
Abstract
Land degradation and desertification are widespread across the world’s drylands. These processes are substantially affected by climatic change, with long-term and severe droughts on the one hand, and high intensity rainstorms and devastating floods on the other hand. Simultaneously, land-use change and mismanagement practices have led to processes of accelerated soil erosion, depletion of soil organic carbon pools, and the degradation of extensive drylands. Forestry has been accepted as an effective means for restoring degraded drylands, and for attaining a range of regulating, provisioning, supporting, and cultural ecosystem services. Specifically, forestry is widely perceived as an effective means for soil erosion control, organic carbon sequestration, microclimate improvement, and climate change mitigation. However, forestry in drylands often proves to generate substantial environmental challenges, resulting in deterioration of ecosystem functions and health. The objective of this essay is to review the challenges and opportunities induced by dryland afforestation and reforestation, and highlight the need to attain climate-smart strategies for establishing and managing these land-uses. Particularly, tree species invasion and allelopathy, which are common in dryland forestry projects, jeopardize species richness and diversity of native vegetation communities. Further, the challenges linked with tree invasiveness necessitate predicting the distribution of potentially invasive species and foreseeing their impacts on the recipient ecosystems under projected climate change scenarios. The effect of allelopathy is significant under limited water availability conditions and is expected to be determined by the expanding drylands and intensifying aridity worldwide. Therefore, judicious selection of tree species should not only focus on ones with high water-use efficiency, low flammability, high pest resistance, and fast growth, but also on low invasiveness and allelopathic capacities. Insights of this essay may be used by land managers, stakeholders, and policy makers involved in environmental development of drylands.
Keywords
Introduction
Desertification is regarded as the degradation of lands in arid, semi-arid, and dry sub-humid areas. The major causes of desertification are changes in climatic (Mirzabaev et al., 2019), atmospheric, and biophysical conditions (Stavi et al., 2010). In recent decades, climate change has become evident, with decreasing precipitation rates and increasing frequency and magnitude of droughts on the one hand, and increasing vigor of rainstorms and severity of floods on the other hand (Bowen, 2015). While these climatic patterns affect vast parts of the world, they are predominant in dryland regions, lowering potential net primary productivity and accelerating land degradation. Over the last decades, the world’s drylands, currently encompassing over 40% of the globe’s terrestrial area (Davies et al., 2016), have faced both aridity aggravation and territorial expansion (Huang et al., 2016). This directly affects the inhabitants of the world’s drylands—encompassing 2.6 billion in 2010 and projected to increase by 40%–50%, to around 4.0 billion, by 2050 (PBL, 2017).
Among the direct “natural” threats to dryland forests, tree desiccation due to increasing aridity and long-term droughts is prominent, causing mass tree mortality over extensive drylands (Choat et al., 2018). An indirect exacerbating effect is the reduced resistance of water-stressed trees to attacks by pests and pathogens (McDowell et al., 2008). This threat is further aggravated by the accelerated territorial expansion and infestation of tree-pests, which in itself is driven by climatic change (Ding et al., 2022). The multifactorial mass mortality of trees is expected to increase forest wildfires’ frequency, intensity (representing the energy that is released during the different stages of fire), and severity (quantifying the above- and below-ground vegetal material that is consumed by fire) (Mcmahon et al., 2020). In turn, carbon dioxide (CO2) greenhouse gas (GHG) emissions increase, and climatic change is further exacerbated (Sommers et al., 2014).
While “natural” factors substantially impact desertification processes, the effect of anthropogenic factors is also prominent (Briassoulis, 2019). Among the anthropogenic effects, land-use change, as well as land misuse and mismanagement, are predominant. Among the land-uses, agricultural malpractices, excess livestock grazing, and clearing of vegetation for fuelwood are most prevalent across the world’s drylands, resulting in degradation of extensive areas. This review study focuses on both the anthropogenic benefits and drawbacks that are related with dryland forestry. Obviously, the impacts of “natural” and anthropogenic factors are usually combined, exacerbating the state of drylands as a whole (Reynolds et al., 2007), and specifically, of dryland forests (de Waroux and Lambin, 2012). Particularly, the combined “natural” and anthropogenic impacts degrade the geo-ecosystem functioning—that is, the inter-relations and feedbacks between the edaphic and vegetal components of the system, which determine its quality and health—across extensive areas. In drylands, among the most prominent aspects of functioning is ecosystem resilience to long-term droughts and climatic change (Stavi et al., 2018). Specifically, since soil-water availability encompasses the major limiting factor for primary productivity in drylands, forestry measures in these regions are required to maximize the accessibility and efficient use of water for the planted trees. Further, the aggravating natural and anthropogenic challenges across the world’s drylands require the implementation of climate-smart forestry measures.
Here we review the impacts of dryland afforestation—that is, tree planting for the creation of new forests in previously non-forested lands—and reforestation—that is, tree planting in previously forested lands (Lund, 2018)—on ecosystem services, which are defined as the ecosystem-derived goods that satisfy human needs, either directly or indirectly (Danley and Widmark, 2016). First, we highlight the regulating services (see the “Climate-smart forestry” section), and then discuss the potential implications for provisioning, supporting, and cultural services (the “Accomplishing additional (non-regulating) ecosystem services” section). Later on, we discuss the main related challenges, focusing on tree species invasion (the “Control and management of invasive tree species” section) and allelopathy, with an emphasis on consequences for native plant communities (the “Managing allelopathic impacts” section). Insights of this essay could shed light on the major challenges and opportunities related with dryland forestry in the face of climate change, assisting in judicious policy-making, and helping to optimize afforestation/reforestation practices.
Climate-smart forestry: Opportunities for enhancing dryland ecosystem services
Afforestation and reforestation can be considered as operative and comparatively low-cost restoration means of degraded non-forest or previously forest lands. Yet, in order to effectively fill this role, these tree-planting schemes should be climate- or (more specifically) climate change-adaptive. Tree species must be carefully selected. Particularly, tree species characterized by high water-use efficiency (WUE) and low water requirements should be preferred wherever possible (Cho et al., 2019). Regardless, the increased frequency and magnitude of wildfires, attributed to global climatic changes, necessitates the selection of relatively low-flammability tree species (Stavi, 2019). Further, the increase in pest infestation that is attributed to climate change, emphasizes the need to select pest-resistant tree species (Woodcock et al., 2018). In terms of carbon (C) sequestration, fast-growing tree species are advantageous over slow-growing trees. By increasing C sequestration in their above- and below-ground biomass, as well as in the soil, fast-growing species potentially reduce atmospheric CO2 concentration (Yosef et al., 2018), thus mitigate climate change (Cunningham et al., 2015). Yet, the common perception of dryland afforestation as an effective tool in mitigating climatic change has been questioned. An approximately decade-old study showed that despite halting desertification (Yakir and Rotenberg, 2010), forestry in (low-cloud high-radiation) dryland regions may increase surface radiation load and decrease albedo effect, thus worsening global warming (Rotenberg and Yakir, 2010). A recent thorough modeling of suitable areas for forestry across the world’s drylands revealed that the forests’ reduced albedo can produce a substantial warming effect, largely counteracting the impact of C assimilation/sequestration in the forests’ trees and soil (Rohatyn et al., 2022). One way or another, through mechanisms such as the REDD + initiative, afforestation and reforestation projects across the world drylands may be eligible for payments for C sequestration, empowering local communities and providing them with an additional source of income (Sadono et al., 2020). However, intra-species (or intra-genus) combinations exhibiting high WUE, low flammability, high pest resistance, and fast growth are not common. For example, in the semi-arid region of central Mongolia, planted Siberian elm (Ulmus pumila L.) were reported to have a smaller leaf size, smaller specific leaf area (SLA), and greater WUE than Populus sibirica Hort. ex Tausch (unknown common name), making them better adaptable to dryland conditions. At the same time, biomass productivity was observed to be ~2.5 times greater for the second species than that for the first one, making it a better choice in terms of C sequestration (Cho et al., 2019). Obviously, substantial differences in these measures are also expected at the species level. For example, in a 5-year experiment in the Great Basin, USA, WUE of limber pine (Pinus flexilis E.James) was found to be generally greater than that of bristlecone (Pinus longaeva D.K.Bailey) (Liu et al., 2021). A schematic illustration of climate-smart tree species selection is presented in Figure 1.

Climate-smart tree species selection in dryland afforestation and reforestation.
An example for a specific strategy in climate-smart afforestation and reforestation projects is the mass application of biochar—a byproduct of C-negative pyrolysis technology for the production of bioenergy—to their soil. Alongside the expected improvement in soil quality and the consequent increase in trees’ primary productivity, this procedure is capable of safely storing tremendous amounts of C, thus substantially lowering atmospheric CO2 concentrations (Stavi, 2012). Hence, this procedure is perceived as a promising strategy in both adaptation to- and mitigation of- climate change (Stavi, 2013). One way or another, while establishing new afforestation and reforestation projects in drylands, special caution should be undertaken to prevent management practices that may accelerate land degradation and desertification (Stavi, 2019).
In addition to C sequestration, afforestation and reforestation may sustain additional regulating ecosystem services that assist in adaptation to climate change impacts (Birch et al., 2010; Li et al., 2020). Specifically, planted forests either on non-forest lands or previous forest lands may be an effective means in increasing conservation of water and soil resources, therefore halting land degradation and desertification. Particularly, the planted trees’ positively impact the formation and stability of soil structure, overall physical quality, water infiltrability, hydraulic conductivity, on-site retaining of surface water, and recharge of underground aquifers (Bargués Tobella et al., 2014). Simultaneously, stemflow and throughfall processes induced by the tree shoots (Krishnan and Venkatesan, 2014), alongside the trees’ litterfall layer, lessen raindrop splash impact underneath the tree canopies, thus lowering the formation of mechanical crusts on the ground surface and lessening water overland flow generation (Leite et al., 2018). Further, both the trees’ canopy and litterfall increase surface roughness, thus decreasing hydrological connectivity (KEFRI, 2014). Consequently, water overland flow is minimized, with subsequently decreasing the runoff’s erosive power, and reducing the loss of mineral and organic materials through erosional processes (Bashan et al., 2012). Further, both the tree-canopy and litterfall layer decelerate wind velocity at the ground surface level, reducing its erosive potential while increasing on-site deposition of aeolian-driven materials (Bird et al., 1992).
Forming sink sites on degraded hillslopes using earthworks engineering has become a common means in afforestation of degraded drylands, by creating embankments or ridges along contours (Stavi and Argaman, 2016). The aim of these systems is to control water runoff and soil erosion at the hillslope scale, and restore the source-sink relations of dis-functioning geo-ecosystems, such as those undergoing inter-rill, rill, and gully erosional processes (Stavi et al., 2015). These systems are often established using dozers and other heavy machineries. Dozer operation is considered to be cost-effective, however, it removes the soil A-horizon from the hillslopes’ ground surface, negating surface roughness, and depleting the soil organic C pool (Stavi and Argaman, 2016; Stavi et al., 2015). The resulted clearing of small-scale geodiversity of the ground surface and eliminating the soil-seed bank, degrade the geo-ecosystem functioning and deteriorate environmental quality and health (Stavi et al., 2019). Additionally, the heavy machinery grinds and compacts the remaining soil on the hillslopes’ ground surface, further degrading its physical quality and lowering hydrological conductivity, while increasing hydrological connectivity at the terrace scale. Although the constructed sink sites increase water availability for the trees that are planted upslope of the terraces, the functioning at the hillslope scale becomes substantially degraded. Studies in the semi-arid northern Negev of Israel revealed that these adverse impacts persevere even a decade after the land-use change from semi-natural lands to the afforestation systems (Stavi and Argaman, 2016; Stavi et al., 2015). At the same time, less intensive systems, such as stone terrace systems—with or without tree planting in the terrace sites—are generally considered to induce a smaller environmental footprint, effectively enabling the restoration of degraded dryland ecosystems (Stavi, 2019). Figure 2 displays a schematic illustration of a bulldozer establishing an earth terrace system (top); the main features of natural and engineered hillslopes (middle); and a 2-year old contour bench terrace’s afforestation system in the semi-arid northern Israeli Negev.
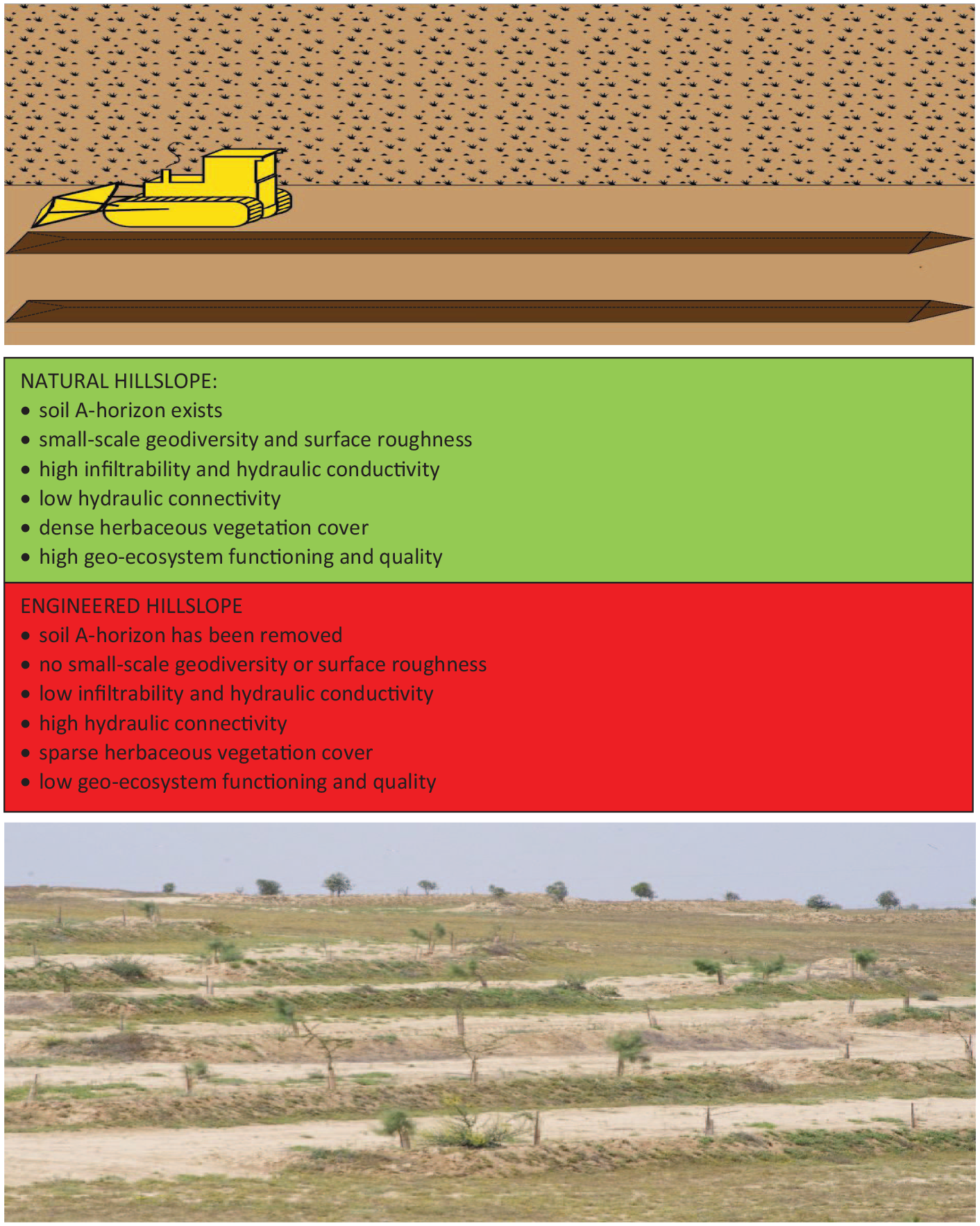
Schematic illustration of a bulldozer establishing an earth terraces system (top); main features of natural and engineered hillslopes (middle); a 2-year old contour bench terrace system in the semi-arid northern Israeli Negev (bottom).
Regardless, dryland forestry may be considered an effective means in phytoremediation. For example, in the Chennai region of south India, saplings of auri (Acacia auriculiformis A.Cunn. ex Benth.), neem (Azadirachta indica A.Juss. 1830), lebbeck (Albizia lebbeck (L.) Benth.), Indian rosewood (Dalbergia sisso Roxb.), and portia tree (Thespesia populnea (L.) Sol. Ex Corrêa) were reported to effectively extract chromium (originated from tannery effluent) from contaminated soils, practically phytoremediating these toxified lands (Manikandan et al., 2016). Further, afforestation or reforestation for alleviation of soil salinity and sodicity—whether derived by natural or anthropogenic causes—is specifically relevant for drylands, which are more susceptible to these processes than non-dryland regions. A wide range of tree and shrubs species can be planted for this purpose (e.g. Yildiz et al., 2017).
A schematic illustration of dryland afforestation and reforestation’s regulating ecosystem services, as well as of their provisioning (e.g. timber production), supporting (e.g. increased biodiversity), and cultural (e.g. recreation sites) ecosystem services (see discussion in the next section), is provided in Figure 3.
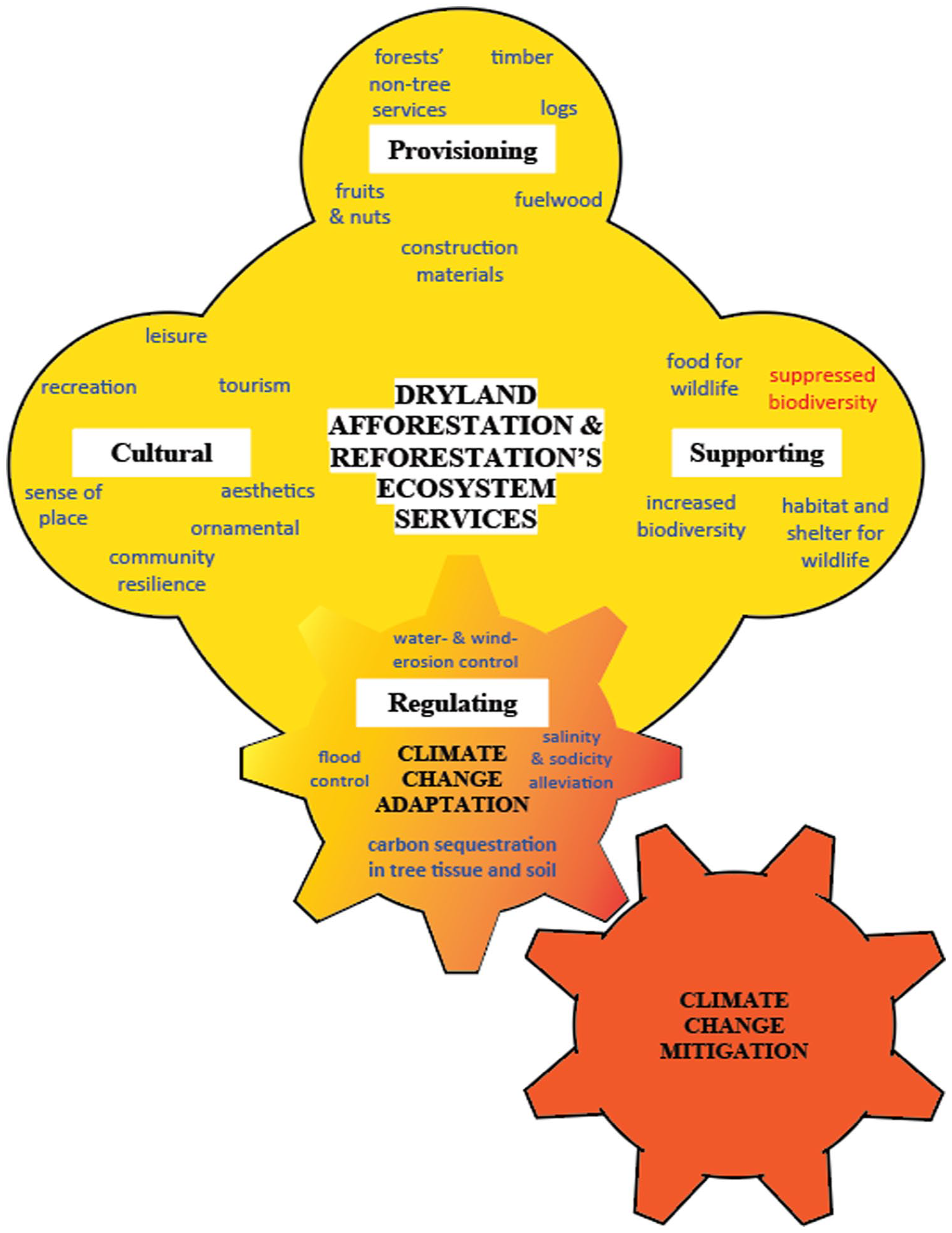
Schematic illustration of ecosystem services in dryland afforestation and reforestation systems, with beneficial effects in blue script and potentially adverse effects in red script. The regulating ecosystem services (within the yellow-orange cogwheel) encompass specific adaptations to climate change. Particularly, carbon sequestration in tree tissue and soil (in bold), reduces atmospheric concertation of the CO2 GHG, thus mitigate climate change.
Ecosystem synergies for accomplishing additional (non-regulating) services
Ecosystem services related with afforestation and reforestation projects are somewhat similar to those of natural forests. Also, to some extent, ecosystem services related with forest systems in drylands are similar to those in non-dryland regions. Yet, a major difference is related to the specific attributes of biomes, ecosystems, and plant communities in dryland regions, as opposed to those in moister regions. Throughout this paper, the discussed specific ecosystem services are (mostly) related to the characteristic conditions of drylands.
In terms of provisioning ecosystem services, the most prevalent ones are timber for logging, wood for construction materials, fuelwood collection, and livestock production (Birch et al., 2010; FAO, 2019a). Another major service encompasses fruits and nuts, which are used by humans as foods and food ingredients. While some of these fruits and nuts can be harvested regularly, other fruits and nuts are usually harvested when the availability of “regular” foods is limited (FAO, 2011, 2019a). A prominent example genus in this terms is the Quercus (oak), of which some of its species produce acorns that can be used as foods or food ingredients (Stavi et al., 2022). For example, in Northern African countries, acorns of the cork oak (Q. suber L.) are considered traditional food, and are known to have a high nutritional value and high anti-oxidative activity (Fennane and Rejdali, 2016). Also, acorns of many oak species are regularly used as feed for livestock (Stavi et al., 2022). Trees’ non-fruit parts may be used as ingredients for either traditional or modern medications (Andersen, 2007). Further, forestry’s non-tree foods include mushrooms, herbs, roots, and tubers (Fennane and Rejdali, 2016). Additional uses of dryland forestry systems are beekeeping for honey production, as well as big and small game (FAO, 2011, 2019a). Obviously, many additional provisioning ecosystem services are species-specific. For instance, cork, which is stripped from cork oak across southern European countries and the Maghreb (Duarte and Bordado, 2015). Another species-specific service is the Arabic gum, which is extracted from the gum arabic (Senegalia Senegal, syn. Acacia Senegal (L.) Britton, syn. Acacia nilotica (L.) P.J.H.Hurter & Mabb., syn. Vachellia nilotica,) and red acacia (Vachellia seyal syn. Acacia seyal (Delile) P.J.H.Hurter) across extensive drylands in Sudan, Chad, and other sub-Saharan countries (FAO, 2011).
In addition to merely being tree plantations, afforestation and reforestation sites can also be designed as multi-purpose agroforestry systems. For example, in alley cropping, tree rows and agricultural crops are interplanted, enabling mixed land-use that can be perceived as both economically viable and environmentally sound. Several types of alley cropping exist across the world’s drylands, of which nitrogen-fixing tree species—which provide nitrogen for agricultural crops—are rather common (Hillbrand et al., 2017). Silvopasture systems encompass planted trees and either a planted inter-tree herbaceous cover, or a natural mix of native herbaceous and shrubby vegetation between tree rows. Both of these systems are common across the world’s drylands, and used for livestock grazing (Pinheiro and Nair, 2018). Agrosilvopastoral systems, a further mixed land-use comprising planted trees, agricultural crops, and natural/planted herbaceous/shrubby vegetation, are also common across the world’s drylands (Soler et al., 2018). Yet, depending on the prevailing biophysical conditions, negative interactions between the trees and the inter-tree crops (e.g. competition for limited resources, or the formation of adverse microclimatic conditions) may decrease crop yields and quality, necessitating the careful planning and designing of such systems (Yang et al., 2021). Yet, when judiciously designed, the planted trees may provide a range of regulating services, such as improved microclimate, soil erosion control, and C sequestration in addition to the provisioning (services) of logs, construction materials, and fuelwood (Lu et al., 2018; Santoro et al., 2020). Additionally, like other forestry means, payments for C sequestration in agroforestry systems through designated funding programs may increase profitability of this land-use (Lu et al., 2018; Minang et al., 2014).
In terms of supporting ecosystem services, afforestation and reforestation can apparently increase plant biodiversity. Specifically, when the target dryland encompasses a degraded ecosystem, and the planted tree species are of native origin, this land-use can restore the land unit and strengthen deteriorated plant communities (Bremer and Farley, 2010). Also, the planted trees and shrubs can provide habitats, shelters, and food for wildlife. In addition, many planted trees and shrubs support pollinator communities (FAO, 2019a). However, dryland afforestation and reforestation sites often lessen native biodiversity (Reisman-Berman et al., 2019). The potentially adverse impacts are thoroughly discussed below, in the sections on “Control and management of invasive tree species” and “Managing allelopathic impacts.”
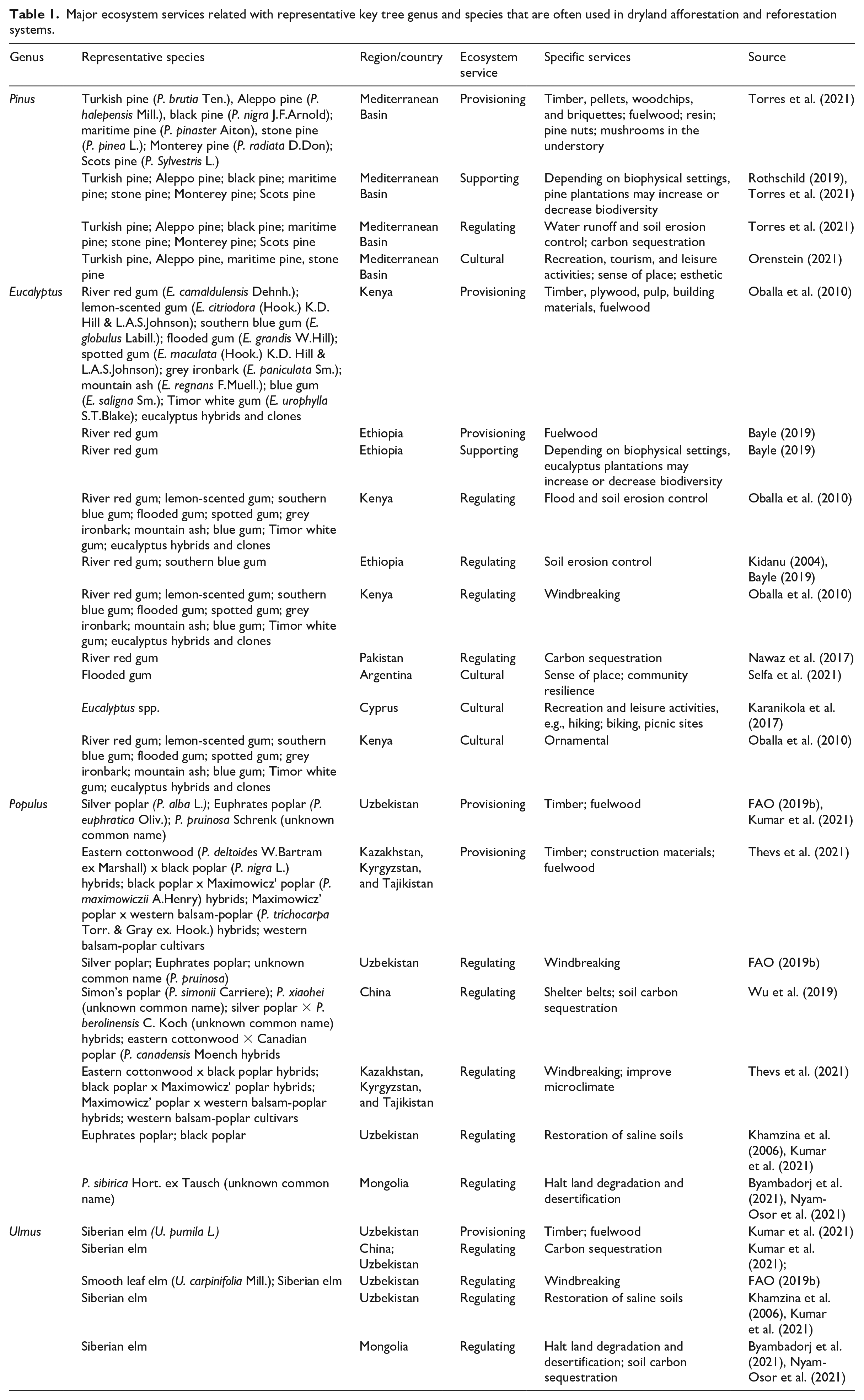
In regards to cultural ecosystem services, some dryland forests fill spiritual and religious functions. For example, the African baobab (Adansonia digitate L.) woodlands fill substantial roles in Senegalese religious traditions. Similarly, the dragon blood tree (Dracaena cinnabari Balf.f.) woodlands bear meaningful spiritual roles in the Yemeni island of Socotra. Yet, these spiritual and religious functions are attributed with natural woodlands more than with planted forests. Regardless, afforestation and reforestation sites are often used for tourism (Birch et al., 2010), leisure, and recreational activities, as well as education and learning, developing sense of place, and strengthening social relationships (Berrahmouni et al., 2015). The leisure experience in dryland forestry sites seems to be particularly important for urban residents, who seek the refuge of the countryside for recreation activities such as hiking and biking, as well as for picnic activities. Further, frequent visits to dryland forestry sites have been reported to increase commitment to environmental protection and conservation (Tal and Billig, 2020). Regulating, provisioning, supporting, and cultural services accomplished by selected key tree genus, which are frequently used for dryland afforestation and reforestation, are detailed in Table 1.
Major ecosystem services related with representative key tree genus and species that are often used in dryland afforestation and reforestation systems.
The alien challenge: Control and management of invasive tree species
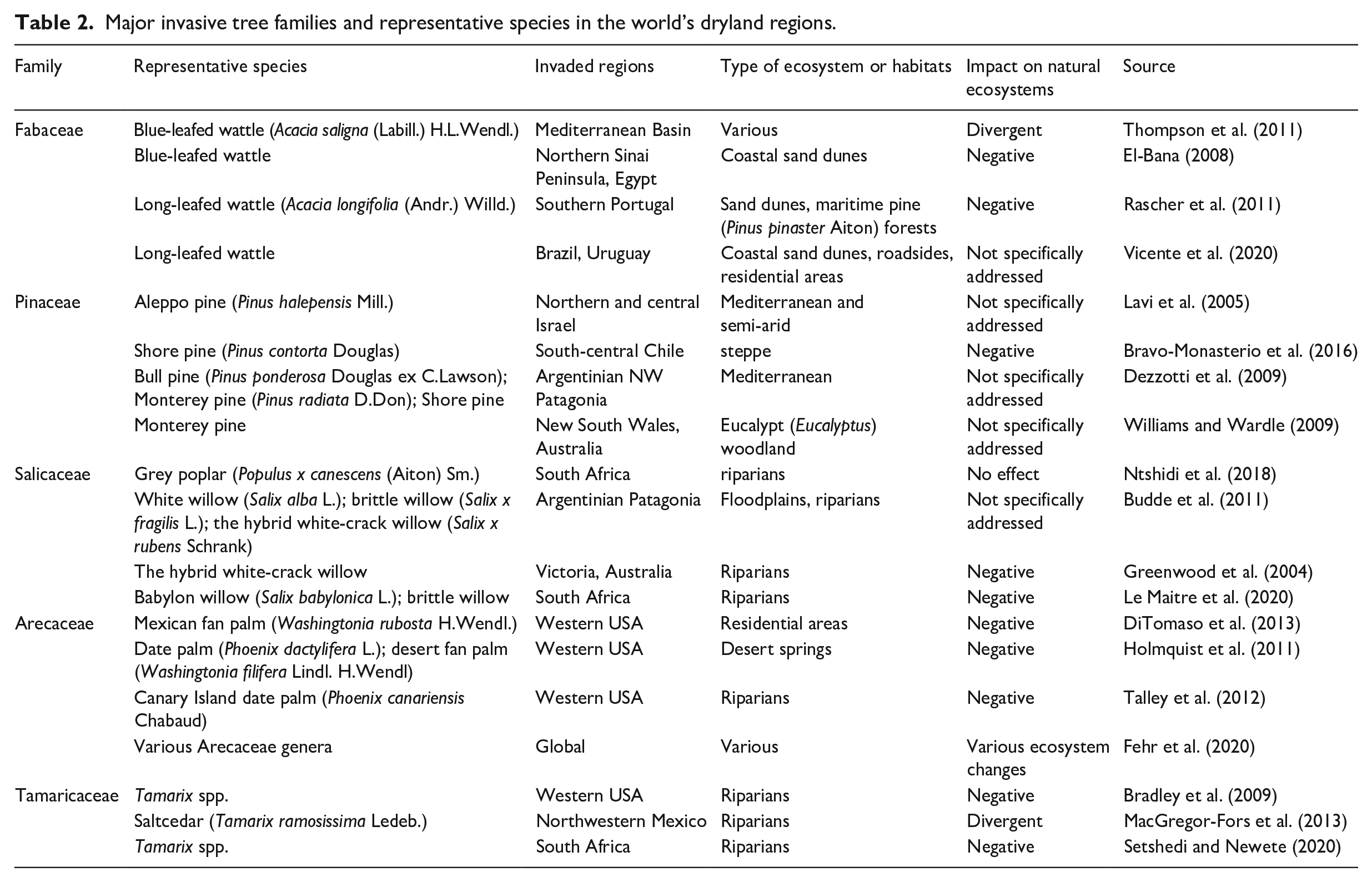
Woody vegetation invasions are rapidly increasing around the world (Lamarque et al., 2011; Richardson and Rejmánek, 2011). The major geographical source regions of invasive tree families are South America and Australia for Fabaceae (Rejmánek, 2014), of which the genus Acacia, and particularly taxa in the subgenus Phyllodineae that is native to Australia are predominant (Richardson and Rejmánek, 2011); North America and Europe for Pinaceae (Rejmánek, 2014), predominated by the genus Pinus (Richardson and Rejmánek, 2011); Eurasia for Salicaceae (Rejmánek, 2014), dominated with genera such as Flacourtia, Populus, and Salix (Richardson and Rejmánek, 2011); Asia for Rosaceae shrubs (Rejmánek, 2014), with the Rubus, Cotoneaster, Rosa, and Pyracantha genera dominating this family (Richardson and Rejmánek, 2011); and tropical and subtropical regions for Arecaceae (Rejmánek, 2014), dominated with Aiphanes, Areca, Archontophoenix, Phoenix, and Washingtonia, among other genera (Richardson and Rejmánek, 2011). Another noteworthy invasive tree family is the Oleaceae (Rejmánek, 2014), predominated by the Ligustrum, Fraxinus, Jasminum, Olea, and Syringa genera (Richardson and Rejmánek, 2011). The most invaded regions are North America, Europe, Southern Africa, Australia, the Pacific Islands (Rejmánek, 2014), New Zealand, the Mediterranean Basin, and the Middle East. The main reasons for invasion are horticulture, forestry, and agroforestry (Richardson and Rejmánek, 2011). Table 2 presents some of the major invasive tree families and representative species across the world’s dryland regions.
Major invasive tree families and representative species in the world’s dryland regions.
Plant invasiveness adversely affect native vegetation composition of recipient ecosystems. Overall, multiple factors determine the success of invasive tree species. For example, in a review analysis of 90 studies with 45 invasive tree species, the tree’s growth rate—and particularly stem elongation, which improves the individual competitiveness for sunlight—was proposed to be predominant (Lamarque et al., 2011). However, this effect seems to rule in non-dryland regions, where sunlight encompasses the major limiting factor. At the same time, in drylands, the fast spread of lateral and vertical roots, aimed at maximizing access to the limited soil-water resource—both over an extended area around the plant core and from relatively deep layers—seems to be the most important factor that determines the success of invasive species. Alongside with soil-water, the root systems of these plants attain limited nutrients, such as soil’s nitrate-nitrogen (NO3-N, see Leakey et al., 1999), further augmenting their competitiveness. The importance of other factors, such as seed mass, mechanism of seed dispersal (Lamarque et al., 2011; Richardson and Rejmánek, 2011), germination rate, length of juvenility, individual’s biomass, and herbivory resistance, seem to be more case-dependent (Lamarque et al., 2011). Regarding seed dispersal, zoochory may accelerate tree invasion through external or internal transport mechanisms. For example, the transport of seeds of the Russian olive (Elaeagnus angustifolia L.) by mammals and possibly also by birds, was reported to accelerate its invasion across the western USA (Campbell and West, 2022). At the same time, seed predation may control plant invasion. For instance, in southern Argentina, the invasive capacity of ponderosa pine (Pinus ponderosa Douglas ex C.Lawson), lodgepole pine (Pinus contorta Douglas), and Douglas fir (Pseudotsuga menziesii (Mirbel) Franco) was found to be moderated through predation of their seeds by rodents and birds (Nuñez et al., 2008).
Climate variables are known to impact the distribution, reproductive success, and survival of non-native plant species (Finch et al., 2021). This impact is generated through three mechanisms: first, by modifying the nature of vectors and pathways; second, by changing the properties of the recipient ecosystem; and third, by alteration of biotic interactions within the recipient community (Robinson et al., 2020). Overall, climatic changes are expected to stimulate vegetation invasions. A widely accepted view is that climatic change boosts the capacity of alien vegetation species to invade new areas, while simultaneously lessening the resistance to invasion of natural ecosystems by hampering their dynamic equilibrium (Thuiller et al., 2007). Particularly, biodiversity loss has been characterized as a slow onset effect of climate change, with resulting adverse ecological, economic, and other impacts (Schäfer et al., 2021).
However, it seems that climatic change impact on plant invasion is complex and scale-dependent. In a systematic review of over 200 modeling case studies on the impact of climate change on plant invasion in terrestrial ecosystems, the effects were found to be highly divergent. Explicitly, in studies conducted at regional or global scales, a decrease in species range size was forecasted more frequently than an increase in it. At the same time, in small-scale or local studies, an increase in species range size was reported to a greater extent than a decrease (Bellard et al., 2018). A modeling study in the western United Stated, which assessed the climatic change impact on distribution of tamarisk (Tamarix spp., which occur mostly in riparian ecosystems) revealed no effect on distribution of this genus (Bradley et al., 2009). At the same time, a study on the spatiotemporal distribution of the invasive long-leaved wattle (Acacia longifolia (Andr.) Willd.) in the coastal regions of southern Brazil and Uruguay, showed a southward migration of the species, suggesting that its density increases with lower temperatures. Also, it was proposed that future climate change will further modify the species distribution’s thermal barrier, pushing it further southward (Vicente et al., 2020).
In addition to the high adaptability of many invasive plant species to limited soil-water availability and drought conditions, making them suitable for drylands, their characteristic positive response to elevated atmospheric CO2 concentrations will likely increase their invasive capacity under the forecasted climatic change scenarios (Finch et al., 2021). Yet, evidence from western and southwestern United States suggest that this CO2 fertilization effect is prominent for herbaceous vegetation more than for trees and shrubs, and specifically, for grass species such as brome grass (Bromus madritensis L.) and downy brome (Bromus tectorum L.), as well as forbs such as purple loosestrife (Lythrum salicaria L.) (Vilà et al., 2007). Overall, the ability to predict the distribution of invasive species and their effects on the recipient ecosystems under projected climate change scenarios is limited, and yet essential for designing operative prevention, control, and restoration strategies (Finch et al., 2021).
In terms of controlling invasive tree and shrub species, the most important aspects that determine the potential risks to natural ecosystems, habitats, or species, were determined as: (1) reproductive potential; (2) dispersal potential; (3) monopolization of resources; (4) ability to outcompete native species; (5) site-dependent settings; (6) facilitation (or inhibition) by climate change; and (7) types and availability of control means. The very same aspects can also be used for the selection of candidate tree and shrub species while planning and establishing afforestation or reforestation projects. Specifically, in terms of control of invasive species, three major types of intervention were proposed, including: (i) prevention; (ii) early detection and rapid eradication; and (iii) management actions such as eliminating, halting of spread, and alleviating the impacts of existing spreading (Vor et al., 2016). Practically, both the early eradication and later elimination practices can include a mixture of physical, chemical, and biological measures (Zahra et al., 2020). One way or another, to a large extent, successful control of tree invasion requires effective policy making, which should support the implementation of climate-smart afforestation and reforestation practices.
The allelopathy challenge: Management and control
Allelochemicals—secondary substances or biochemicals that are not required for metabolism, and cause phytotoxic effects for other organisms—play an important role in plant-plant interactions. These phytochemicals are released from the plant through leaching, root exudation, volatilization, residue decomposition, and other processes (Ferguson et al., 2003). Allelopathic substances involve phenolic compounds, flavonoids, terpenoids, alkaloids, steroids, carbohydrates, and amino acids, as well as mixtures of them (Ferguson et al., 2003). Allelopathic impacts can be either direct or indirect, through mediating agents. Mostly, allelochemicals impede germination, respiration, cell division and elongation, photosynthesis, water and nutrient uptake, and certain enzyme functions (Ferguson et al., 2003; Zahra et al., 2020). At the landscape level, allelopathy in afforestation and reforestation lands may hinder the dispersal and spread of native plant species by fragmenting vegetation connectivity and restricting gene flow (Reisman-Berman et al., 2019). Due to the adverse impacts on root growth of competing plant species and on mycorrhizal fungi, allelopathy is expected to have the most substantial effect where nutrients are scarce and/or where water availability is limited (Conard et al., 1985). Therefore, allelopathy is expected to be strongly determined by the worldwide increasing aridity.
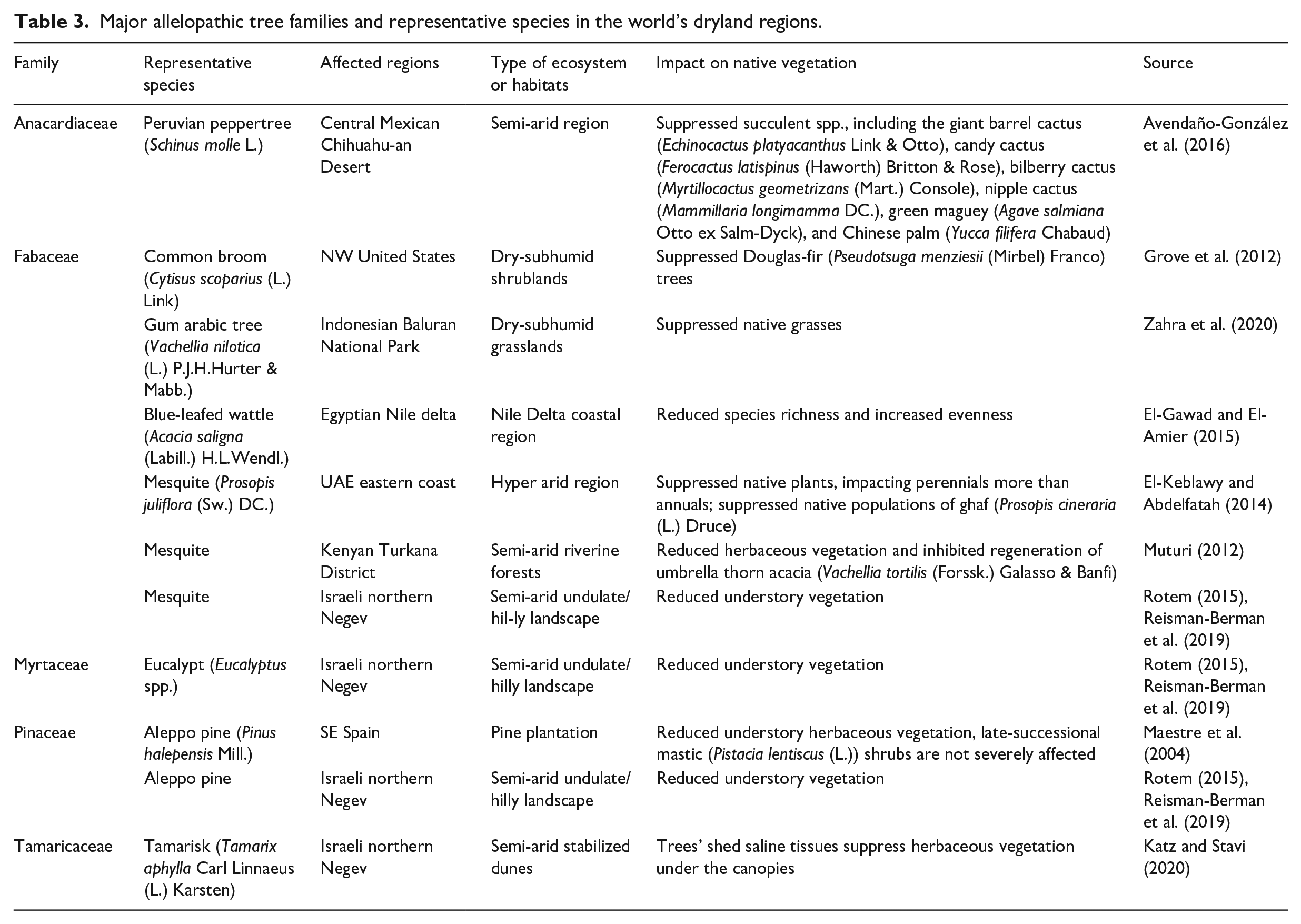
For example, invasion of the nitrogen-fixing common broom (Cytisus scoparius (L.) Link) into the western United States has suppressed the native Douglas fir (Pseudotsuga menziesii (Mirbel) Franco) trees. In addition to the immediate and direct allelopathic impact on growth of this tree species, an indirect effect on the native ectomycorrhizal fungi community is long-term and persistent (Grove et al., 2012). In the Mexican Chihuahuan Desert, the introduction of the Peruvian peppertree (Schinus molle L.) has resulted in substantial suppression of germination and seedling development of native succulent communities. To one extent or another, the giant barrel cactus (Echinocactus platyacanthus Link & Otto), candy cactus (Ferocactus latispinus (Haworth) Britton & Rose), bilberry cactus (Myrtillocactus geometrizans (Mart.) Console), nipple cactus (Mammillaria longimamma DC.), green maguey (Agave salmiana Otto ex Salm-Dyck), and Chinese palm (Yucca filifera Chabaud), were all suppressed by the peppertrees. This allelopathic effect was stronger when induced by male peppertree individuals than by female individuals (Avendaño-González et al., 2016). An example from Africa, the Middle East, and India is the highly invasive gum arabic tree, which contains a wide range of allelochemical tannins in its leaves, bark, and pods, potentially suppressing native vegetation in the recipient ecosystem (Zahra et al., 2020). In the afforestation and savanization lands across the semi-arid northern Negev of Israel, introduced Aleppo pine (Pinus halepensis Mill.), eucalypt (Eucalyptus spp.), and mesquite (Prosopis juliflora (Sw.) DC.) trees were reported to impose allelopathic impacts on native understory vegetation, adversely affecting plant species richness and diversity, and degrading ecosystem functions and services (Reisman-Berman et al., 2019; Rotem, 2015). In the dryland forests of Kenya, allelopathic effect of invasive mesquite trees included reduction in herbaceous vegetation cover and inhibition of umbrella thorn acacia (Vachellia tortilis, syn. Acacia tortilis (Forssk.) Galasso & Banfi) (Muturi, 2012). A specific allelopathic mechanism is attributed to the trees’ saline tissues, which shed and cause soil salinization and sterilization. This effect was reported for a tamarisk (Tamarix aphylla Carl Linnaeus (L.) Karsten) afforestation project in the semi-arid Negev region, aimed at stabilizing mobile sand dunes. The shedding of leaves and litter has formed “salinity islands” that impede herbaceous plant development under the trees’ canopies (Katz and Stavi, 2020). Table 3 presents some of the main allelopathic tree families and representative species across the world drylands.
Major allelopathic tree families and representative species in the world’s dryland regions.
It has been suggested that allelopathy corresponds with invasiveness. Specifically, the “novel weapons hypothesis” proposes that invaders may succeed as they possess novel biochemicals, which favor them over native species (Bais et al., 2003; Callaway et al., 2008). Plant species that coevolved can coexist because they tolerate each other’s toxicity. At the same time, when a species invades a new territory, its toxic compounds may adversely affect native species that have not adapted to its allelopathic mechanisms (Fitter, 2003; Hierro and Callaway, 2003). In the western United States, although not a tree species, the invasive biennial or short-lived Eurasian spotted knapweed species (Centaurea stoebe syn. Centaurea maculosa L.) is an example of native plant displacement. The displacement was attributed to the knapweed roots that exude the phytotoxin catechin. This compound accumulates in soil, hindering germination and growth of native grass species in the invaded range. At the same time, in the knapweed’s original range, germination and growth of other native grass species were not hampered by the spotted knapweed (Bais et al., 2003). Similar mechanisms were also reported for non-dryland regions (Callaway et al., 2008; Hierro and Callaway, 2003).
While allelopathy is perceived as an adverse agent for indigenous ecosystems, it has been suggested that judicious utilization of allelopathic principles may be used as an effective management tool for controlling invasive vegetation. Specifically, it has been shown that native plant species may impose allelopathic impacts on alien species, controlling their spread and infestation. Practically, many native species produce allelochemicals that may function as “novel weapons” against aliens (Chen et al., 2017). This accords with a study in riparian forests of the Spanish continental Mediterranean region, which assessed the allelopathic impact imposed by invasive (tree of heaven (Ailanthus altissima, black locust (Robinia pseudoacacia L.), and Siberian elm) (Mill.) Swingle) and native (white poplar (Populus alba L.)) trees on herbaceous vegetation. Overall, no clear allelopathic differences were recorded between the invasive and native trees (Catalán et al., 2013). Although not in drylands, studies in a subtropical region of southern China showed that in successional forests, abscisic acid in soil caused allelopathic effects on invasive herbaceous plant species (Chen et al., 2017), and that the soil’s concentration of abscisic acid increased with successional stage (Zhao et al., 2011). In a late-successional-stage of forest at the very same region, allelopathy was recorded for the herbaceous vegetation species of goatweed (Ageratum conyzoides L. 1753), black-jack (Bidens pilosa L. 1753), thoroughwort (Eupatorium catarium Veldkamp), morning glory (Ipomoea cairica (L.) Sweet), ivy morning glory (Ipomoea nil (L.) Roth), and bitter vine (Mikania micrantha Kunth) (Liu et al., 2015). Thus, wise selection and planting of certain native tree species may be beneficial in targeting specific exotic species. Compared to conventional management practices such as uprooting, weeding, or herbicide application, the intended use of native trees’ allelopathic mechanisms is more economic and has a lesser potentially adverse environmental footprint (Chen et al., 2017). Yet, it seems that only relatively scant research on this topic has been conducted so far. Moreover, it seems that rather than focusing on management of invasive trees and shrubs, past and current research has predominantly focused on the control of exotic herbaceous vegetation species, and mostly in non-dryland regions. Therefore, it is recommended that decision makers will create supporting conditions, which allow the allocation of resources for assessing the use of allelopathy for controlling alien vegetation in afforestation and reforestation lands.
Conclusions
Land degradation processes are widespread throughout the world’s drylands. These processes, driven by either “natural” or anthropogenic causes, result in the decrease in functioning and quality of extensive lands. Afforestation and reforestation are widely used as a low-cost and viable means of dryland restoration, resulting in improvement in regulating, provisioning, supporting, and cultural ecosystem services. However, these forestry practices may impose substantial challenges to dryland functions and sustainability. Among other risks, accelerated soil erosion and land degradation, species invasion, and allelopathy, are predominant, jeopardizing the functioning, quality, and health of native ecological systems. Therefore, upon establishment of such projects, special caution should be undertaken, ensuring ecological restoration, while negating potentially adverse environmental impacts. Sustainable forestry practices, alongside environmentally-sound selection of tree species, could halt land degradation processes and restore degraded drylands, thus recovering on-site and off-site ecosystem services, and improving quality of life for local communities. Policy makers at all levels (between local to international) should support the extensive use of climate-smart afforestation and reforestation practices across the world drylands.
Footnotes
Acknowledgements
The authors are thankful to two anonymous reviewers, whose comments on the original version allowed the substantial improvement of the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors gratefully acknowledge the Joint Program of the Israel Science Foundation (ISF Grant No. 3257/20) and the National Natural Science Foundation of China (NSFC Grant No. 32061143014) for supporting the study. The Dead Sea and Arava Science Center is supported by the Israel Ministry of Science and Technology. The original sketch of the bulldozer in Figure 2 was generously provided by Yaniv Golan. The authors are grateful to Michelle Finzi for proofreading of the manuscript.