
Abstract
The purpose of this study was to focus on the underlying relationship between the hyperactivity for the peripheral monocytes and heat stroke by investigating the inflammatory oxidative activity of and the expression of superficial molecules. Peripheral blood samples were collected from 10 healthy adult volunteers. Human blood monocytes were isolated by density gradient centrifugation and sequent adherent culture. The objectives were divided into four groups: 43°C heat stress combined with lipopolysaccharide (LPS) group, 43°C heat stress group, LPS group, and control group. There were 10 cases in each group. An enzyme-linked immunosorbent assay (ELISA) test was used to measure the concentrations of supernatant inflammatory mediators (tumor necrosis factor alpha (TNF-α), interleukin-1β (IL-1β) and interleukin-10 (IL-10)). After loaded by 2,7-Dichlorodi-hydrofluorescein-diacetate (DCFHDA) fluorescent probe, intracellular reactive oxygen species (ROS) levels were determined by a flow cytometry. After fluorescent microspheres incubation, the phagocytosis of monocytes was observed under a fluorescent microscope. Respectively, the flow cytometry and Western blot were used to evaluate the level of triggering receptor expressed on myeloid cells-1 (TREM-1) and Toll-like receptor-4 (TLR-4) on the monocytes. Furthermore, the mRNA expression of TREM-1 and TLR-4 was detected by real-time polymerase chain reaction (RT-PCR). The heat stress combined with LPS stimulation promoted the peripheral monocytes to produce inflammatory mediators (TNF-α, IL-1β, and IL-10) and release ROS. Otherwise, such complex strike significantly suppressed the phagocytic activity of monocytes in peripheral blood. Moreover, the expression of TREM-1, TLR-4 and CD86 was measured by the flow cytometry on peripheral monocytes which were respectively promoted by the union of heat stress and LPS. The results of Western blot and RT-PCR demonstrated the similar kinetics on these superficial molecules (TREM-1, TLR-4, and CD86) stimulated by the combination of heat stress and LPS. The underlying mechanism of the dysfunction for the peripheral monocytes may be related to the abnormal expression of superficial molecules TREM-1, TLR-4, and CD86 on the monocytes induced by heat stress and LPS.
Introduction
Heat stroke is a kind of severe lethal disease which is characterized by high core temperature over 40°C accompanied with central nervous system dysfunction such as convulsions, delirium, and coma. In spite of simultaneous cooling, most of the cases will develop multiple organ dysfunction syndrome (MODS) quickly. 1 Recently, the researchers preferred to consider systemic inflammatory response syndrome (SIRS) as the pathological mechanism of heat stroke instead of mono heat stress. 2 The activation and infiltration of inflammatory cells, the damage of vascular endothelial cells and intestinal epithelial cells, the presentation of lipopolysaccharide (LPS) in peripheral blood, and general hemorrhage and thrombosis were evident in the organism those was suffered to heat stroke. 3 The monocytes were the most potent pro-inflammatory agents known as the over-production of inflammatory mediators, cytokines, and oxidation. In the study of sepsis, monocytes were proved to initiate and amplify the inflammatory cascade reaction. Furthermore, the inhibitor or regulator of the monocytes showed a promising protective effect on sepsis. 4 In reviewing the literature, a strong relationship between heat stroke and SIRS has been reported currently. However, few literatures were found that concentrated on the question of the peripheral monocytes whether or not to play a significant role in SIRS initiated by heat stroke. This study set out with the aim of assessing the dysfunction of the peripheral monocytes about activity and superficial molecules in heat stroke-induced inflammation. The result of present research might identify the role of the peripheral monocyte dysfunction in the pathogenesis of heat stroke and help to explore the potential target of clinical treatment for severe heat stroke.
Methods
Isolation and culture of the monocytes
Acquisition of human peripheral blood samples
Peripheral blood (40 mL) was drawn from 10 healthy male volunteers whose ages were from 20 to 27 years old. Anticoagulation was performed to the blood samples with ethylenediaminetetraacetic acid (EDTA) (0.25%). The blood samples were stored at 4°C condition after harvest and tested within 1 h. The blood samples from each volunteer were assigned to each group for investigation. Each sample was measured for three times, and the mean value was collected. This study was approved by the ethics committee in our hospital, and all the subjects had signed informed consent.
Isolation, culture, and assignment of the monocytes
A density gradient equilibrium centrifugation with Ficoll (Maximum density 1.078 g/mL, GE, America, lot number: 17-1440-03) was used to harvest the monocytes from peripheral blood. Further purification was performed by adherent culture. The Trypan Blue test was used to verify the activity of monocytes was over 99% before stimulation. The primary monocytes were inoculated in 60 mm cell culture cluster in three wells, and the quantity of cells in each well was adjusted to 2 × 107. These cells were cultured at 37°C for 12 h. The cells in heat stress(HS) + LPS group were suffered to a 43°C stimulation for half an hour culture at a cell temperature control box, and then another LPS (500 ng/mL) strike was sequently performed to the objectives for 30 minutes. The cells in HS and LPS group were only applied to equal heat stress or LPS stimulation. The cells in control group were only cultured for half an hour at a cell temperature control box at 37°C.
Simulation of the monocytes under heat stroke in vitro
HS and endotoxemia were considered to be the most important and initial pathophysiological mechanism of the development for heat stroke to MODS. Accordingly, LPS combined with HS were performed to prepare the in vitro cell model of heat stroke in several studies.5,6 All the cells were assigned to HS + LPS group and HS group that were pretreated in the incubator (41111FO; Thermo Fisher Scientific, MA, USA) under 43°C ambient temperature for 30 min. Then a 500 ng/mL the management of the concentration of LPS management was practiced in HS + LPS group sequentially for another 30 min. LPS group or control group was subjected to the same LPS or heat stress stimulation alone. After that, all the monocytes were incubated in the condition of 37°C and 5% CO2 for 12 h. The anchorage-dependent cells were collected, and the account of cells in each well was adjusted to 1 × 106 by phosphate-buffered saline (PBS) buffer solution (GE, CT, USA) for the next test.
Detecting the activity and function of human blood monocytes suffered to heat stress and LPS
Enzyme-linked immunosorbent assay test
An enzyme-linked immunosorbent assay (ELISA) kit (RD, Art. No. D1B00B, DTA00C, and D1000B) was used to measure the concentrations of inflammatory mediators (tumor necrosis factor alpha (TNF-α), interleukin-1β (IL-1β), and interleukin-10 (IL-10)) in the peripheral monocytic supernatant according to the manipulation.
Flow cytometry determination
Flow cytometry (BD, LSRFortessa™, USA) was performed to test the level of reactive oxygen species (ROS) in cultured cells after DCFHDA probe fluorescence (Beyotime Biotechnology, China, Art. No. S0033) staining at excitation wavelength of 488 nm and emission wavelength of 525 nm.
Phagocytosis evaluation
The phagocytosis of human monocytes was observed under fluorescence microscope (Olympus IX71, Japan) and detected by flow cytometry after loading by fluorescent microspheres. The phagocytosis of fluorescent microspheres by human monocytes was detected by flow cytometry. After preparation with the fluorescent microspheres, the monocytes (CD45-APC: BD, USA, Art. No. 560973; CD14-PerCP-Cy5.5: BD, USA, Art. No. 561116; microspheres-fluorescein isothiocyanate (FITC): Thermo Fisher Scientific, USA, Art. No. ZZM-2112B) were labeled with CD45 and CD14, respectively, in four experimental groups. The mononuclear cells were labeled by CD45 and then by CD14, and the monocytes were labeled by FITC. There was no flow antibody marker in blank control group. The counting and average fluorescence intensity of peripheral monocytes were measured by flow cytometry at 488 nm excitation wavelength and 525 nm emission wavelength.
Western blot
Human monocytes were ground repeatedly in Dounce homogenizer for 50 times. Then the supernatant was obtained by centrifugation at 4°C for 10 min at 700g speed. Subsequently, the cells were centrifuged at the speed of 14,000g for 30 min under 4°C. Protein concentration was determined by bicinchoninic acid (BCA) assay. The protein samples were denatured after being prepared and heated in boiling water at 100°C for 5 min. The glue was manufactured on the Bio-Rad shelf. The protein samples at the quality of 20 µg were added into each channel for electrophoresis. The voltage was manipulated at 80 V for concentration gum and 120 V for separation gum. Membrane transfer was performed at 300 mA for about 120 min. Then, the membrane was sealed in 5% sealant for 2 h. The primary antibody was incubated at 4°C cold storage and spent the night in a slow shaker as soon as the accomplishment of washing. Then the second antibody was incubated in a slow shaker at room temperature for 2 h. In the darkroom, the film is put into the exposure plate; 200 µL luminous liquid was added to the exposure plate, and the image Quant LAS 4000 mini instrument was put into the detection equipment to analyze the exposure. The gray value of protein was measured and analyzed by using ImageJ software.
Real-time polymerase chain reaction (RT-PCR)
The RNA was extracted from human blood monocytes by Trizol method. The genome was removed by using DNase of RNase-free. Then the extracted RNA and primer mixtures in the PCR tube were stored at 42°C for 60 min, and then the reverse transcription was performed at 72°C for 15 min. Finally, a single gene product was amplified by Roche 480 software, and only a single product was amplified. The mRNA expression level was exhibited as triggering receptor expressed on myeloid cells-1 (TREM-1)/beta-actin (ACTB) and Toll-like receptor-4 (TLR-4)/ACTB = 2–ΔΔCт (ΔΔT = target gene CT – internal reference CT; ΔΔCт = target gene in tested samples ΔCт – target gene in reference samples ΔCт). The primer sequence of TREM-1, TLR-4 and ACTB were listed in Table 1.
The primer sequence of TREM-1 TLR-4 and ACTB.
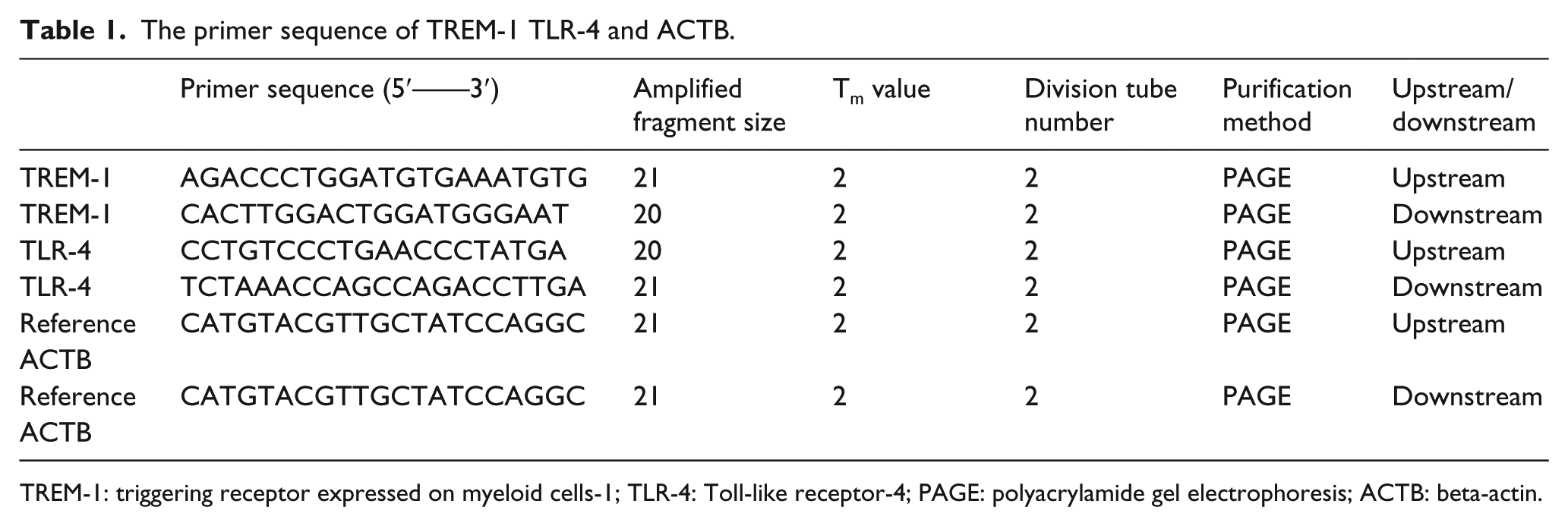
TREM-1: triggering receptor expressed on myeloid cells-1; TLR-4: Toll-like receptor-4; PAGE: polyacrylamide gel electrophoresis; ACTB: beta-actin.
The human blood monocytes were divided into four experimental groups and one blank homotypic control groups, and 1100 r/min was centrifuged for 5 min. Then 300 µL PBS was used to adjust the cell concentration to 1 × 106 and transfers to the dedicated flow tube. Flow antibodies CD45 and CD14 were added to four experimental groups to mark the monocytes (CD45-APC: BD, USA, Art. No. 560973; CD14-PerCP-Cy5.5: BD, USA, Art. No. 561116; CD86-PE: BD, USA, Art. No. 560957; TLR4-PE: BioLegend, USA, Art. No. 312805; TREM-1-PE: BD, USA, Art. No. 565555). The monocytes were labeled by CD45, and then the human blood monocytes were labeled by CD14. The target surface molecules were labeled by phycoerythrin (PE). There was no flow antibody marker in blank control group. The cell counts and percentages were detected by 488 nm excitation wavelength and 525 nm emission wavelength by flow cytometry.
Statistical method
Statistical data were analyzed by software SPSS 20.0, and all the data were represented by Mean ± SD (standard deviation). Multiple data were analyzed by single-factor descriptive analysis of variance (ANOVA) and homogeneity of variance. If the variance is uniform, the method is tested by least significant difference (LSD) method. If not, the method is tested by Dunnett’s T3 method. The difference of P < 0.05 between two groups was statistically significant.
Results
The kinetics of TNF-α, IL-1β and IL-10 in peripheral monocyte supernatant suffered to the stress
The experimental results for the concentration of TNF-α, IL-1β, and IL-10 were demonstrated in Figure 1. The stimulation of heat stress plus LPS obviously increased the production of TNF-α, IL-1β, and IL-10 in peripheral monocytic supernatant compared to other groups (P < 0.05). Moreover, the value of TNF-α/IL-10 was the peak level for HS + LPS group in comparison with other groups (P < 0.05) (Figure 1).

The concentration of TNF-α, IL-1β, IL-10, and ROS in monocytic supernatant under stress—a: 43°C + LPS group, 43°C for 30 min, then 500 ng/mL concentration LPS for half an hour under 37°C; b: 43°C group, 43°C for 30 min; c: 37°C + LPS group, 500 ng/mL concentration LPS for 30 min under 37°C; d: 37°C group, cultured under 37°C. (*) compared with 43°C + LPS group, P < 0.05; (**) compared with 37°C group, P < 0.05; and (***) compared with 37°C + LPS group, P < 0.05.
The intracellular ROS levels in monocytes after irritation
Intracellular ROS levels were detected in flow cytometry after loaded by DCFHDA fluorescent probe. These results illustrated that the combination stimulation of heat stress and LPS promoted the value of ROS in the peripheral monocytes (P < 0.05; Figure 1).
The phagocytic activity of monocytes after stimulation
The monocytes that pretreat with fluorescent microspheres were evaluated under a fluorescent microscope. Flow cytometry was used to detect the number of fluorescent microspheres which were swallowed by monocytes. The results discovered that the fluorescence intensity of microspheres in the monocytes assigned to 37°C + LPS group was significantly greater than the other groups (P < 0.05). Whether or not the involvement of LPS, the phagocytosis of monocytes under 43°C stimulation was lower than the cells under 37°C (P < 0.05; Figure 2).

The percentage of the monocytes with fluorescent microspheres by flow cytometry—a: 43°C + LPS group, 43°C for 30 min, then 500 ng/mL concentration LPS for half an hour under 37°C; b: 43°C group, 43°C for 30 min; c: 37°C + LPS group, 500 ng/mL concentration LPS for 30 min under 37°C; d: 37°C group, cultured under 37°C. (*) compared with 43°C + LPS group, P < 0.05; (**) compared with 37°C group, P < 0.05; and (***) compared with 37°C + LPS group, P < 0.05.
The dynamics of CD86, TREM-1, and TLR-4 on the monocytes under strike
The results of CD86, TREM-1, and TLR-4 on human monocytes with flow cytometry illustrated that 43°C combined with LPS boosted the expression of CD86, TREM-1, and TLR-4 on monocytes rather than those only induced by heat or LPS stress (P < 0.05; Figure 3).

The expression of superficial molecules CD86, TREM-1, and TLR-4 in monocytes with a compound of heat stress and LPS. a: 43°C + LPS group, 43°C for 30 min, then 500 ng/mL concentration LPS for half an hour under 37°C; b: 43°C group, 43°C for 30 min; c: 37°C + LPS group, 500 ng/mL concentration LPS for 30 min under 37°C; d: 37°C group, cultured under 37°C. (*) compared with 43°C + LPS group, P < 0.05; (**) compared with 37°C group, P < 0.05; and (***) compared with 37°C + LPS group, P < 0.05.
The abnormal expression of TREM-1 and TLR-4 on monocytes induced by stress by Western blot
The results showed that the protein expression of TREM-1 and TLR-4 on monocytes in 43°C + LPS group was significantly increased compared to other groups (P < 0.05). Furthermore, the values of TREM-1 and TLR-4 in 43°C group and 37°C + LPS group were more than 37°C group (P < 0.05; Figure 4).

The protein and mRNA expression levels of TREM-1 and TLR-4 in human blood monocytes—a: 43°C + LPS group: 43°C for 30 min, then 500 ng/mL concentration LPS for half an hour under 37°C; b: 43°C group: 43°C for 30 min; c: 37°C + LPS group: 500 ng/mL concentration LPS for 30 min under 37°C; d: 37°C group: cultured under 37°C. (*) compared with 43°C + LPS group, P < 0.05; (**) compared with 37°C group, P < 0.05; and (***) compared with 37°C + LPS group, P < 0.05.
The dysfunctional mRNA expression of TREM-1 and TLR-4 on monocytes by RT-PCR
The results showed that the mRNA expression level of TREM-1 and TLR-4 on the peripheral monocytes of 43°C + LPS group was significantly increased in comparison with other groups (P < 0.05). Moreover, the expression of 43°C group, 37°C + LPS group and 37°C group were presented a gradual decrease in order of precedence (Figure 4).
Discussion
Heat stroke is a common disease induced by outdoor exercise in summer. The underlying relationship between heat stroke and dysfunctional monocytes may be a promising target for heat stroke prevention and treatment. Myeloid cell triggering receptors and Toll-like receptors play significant roles in activating and amplifying inflammatory reaction.5,6 In sepsis patients, the raise of the TREM-1 on neutrophils and monocytes had a positive correlation with the concentration ascending of the inflammatory mediators in peripheral blood. 7 TLR ligand induced the release of IL-1β from human monocyte–derived dendritic cells. 8 Therefore, it was generally assumed that the hyperactivity of monocytes induced by heat stroke was associated with the abnormal expression of superficial molecules.
As the classical pro-inflammatory cytokines, TNF-α and IL-1β were the primary markers of the monocyte inflammatory activity.5,9 Our results showed that values of TNF-α, IL-1β, IL-10, and TNF-α/IL-10 were increased, which is the objective proof for extreme pro-inflammatory activity of the monocytes under heat stress combined with LPS stimulation. ROS was documented to be a classical marker associated with oxidative stress in the tissue. 10 The results illustrated that heat stroke led to the boost of oxidative activity in monocytes. Some studies also indicated that Hsp70 may decrease endotoxin-induced production of ROS and TNF-α in human phagocytes. 11
The clusters of protein located on the surface of monocytes were considered to be the significant modules for initiating immune reaction and inflammatory response. Several studies concluded CD86, TREM1-1, and TLR-4 were co-stimulatory molecules on antigen-presenting cells and played critical roles in the immune system. The results of blocking or inhibiting these molecules showed a protective effect on ameliorating the production of pro-inflammatory cytokines. 12 We suggested that upregulated expression of CD86, TREM-1, and TLR-4 on peripheral monocytes might be the vital molecular mechanism of SIRS or MODS in severe heat stroke.
Footnotes
Acknowledgements
JL and YC contributed equally to this study and share first authorship.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by the National Natural Science Foundation of China (Grant Nos 81503460 and 81671896), Science and Technology Developing Project of Dongguan City (No. 2014108101012), and Guangdong Province Science and Technology Plan Project (No. 2017A020215001).