
Abstract
Background
Current multiple sclerosis management primarily targets symptom alleviation and immune modulation, with limited success in halting progression or achieving sustained remission. Consequently, the development of novel therapeutic strategies targeting the underlying mechanisms of multiple sclerosis (MS) remains a critical area of research.
Objectives
This study investigated the putative neuroprotective properties of Nigella sativa oil (NSO) in a cuprizone-induced demyelination model in adult male Wistar rats.
Methods
Twenty-four adult male Wistar rats were divided into four groups: Group A (Control) received normal mash feed; Group B received 0.2% cuprizone diet; Group C received 5 ml/kg NSO, while Group D received 0.2% cuprizone diet and 5 ml/kg NSO. After 35 days, rats were tested for memory and behaviour (Y-maze, Morris water maze, open-field test). Rats were euthanized, brains were excised then examined for myelin integrity, oligodendrocyte loss, and microglial activation using immunohistochemistry (antibodies: myelin basic protein, oligodendrocyte transcription factor, ionized calcium-binding adaptor molecule 1).
Results
Cuprizone exposure resulted in impaired memory function, reduced exploratory behaviour, and increased anxiety-like behaviours. Treatment with NSO mitigated these behavioural deficits. Additionally, NSO treatment reduced microglial activation and preserved myelin integrity.
Conclusion
Nigella sativa oil ameliorated behavioural alterations, neuroinflammation and demyelination in cuprizone model of MS, suggesting that NSO may have therapeutic potential for MS.
Introduction
Multiple sclerosis (MS) is a chronic autoimmune disorder of the central nervous system (CNS), characterized by inflammation, demyelination, gliosis and neuronal loss. 1 The clinical presentation of MS encompasses a diverse array of neurological manifestations. These include visual dysfunction, paresthesias, focal motor deficits, bladder and bowel dysfunction and cognitive decline. 2 In 2019, MS affected an estimated 1.8 million people worldwide, corresponding to an age-standardized prevalence of 21.3 per 100,000, contributing substantially to reduced quality of life and socio-economic burden. 3 Disease-modifying therapies (DMTs) constitute the cornerstone of MS management. Despite therapeutic progress, MS remains incurable. 4 Current strategies aim to suppress CNS inflammation, delay progression and reduce relapses, yet the expanding range of DMTs for relapsing MS complicates treatment selection. 5
The cuprizone (CPZ) model is widely used in MS research to study the pathogenesis, pathophysiology and behavioral effects of demyelination. 6 In this toxin-induced model, rodents fed 0.2% CPZ for ∼5 weeks develop acute demyelination in distinct white and grey matter regions. 7 Withdrawal of CPZ and return to a normal diet promote spontaneous remyelination within weeks. 8 Although applied for over half a century, the mechanism of CPZ-induced demyelination remains unclear.9,10 While the corpus callosum has been most studied, demyelination is also evident in regions such as the hippocampus and cortex.11,12
Medicinal plants have gained attention for their potential in neurological disorders, as single species often exhibit multiple pharmacological properties. 13 Nigella sativa (black seed/black cumin) has attracted growing scientific and pharmaceutical interest for its neuroprotective potential. 14 Its active compound, thymoquinone (TQ), exhibits antioxidant, anti-inflammatory 15 and neuroprotective properties, demonstrated across various models. 16 Given that current MS treatments are largely symptomatic, Nigella sativa oil (NSO) presents a promising alternative due to its broad pharmacological profile targeting CNS neuroinflammation.
Despite evidence supporting the antioxidant and anti-inflammatory effects of Nigella sativa,14,16 its molecular impact on corticohippocampal regions under demyelinating conditions is still underexplored. The present study addresses this gap by combining behavioral testing with immunohistochemical profiling of myelin basic protein (MBP) and oligodendrocyte transcription factor (OLIG2) in the hippocampus and prefrontal cortex, thereby offering new insights into the therapeutic potential of NSO in MS-like pathology. Hence, this study investigates the potential neuroprotective properties of NSO in a CPZ-induced demyelination model in adult male Wistar rats. The research aims to evaluating the efficacy of NSO in preserving myelin integrity and attenuating neuroinflammatory processes associated with demyelination.
Materials and methods
Twenty-four male albino Wistar rats, aged five weeks and weighing between 100 and 120 g were utilized for this study. The study was conducted within the Animal holding facility of the Faculty of Basic Medical Sciences, University of Ilorin, Ilorin, Nigeria. The rats were fed a standard rodent chow diet and provided with ad libitum access to water. Cuprizone (99%) was purchased from Sigma-Aldrich® (Germany) and prepared at a dose of 200 mg CPZ in 100 g of rat feed (0.2% w/w CPZ diet). 10 Freshly cold-pressed, unrefined virgin NSO containing 0.95% TQ was purchased from Hermani International, KEPZ, Karachi, Pakistan, and administered orally at a dose of 5 ml/kg. 17
The rats were randomly assigned to four groups (n = 6 per group):
Neurobehavioral assessment
At day 35, the rats were evaluated for learning and memory using Morris water maze (MWM) and Y-maze tests 18 also, anxiety and explorative drive were evaluated using the open-field apparatus. 19 The tests were videotaped and later graded by trained blind observers.
Assessment of short memory index
The Y-maze test evaluates spatial working memory. It involves a Y-shaped maze with three wooden arms, labelled A, B and C. Spatial working memory is assessed through spontaneous alternation, where consecutive entries into all arms occur. This method involves interaction with brain regions such as the hippocampus and prefrontal cortex. The Y-maze test was conducted following the procedure reported in Ijomone et al. 20
Long-term memory index
The MWM was used to assess spatial memory and learning in rats. The technique was conducted in accordance with the detailed description provided by Vorhees and Williams. 18
Exploratory drive and anxiogenic behaviour
The open-field test is used to evaluate locomotion, anxiety and stereotyped behaviours in rodents. The open-field test was conducted following the procedure outlined by Gould et al. 19
Collection and preservation of brain samples
Rats were anaesthetized with ketamine (20 mg/kg, intra-peritoneal) while adhering to Institutional Animal Care and Use Committee (IACUC) guidelines. The rats were observed for loss of motor function and then subjected to transcardial perfusion.
Tissue processing
Immunohistochemistry was carried out as previously described in Ijomone et al. 20 In this study, immunohistochemical localization of MBP, Olig2 and IBA-1 (ionized calcium-binding adaptor molecule 1) were carried out.
Image analysis of immunostained slides
Immunostained slides were examined using Olympus (Tokyo, Japan) light microscope and photographed using the AmScope 5.0 Mega pixel USB Still Photo and Live Video Microscope Imager Digital Camera 5MP, manufactured by iSCOPE corp., USA.). Non-overlapping images of the hippocampus and frontal cortex were captured using the 40× objective of the microscope. Image Analysis and Processing for Java (Image J), a public domain software sponsored by the National Institute of Health (USA), was used to analyze and quantify photomicrographs. Quantification of glial cell density (Olig2 and IBA-1 positive cells) and MBP were performed using ImageJ software with automated thresholding and particle analysis.
Statistical analysis
Quantitative data from the behavioural assays were analysed using one-way analysis of variance to assess significant differences among the means of the four groups, followed by Tukey's post hoc test as a confirmatory test. Statistical analysis was done using GraphPad Prism Version 8 (GraphPad Inc, San Diego, USA) statistical software. Normally distributed variables were summarized as mean ± SEM. A p-value < 0.05 was considered statistically significant.
Results
Effect of NSO on CPZ-induced alterations in corticohippocampal-dependent short-term memory
The short-term spatial working memory of rats was evaluated using the Y-maze test on day 35 (Figure 1). The percentage of correct alternations was measured across all groups (n = 6 per group) and expressed as mean ± SEM.
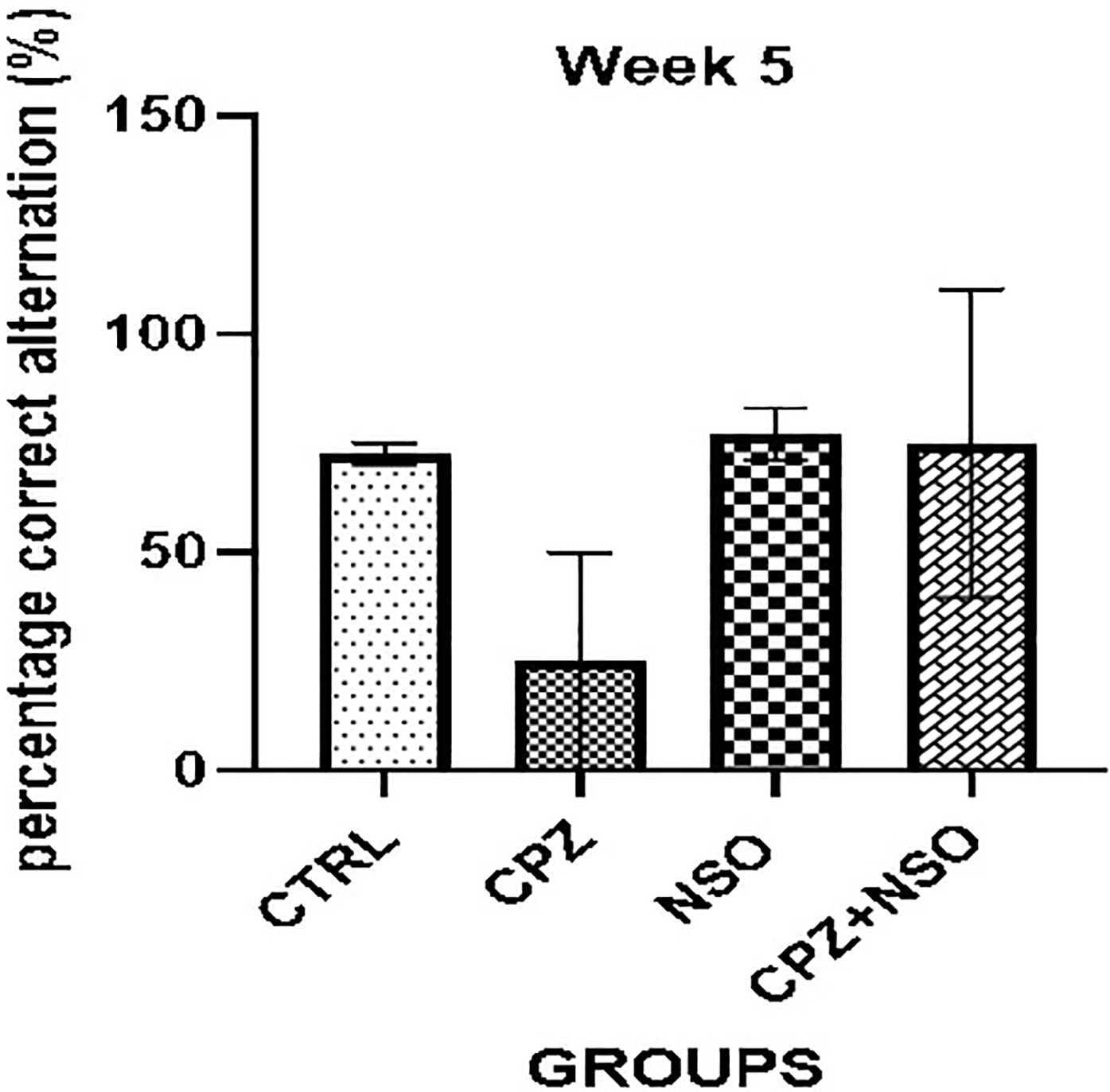
Spatial working memory performance assessed on day 35 using the Y-maze test. Percentage of correct alternations is shown for four experimental groups: control (CTRL), cuprizone-treated (CPZ), Nigella sativa oil-treated (NSO), and cuprizone + Nigella sativa oil (CPZ + NSO). Male Wistar rats (five weeks old, n = 6 per group) were used. Data are expressed as mean ± SEM. No statistically significant differences were observed (p > 0.05). Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey's post hoc test.
The Control (CTRL) group exhibited 72 ± 3% correct alternation. In contrast, the CPZ group showed a marked reduction to 30 ± 10%, indicating impaired working memory. Nigella sativa oil alone led to 75 ± 5%, while the CPZ + NSO group recorded 70 ± 15%. Although CPZ treatment reduced alternation performance, both NSO and CPZ + NSO improved performance towards CTRL levels. However, these differences did not reach statistical significance (p > 0.05 for all comparisons).
Corticohippocampal-dependent spatial learning and memory in CPZ-treated rats following NSO administration
The escape latency period in the MWM test was used to assess long-term spatial learning and memory in all groups at day 35 (Figure 2). Data are expressed as mean ± SEM (n = 6 per group).

Spatial learning performance assessed on day 35 using the Morris water maze test. Escape latency is shown for four experimental groups: control (CTRL), cuprizone-treated (CPZ), Nigella sativa oil-treated (NSO), and cuprizone + Nigella sativa oil (CPZ + NSO). Male Wistar rats (five weeks old, n = 6 per group) were used. Data are expressed as mean ± SEM. No statistically significant differences were observed (p > 0.05). Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey's post hoc test.
The CTRL group recorded an average escape latency of 5.5 ± 1.5 s, while the CPZ-treated group showed an increase to 9.5 ± 5.5 s, indicating impaired long-term memory. However, this difference was not statistically significant (p > 0.05). The NSO group showed 6.5 ± 2.5 s, and the CPZ + NSO group recorded 6.0 ± 0.5 s, both similar to the CTRL group (p > 0.05).
Although CPZ treatment increased escape latency, suggesting impaired learning, co-treatment with NSO appeared to reduce this effect. These differences, however, did not reach statistical significance.
Exploratory behaviour and anxiety-related responses following NSO treatment in CPZ-exposed rats
The activities of rats were examined in the open-field test which is a common measure of exploratory behaviour and general activity in rats. There was a reduction in the number of lines crossed by the CPZ group when compared to the CTRL group but this was not statistically significant (Figure 3(a)). The NSO and CPZ + Nigella sativa groups had an increased number of lines crossed and centre square entry compared to the CPZ group.

Exploratory behaviour and anxiety-related parameters assessed on day 35 (week 5) using the open-field test. Male Wistar rats (five weeks old, n = 6 per group) were divided into four groups: control (CTRL), cuprizone-treated (CPZ), Nigella sativa oil-treated (NSO), and cuprizone + Nigella sativa oil (CPZ + NSO). (a) Number of lines crossed indicates locomotor activity.(b) Frequency of centre square entries and(c) Duration spent in the centre square (seconds) reflect anxiety-related behaviour. (d) Rearing frequency assesses vertical exploratory activity.(e) Stretch-attend frequency reflects risk assessment behaviour. Data are expressed as mean ± SEM. Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey's post hoc test. No statistically significant differences were observed (p > 0.05).
The centre square entry of the CPZ group showed no statistically significant difference when compared to the CTRL group (Figure 3(b)). However, the centre square duration of the CPZ group was lower when compared to the CTRL group (Figure 3(c)). There was a noticeable reduction in rearing frequency in CPZ group (Figure 3(d)). Nigella sativa oil was found to ameliorate this decrease (Figure 3(d)).
Furthermore, the CPZ-treated rats showed an increase in stretch attend frequency (p > 0.05) compared to the CTRL group (Figure 3(e)).
Nigella sativa oil is associated with preserved myelin integrity following CPZ-induced demyelination in corticohippocampal regions
Myelin integrity was assessed by evaluating the expression of MBP and the presence of Olig2-positive cells in brain sections. Rats fed with the CPZ diet exhibited a marked reduction in MBP expression within the hippocampus and prefrontal cortex compared to the other experimental groups. In contrast, the CTRL, NSO, and CPZ + NSO groups demonstrated comparable levels of MBP expression, indicating relative preservation of myelin integrity (Figure 4).
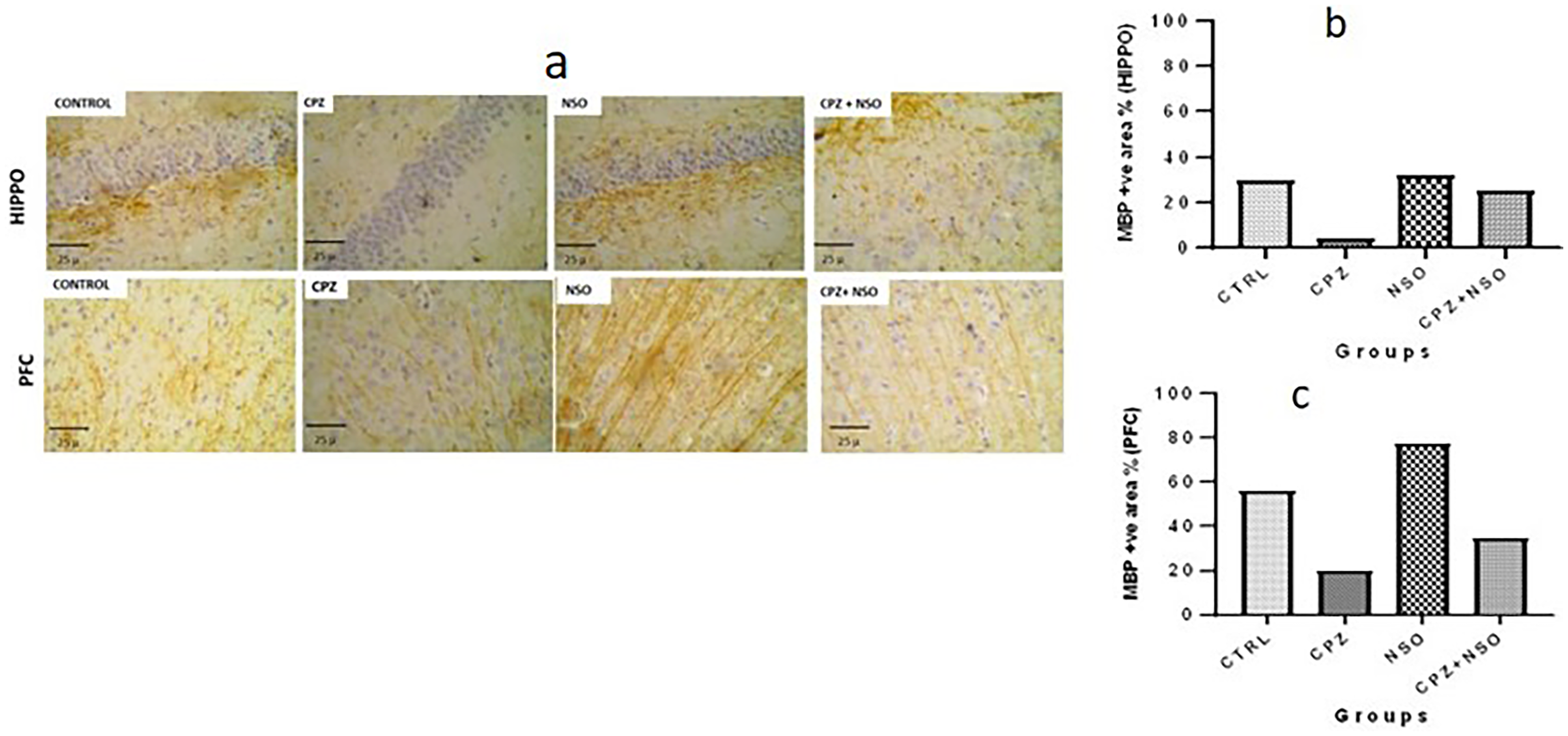
Myelin basic protein (MBP) expression in the hippocampus and prefrontal cortex assessed by immunohistochemistry. Male Wistar rats (five weeks old, n = 6 per group) were assigned to four experimental groups: control (CTRL), cuprizone-treated (CPZ), Nigella sativa oil-treated (NSO), and cuprizone + Nigella sativa oil (CPZ + NSO). (a) Representative micrographs showing MBP-positive staining in the hippocampus (HIPPO) and prefrontal cortex (PFC) across treatment groups. Scale bars = 25 µm. (b) Quantification of MBP-positive area in the hippocampus using ImageJ analysis. (c) Quantification of MBP-positive area in the prefrontal cortex using ImageJ analysis. Data are presented as mean ± SEM. No statistically significant differences were observed (p > 0.05). Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey's post hoc test.
Oligodendrocyte transcription factor 2 expression also revealed a marked reduction in oligodendrocytes in brain sections of CPZ-treated rats. There was a significant decrease in the number of olig2-positive cells in the hippocampal and prefrontal cortical sections of the CPZ group compared to CTRL. On the contrary, there was a significant increase in the number of intact oligodendrocytes in the NSO-treated rats compared with the CTRL, while the number was higher in CPZ + NSO group compared to CPZ only group (Figure 5(a)–(c)).
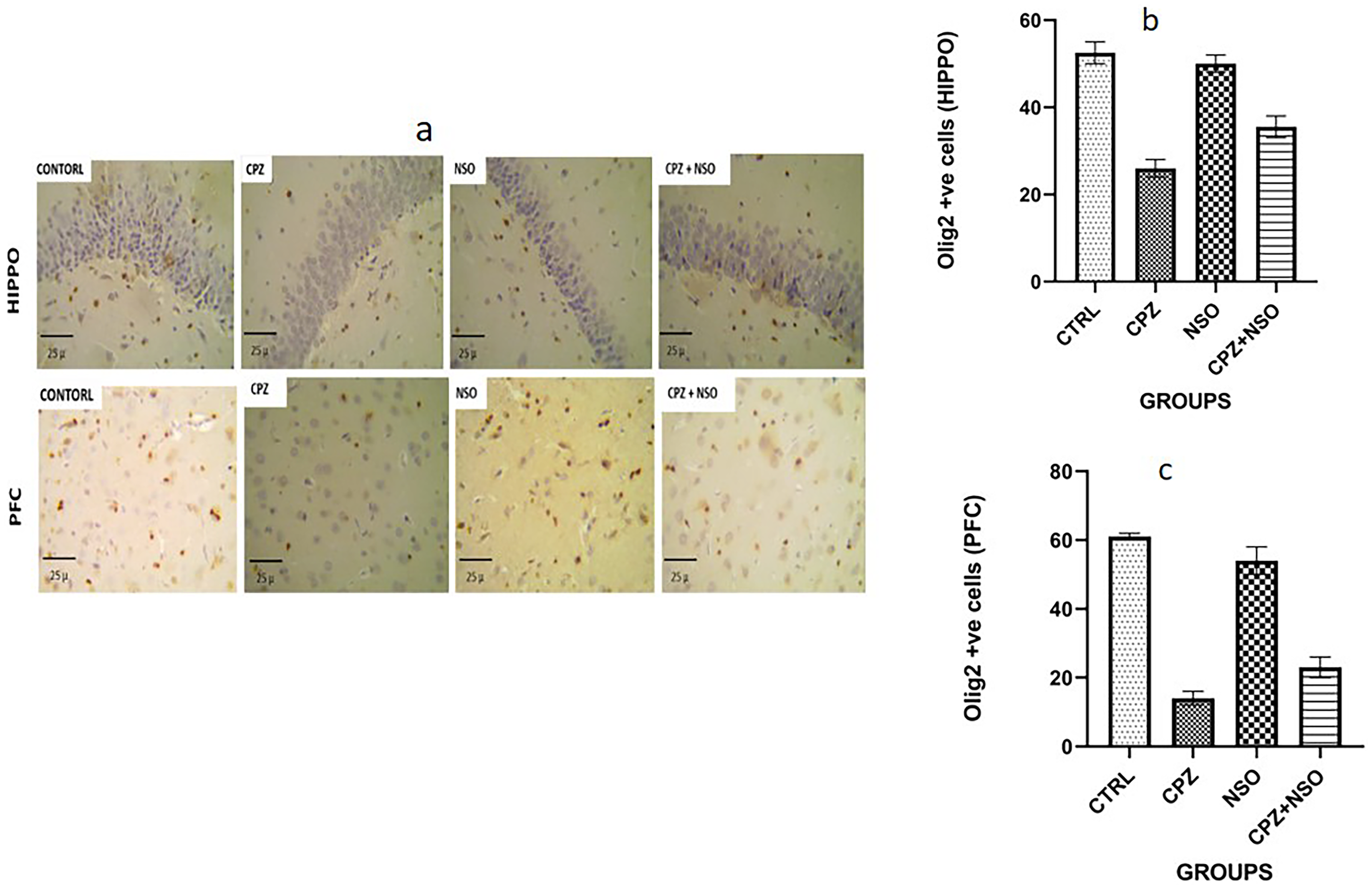
OLIG-2 expression in the hippocampus and prefrontal cortex of rats on day 35. Groups: control (CTRL), cuprizone (CPZ), Nigella sativa oil-treated (NSO), and cuprizone + Nigella sativa oil (CPZ + NSO); n = 6 per group. (a) Representative immunohistochemical micrographs showing OLIG-2-positive cells (brown-stained nuclei) in the hippocampus and prefrontal cortex. Scale bars = 25 µm. (b) Quantification of OLIG-2-positive cells in the hippocampus using ImageJ analysis. (c) Quantification of OLIG-2-positive cells in the prefrontal cortex using ImageJ analysis. Data are expressed as mean ± SEM. Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey's post hoc test. No statistically significant differences were observed (p > 0.05).
Nigella sativa oil modulates CPZ-induced microglial activity in the hippocampus and prefrontal cortex
Ionized calcium-binding adaptor molecule-1 was employed as the immunohistochemical marker for both quiescent and activated microglia in the hippocampus and prefrontal cortex of animals across all experimental groups.
Figure 6 shows the immunohistochemical staining of microglia in the hippocampus, and prefrontal cortex of experimental animals at day 35. In the hippocampus and prefrontal cortex, IBA-1 positive microglia cells were detected in the CTRL, NSO and CPZ + NSO groups; increased IBA-1 staining intensity with more illustrious IBA-1 positivity and intensity was seen in the CPZ group.

(a) Representative immunohistochemical micrographs showing ionized calcium-binding adaptor molecule-1 (IBA-1)-positive cells (brown-stained microglia) in the hippocampus and prefrontal cortex of rats from the control (CTRL), cuprizone (CPZ), Nigella sativa oil (NSO), and CPZ + NSO treatment groups. Scale bars = 25 µm. (b) Quantification of IBA-1-positive cells in the hippocampus at day 35 using ImageJ analysis. Values are expressed as mean ± SEM (n = 6). One-way analysis of variance (ANOVA) followed by Tukey's post hoc test. *p < 0.05, **p < 0.01, ***p < 0.005. (c) Quantification of IBA-1-positive cells in the prefrontal cortex at day 35 using ImageJ analysis. Values are expressed as mean ± SEM (n = 6). One-way ANOVA followed by Tukey's post hoc test. *p < 0.05, **p < 0.01, ***p < 0.005.
Discussion
Prior studies have established a correlation between the progressive deterioration of cognitive abilities and memory function and the demyelination of the hippocampus and prefrontal cortex. 21 According to McNamara et al., 22 modifications in the structure of myelin are sufficient to induce cognitive deficits in animal models of MS.
The results of the Y-maze and MWM experiments suggest that NSO may have mitigated short- and long-term memory impairment in these animals, although these effects were not statistically significant. In a study by Imam et al., 23 NSO reversed the cognitive impairment induced by scopolamine in the Morris Water paradigm by decreasing latency period in both short-term memory and long-term memory tasks, thereby enhancing hippocampal-dependent memory tasks. Thus, this indicates that NSO might prevent memory loss and cognitive dysfunction by acting as a neuroprotective agent.
Furthermore, Bin Sayeed et al. 24 documented the potential modulatory effect of NSO on cognition, attention, and memory. Memory improvement was observed in rats that were subjected to pentylenetetrazole-induced repeated seizures, and memory impairment was reversed in the medial temporal lobium subsequent to scopolamine-induced amnesia in rats. These findings further corroborated the enhanced short-term memory and long-term memory that were observed in the current study. However, at this time, no findings had been reported concerning the impact of NSO on memory and cognitive function preservation in the CPZ rat model of MS. In comparison to prior work, such as the study by Zinvand et al., 25 which examined the combined effects of aerobic training and Nigella sativa extract on neurotrophic factors, tissue damage, and muscle weakness in a CPZ-induced MS model, the present study provides distinct insights. While their findings underscored the systemic benefits of Nigella sativa plant extract in conjunction with exercise, our results demonstrate that NSO alone directly promotes remyelination, enhances oligodendrocyte survival, and reduces microglial activation in the CPZ model. This extends the therapeutic understanding of NSO by shifting the focus from general neurotrophic and functional outcomes to cellular and molecular mechanisms specifically MBP expression, Olig2 regulation, and IBA-1 modulation thereby introducing a novel perspective on its role in MS pathology. Other studies have also explored effect of Nigella sativa plant extract in combination with other formulations in other models26,27 and single brain region. 27
The administration of NSO significantly increased the frequency of line crossings and rearing, which is a gradual indication of enhanced locomotor activities and exploratory drive. This is consistent with our previous findings in permethrin-induced neurotoxicity where we reported that administering NSO to Wistar rats increased their open-field activity and decreased their anxiogenic behaviour. 28 Research has identified several anti-inflammatory and antioxidant constituents in NSO, including TQ, quercetin, kaempferol, and polyphenols. 29 The synergistic action of these compounds facilitate effective remyelination and recover saltatory action potential conduction, thereby aiding in the restoration of motor impairment. 30
The integrity of myelin was assessed in this study through the examination of oligodendrocytes and the expression of MBP. Although MBP is not the only myelin-associated protein, it plays a crucial role in myelin sheath formation and compaction in the CNS and is also involved in signalling and structural scaffolding between oligodendrocytes and the extracellular environment. 31 Demyelination in MS is marked by its degradation. Olig2, a transcription factor that is expressed by cells of the oligodendrocyte lineage, has been linked to compromised myelination when its expression is reduced. 32
According to the hypothesis put forth by Wolf et al., 33 modifications in myelin are responsible for cognitive impairment in rodents and can also be used as a marker for memory loss in aged nonhuman primates and humans. As stated by the author, inflammatory lesions in the white matter are another cause of demyelination in the CPZ mouse model of MS; these lesions are also responsible for the observed clinical deficits. Moreover, in this way, the neurobehavioral deficits observed in the CPZ-treated rodents might have been explicable.
The immunoreactivity of MBP in the prefrontal cortex and hippocampus of rats was generally reduced. Aligned with the findings documented in a comprehensive analysis carried out by Zirngibl et al., 10 the ingestion of CPZ may potentially trigger demyelination via the suppression of myelin protein synthesis, consequently leading to the degradation of the myelin sheath.
Our results indicate that NSO increased the number of MBP immunoreactive fibres in the cortico-hippocampal region, whether administered alone or in combination with CPZ. In an experimental autoimmune encephalomyelitis model of MS, NSO preserved neuronal ultrastructure by mitigating injury to myelin integrity, confirming earlier findings. 34
With the aim of ascertaining whether these deficiencies in MBP were attributable to a regulatory mechanism, we examined the condition of the myelin-producing oligodendrocytes in greater detail. The results of the cell count analysis indicated a decrease in the number of viable oligodendrocytes in CPZ group when compared to the CTRL group.
On the contrary, the NSO group exhibited a higher density of oligodendrocytes and MBP immunoreactive fibres in comparison with the CTRL group. Furthermore, the MBP immunoreactivity and oligodendrocyte density observed in the rats co-administered with CPZ and NSO were only marginally comparable to those observed in the CTRL rats. This result is consistent with the findings of Raimi et al., 35 who investigated the capacity of NSO to regenerate oligodendrocytes in the brain of CPZ-intoxicated quail. In addition, the increased number of oligodendrocytes observed in the NSO-treated group relative to the CPZ-treated group indicated that NSO could hasten remyelination following CPZ-induced injury, which may indicate that NSO possesses neuroprotective properties.
Thymoquinone, the primary bioactive constituent in NSO, is hypothesized to be accountable for the majority of the properties of Nigella sativa. 13 Previous studies have demonstrated that TQ increases the expression of MBP and Olig2, thereby protecting the integrity of myelin. 36
In an experimental model, Ahmad et al. 37 found that TQ treatment increased MBP expression. One plausible mechanism that could account for this upregulation is the facilitation of oligodendrocyte differentiation and myelination. Thymoquinone may increase MBP production by promoting the differentiation and survival of oligodendrocytes. 38
Upregulation of Olig2 expression was observed in response to TQ administration, as shown by Ahmad et al., 37 this finding may indicate that TQ has a protective effect on oligodendrocytes. The precise mechanism by which this modulation occurs is not yet completely understood. By associating motor improvements and memory preservation with reduced demyelination in the CPZ model, this study contributes to existing knowledge by highlighting the potential of NSO in supporting functional recovery under MS-like conditions. This is consistent with evidence suggesting that NSO possesses broad therapeutic properties 29 and may represent a complementary approach alongside current symptom-focused synthetic therapies for MS.
Antioxidant properties have also been identified in TQ, and neurodegenerative processes have been linked to oxidative stress. 37 Thymoquinone potentially bolsters the survival of oligodendrocytes and aids in the preservation of myelin sheaths by impeding oxidative damage and scavenging reactive oxygen species (ROS). The antioxidant potential of NSO was not investigated in this study, although this may represent a mechanism by which it exerts its neuroprotective effects.
García-Revilla et al. 38 have established that heightened microglial expression serves as an indicator of neuroinflammation induced by CPZ.
According to the expression of IBA-1 positivity, microglial activation was observed in the prefrontal cortex and hippocampus of CPZ groups. By secreting vital inflammatory mediators and releasing a variety of substances (e.g., neurotrophic compounds and ROS), microglia contribute to the orchestration of neuroinflammation. Microglia are critical for the subsequent regulation of myelin growth and the associated cognitive function, in addition to preserving myelin integrity through the prevention of its degeneration. 22 Indicators of MS include microglial activation and hyperproliferation, culminating in microgliosis. 33
Positive expression of IBA-1 was reduced in this study following NSO treatment, suggesting that it may possess anti-inflammatory properties. Anti-inflammatory response modulation is a fundamental component of the neuroprotective function of TQ. Microglia and macrophages are integral components in the chronic neuroinflammation that defines MS. According to Pottoo et al. 39 TQ has the potential to inhibit the expression of pro-inflammatory cytokines and chemokines that are involved in the pathogenesis of MS. Thymoquinone potentially inhibits microglial activation, IBA-1 expression and subsequent neuroinflammatory responses by fostering an anti-inflammatory milieu. 39 Microglial activation is frequently accompanied by the secretion of pro-inflammatory cytokines such as tumour necrosis factor-alpha, 40 suggesting that TQ may indirectly contribute to the downregulation of IBA-1 expression.
In addition to polyphenols, quercetin and kaempferol, NSO comprises numerous anti-inflammatory and antioxidant constituents that effectively inhibit inflammation by stimulating anti-inflammatory signalling and alleviate oxidative stress via antioxidant defense mechanism activation.
37
These results provide novel insight into the ability of NSO to act on multiple mechanistic pathways—cognitive, behavioral, inflammatory, and remyelination within a mammalian CPZ model, thereby advancing current understanding of how natural compounds may complement existing MS therapies
Conclusion
This study demonstrates the therapeutic potential of NSO in mitigating CPZ-induced demyelination in adult male Wistar rats, as evidenced by preserved myelin and reduced neuroinflammation. To further elucidate the mechanisms involved, future studies should examine inflammatory cytokines, oxidative stress markers and remyelination-related proteins. Isolating and testing individual bioactive compounds (e.g., TQ) in in vitro oligodendrocyte or microglial models is also recommended to determine their specific contributions to the observed effects. These follow-up investigations will strengthen the scientific basis for NSO as a candidate therapy for demyelinating disorders.
Footnotes
Acknowledgement
The authors acknowledge the collaboration and dedication of all authors involved in this work, whose collective efforts made this research and its publication possible.
Ethics approval
This study was conducted in strict compliance with the guidelines of the IACUC. All experimental procedures were approved by the University Ethical Review Committee (UERC) of the University of Ilorin, Nigeria, under approval number UERC/ASN/2023/2518, and were carried out in accordance with the ARRIVE 2.0 guidelines for the reporting of animal research. Adult male Wistar rats (180–220 g) were housed in plastic cages in a well-ventilated vivarium at room temperature under a 12-h light/dark cycle. Animals were assigned to the respective experimental groups through simple randomisation based on cage grouping and balancing of baseline body weights. All behavioural assessments, histological evaluations, and data analyses were conducted by investigators blinded to group allocations to minimise observer bias. Predefined exclusion criteria included signs of unrelated illness, severe distress or weight loss exceeding 20% of baseline. No animals met these criteria, and all animals initially enrolled were included in the final analysis.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.