 IHC kit was added. The color reaction was finished with a 3′-diaminobenzidine kit (Zhongshan Golden Bridge Biotechnology LLC, Beijing, China). Tissues were cut into 5-μm-thick slices, dewaxed, and dehydrated. Endogenous peroxidase activity was quenched by 3% H2O2. Through observation under microscope, we randomly chose six fields of each section. The positive signs of the proteins were brown granules in the cytoplasm, and the signals were classified according to their color: light yellow signaled weak staining and were recorded as “+”; brown was considered as moderate staining and thus were recorded as “++”; chocolate brown signaled strong staining and were recorded as “+++.”
Western blot analysis
P56, bcl-2, and cIAP-1 antibodies were purchased from Abcam (Cambridge, MA, USA). A sample weighted 200 mg was sectioned into small pieces and lysed with 1-mL radio immunoprecipitation assay (RIPA) lysis buffer (Roche Applied Science, Penzberg, Germany) and 10 μL PMSF (Roche Applied Science) buffer. The concentration of protein was determined by bicinchoninic acid (BCA) assay kit (Beyotime, Shanghai, China). Then, equal amounts of total proteins were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). After blocking with 5% nonfat milk, the primary antibodies were added and incubated at 4°C overnight. The next morning, the proteins were transferred to filter paper membranes; then, the appropriate rabbit monoclonal antibody for human P56, bcl-2, and cIAP-1 was added. The membranes were incubated in a blocking buffer (0.1 mL/cm2) on a shaker at 4°C. Then, the horseradish peroxidase (HRP)-conjugated secondary antibodies diluted in a blocking buffer were added and incubated with the membranes for 1–2 h at room temperature. After that, PBST (phosphate-buffered saline with Tween 20) was used to wash the membranes for 5 min and four times.
RNA extraction and amplification by RT-PCR
The TRIzol reagent was used for isolation of the RNA from the tissues according to the manual. The RNA quality was detected by checking 260/280 nm absorbance, and a ratio of 1.8–2.1 was considered to be qualified nucleic acid purity. M-MLV reverse transcriptase (Takara, Tokyo, Japan) was used to amplify 0.5 μg of total RNA to the final complementary DNA (cDNA) at 42°C for 50 min. The PCR cycles were set as 95°C for 3 min; then, 35 cycles were set under the following condition: 50 s at 94°C, 30 s at 55°C, and 60 s at 72°C. After that, the final mixtures were incubated at 72°C for 3 min. Primers were synthesized and purified by Takara (Dalian, China): bcl-2 (F, 5′-GGATCCAGGATAACGGAGGC-3′; R, 5′-GGGCCAAACTGAGCAGAGTC-3′), cIAP-1(F, 5′-GGCCGTATCTCCTTGTCGG-3′; R, 5′-TGCAGGGGGACAAAATAGGG-3′), and P56 (F, 5′-AACTAAGAGCACTGGGAGGCAT-3′; R, 5′-GTGCAAGAGCATCATCCAGTT-3′). Then, the ratios of density from the UVPro color image analysis system were calculated.
Statistical analysis
SPSS 17.0 software was used to analyze all the data which were presented as the mean ± standard error of the mean (SEM). We performed statistical comparison using t test and Wilcoxon two-sample test. The P value of <0.05 was considered as statistically significant.
Results
Patient characteristics are shown in Table 1. A total of 32 patients who developed esophageal stricture and 50 no stricture controls were included in this study. In terms of the general characteristics between the two groups, we observed no significant difference among age, gender, and cancer location. About 17 cases were of grade 2 dysphagia and 12 cases were of grade 3 dysphagia. Only three cases were of grade 4 dysphagia. The anastomotic stricture developed most often in the pathological stage III.
Protein level of P56, bcl-2, and cIAP-1 examined by IHC
P56, bcl-2, and cIAP-1 proteins in the tissues were detected by IHC. Lower expression of P56, bcl-2, and cIAP-1 proteins was observed in the para anastomotic tissue (Figure 1), while in the esophageal stricture’s tissue, significant increase in protein levels of P56, bcl-2, and cIAP-1 was detected (P < 0.05). Most of these proteins were located in the cytoplasm of the cells. The positive expressions of P56, bcl-2, and cIAP-1 in the cells were indicated by the brown-yellow granules (Table 2).

Proteins of P56, bcl-2, and cIAP-1 in the stricture tissue and para anastomotic tissue (IHC ×200).
Expression of P56, bcl-2, and cIAP-1 in the stricture tissue and para anastomotic tissue.

Messenger RNA levels of P56, bcl-2, and cIAP-1 in the anastomotic and para anastomotic tissues
We further evaluated the messenger RNA (mRNA) levels of P56, bcl-2, and cIAP-1 by RT-PCR. The results showed that the mRNA levels of P56, bcl-2, and cIAP-1 in the stricture tissue were significantly increased (P < 0.05) in comparison with the para anastomotic tissue (Figure 2). Meanwhile, the mRNA levels in the samples of grade 3 dysphagia were significantly raised higher (P < 0.05) than those in the grade 1 and 2 dysphagia. At the same time, the mRNA levels of P56, bcl-2, and cIAP-1 in the normal esophageal epithelial cell of stricture patients were upregulated compared with that of no stricture patients.
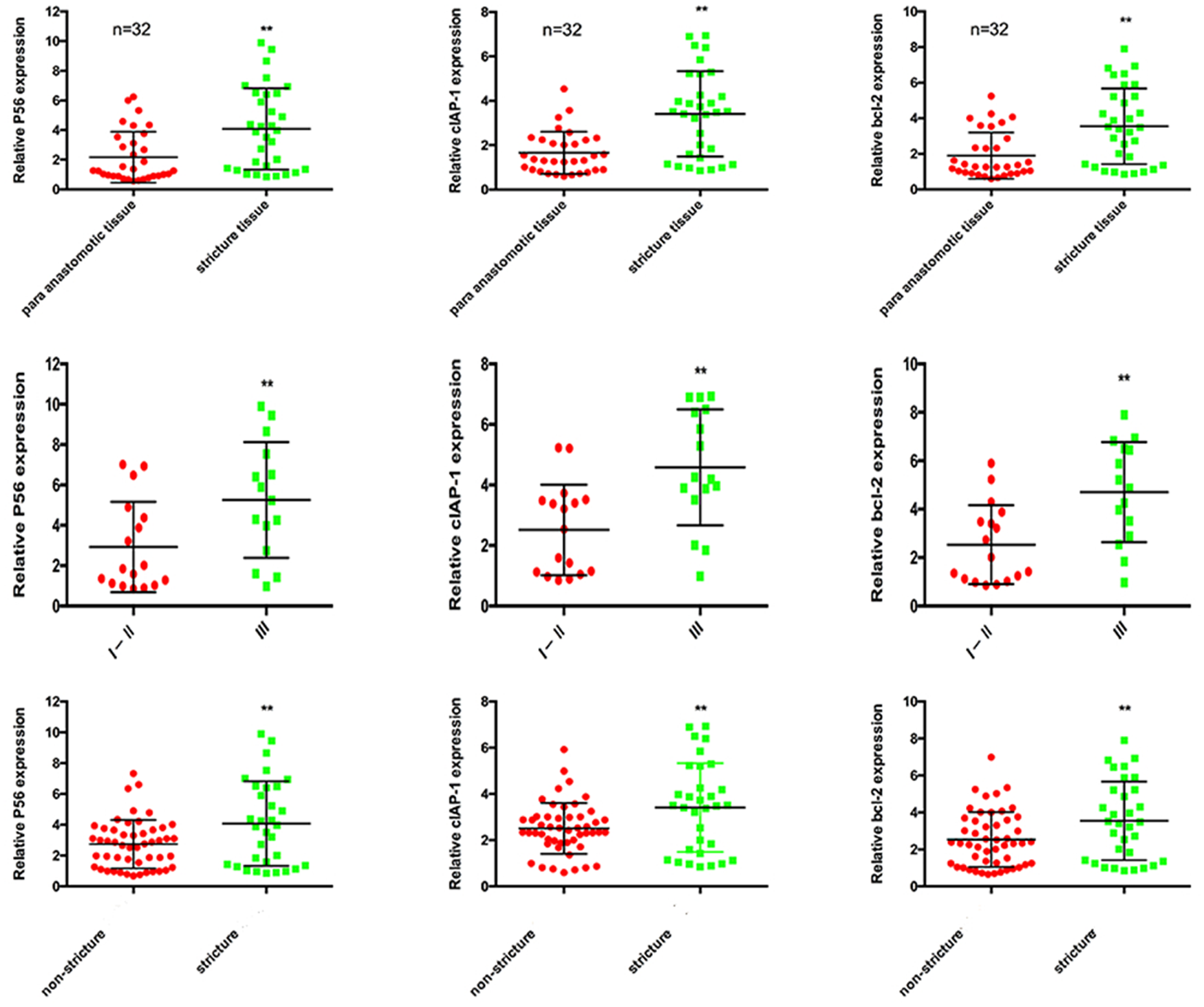
Line 1 showed the mRNA levels of P56, bcl-2, and cIAP-1 in the para anastomotic tissue and stricture tissues. The mRNA levels were upregulated in the stricture tissue apparently (P < 0.05). Line 2 showed the mRNA levels of P56, bcl-2, and cIAP-1 in tissue of grade 1 and 2 dysphagia patients and grade 3 dysphagia patients. The sample’s mRNA was upregulated in the grade 3 dysphagia patients apparently (P < 0.05). Line 3 showed the mRNA levels of P56, bcl-2, and cIAP-1 mRNA in the normal tissue of non-stricture patients and stricture patients. The sample’s mRNA was upregulated in the stricture patients apparently (P < 0.05).
Protein levels of P56, bcl-2, and cIAP-1 detected by western blot analysis
P56, bcl-2, and cIAP-1 proteins were detected by western blot analysis. As shown in Figure 3, significant raised levels of P56, bcl-2, and cIAP-1 proteins were observed compared with those from the para anastomotic tissues (P < 0.05).
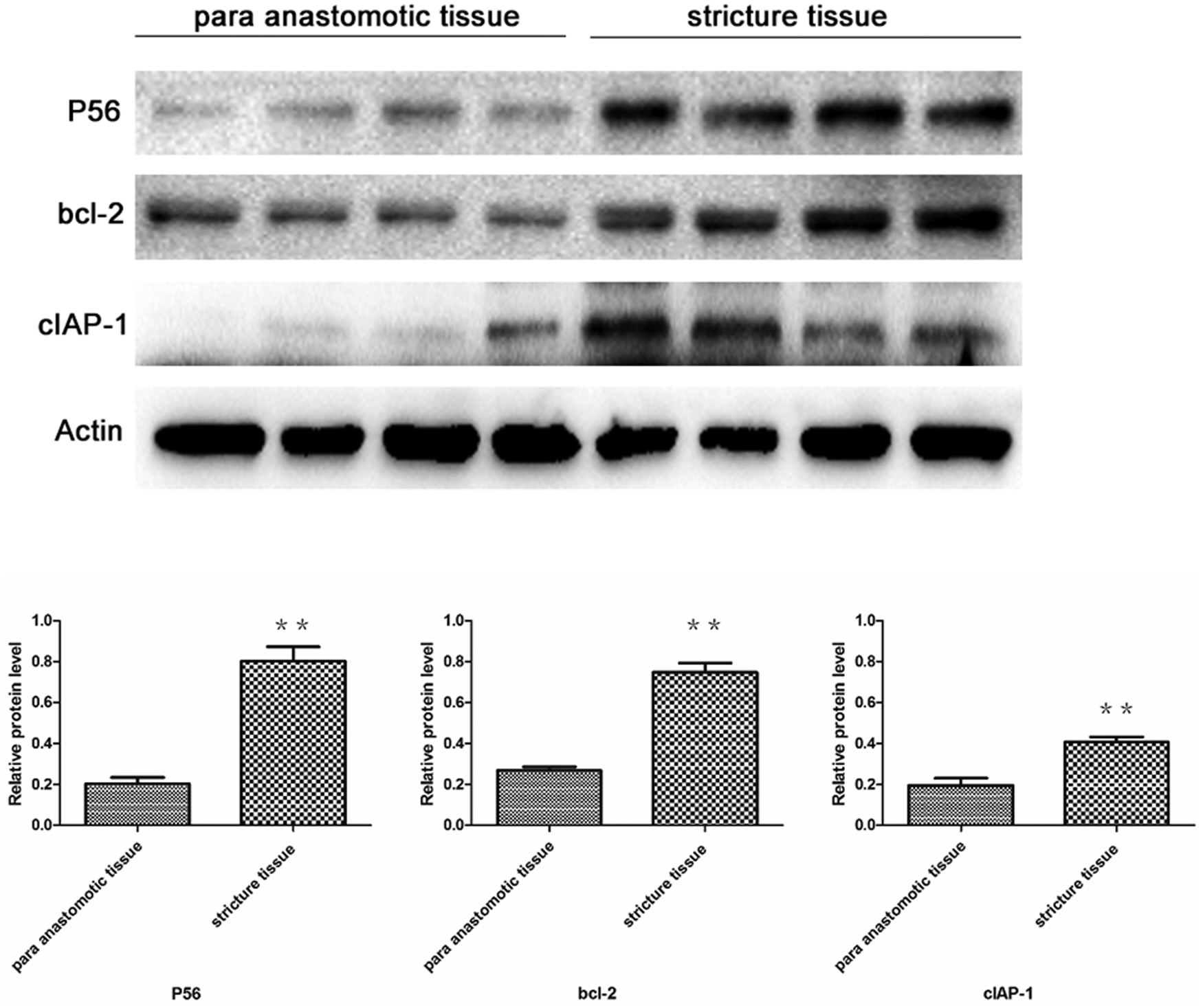
Western blotting of P56, bcl-2, and cIAP-1 protein expression in the para anastomotic tissue and stricture tissue.
Discussion
Anastomotic stricture is a main complication of the esophageal reconstruction. The patients may have severe malnutrition or weight loss due to the dysphagia. 10 Through the literature review, there are two dominating factors which may result in anastomotic stricture. One is the tension, the higher tension located on the anastomosis, the higher risk of developing the stricture; the other is the inflammation, the more inflammatory factors stimulate the anastomosis, the higher risk of developing the stricture.11–13 At the present time, the primary prevention and treatment of esophageal strictures involve the advanced anastomotic technique, bougie dilatation, balloon dilatation, steroid injection, mitomycin C incision, and stent. 14 But none of them can completely heal up the stricture, and up to 40% of esophageal strictures may recur after a few months, leading to repeated treatment. 15 The anastomotic stricture is the process of pathological hyperplasia. When getting some therapeutic targets in the gene and molecular level, we may make a big progress in the treatment of anastomotic stricture.
The NF-κB is a transcriptional factor involved in the cell proliferation, autoimmune diseases, and cancer development. 7 Meanwhile, NF-κB has an essential role in the process of fibrous hyperplasia. The overexpression of NF-κB was detected in the keloid fibroblasts, and abnormal activation of NF-κB was associated with numerous inflammatory diseases, infection, and cancer. 16 Through the literature, we found that the NF-κB signal pathway has the relationship with the fibroblast hyperplasia.17,18 The overexpression of NF-κB could activate the anti-apoptotic genes which can resist cell death, eventually leading to hyperplasia of anastomotic fibers and strictures. 19 Thus, we plan to evaluate the roles of NF-κB signal pathway in the anastomotic stricture formation after the operation of esophageal carcinoma.
It is also known that NF-κB induces the activation of various anti-apoptotic genes. In this study, we detected the protein and mRNA of bcl-2 and cIAP-1 to verify the activation of the pathway. The gene of bcl-2 plays a key role in inhibiting multiple tissue and cell apoptosis, and the main mechanisms include the following: (1) antagonizing apoptotic gene bax, (2) inhibiting apoptotic cytochrome C released into cytoplasm from mitochondria, (3) preventing cytochrome C from activating the gene of capsize in the cytoplasm, and (4) oxidation resistance and maintenance of intracellular calcium homeostasis. 20 And we also learned that bcl-2 was overexpressed in many cancers and activated by NF-κB. Thus, the bcl-2 may be an ideal target to prevent and cure the esophageal cancer and anastomotic stricture after operation. The cIAP-1 gene also plays an important role in inhibiting the apoptosis and can be regulated by NF-κB. The main mechanisms of cIAP-1 inhibiting the apoptosis include the following: (1) IAP can inhibit the activation of capsize 3, 7, 9 directly and (2) IAP can promote the degradation of capsize by special protease. 21 Imoto et al. 22 reported that cIAP-1 was overexpressed in the cells of esophageal squamous carcinoma. Messadi et al. 9 reported that cIAP-1 had the higher level of expression in the fibroblast during the development of keloid.9,22 In this study, it has the same role as the bcl-2. So, the effects of blocking NF-κB activation may not be limited to the fibroblast.
In this study, compared with the para anastomotic tissue, the levels of P56, bcl-2, and cIAP-1 in the anastomotic stricture were significantly raised. Meanwhile, the levels of P56, bcl-2, and cIAP-1 in the normal tissue of anastomotic stricture patients were higher than the no stricture patients. These results suggested that the development of stricture had the relationship with NF-κB level and inhibited expression or activation of NF-κB’s downstream genes, and proteins may be beneficial to reduce the rate of postoperative anastomotic stricture. The inflammation can activate the NF-κB signal pathway and fasten the process of the anastomotic hyperplasia. Therefore, we should control the inflammation around the anastomosis as possible as we can, such as improving the anastomotic technique to reduce the anastomotic leak, regular acid suppression therapy after operation, and so on. All of these methods will reduce the inflammatory reaction around the anastomosis, and it has been reported that these methods can prevent the anastomotic stricture effectively, but the reason is not clarified. Although this study suggests a relative robust result, there still exists limitation such as the small sample size. Therefore, further study is needed.
Conclusion
Through the result of our study, we can confirm that the anastomotic stricture has the relationship with the NF-κB signal pathway. And the bcl-2 and cIAP-1 which are downstream of NF-κB signal pathway could be a new preventive and therapeutic target for the anastomotic stricture. This research provides fundamental information to develop novel therapies for anastomotic stricture.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This study is supported by the Youth Scientific Foundation of The First Hospital of Jilin University (JDYY72016039).