
Abstract
The triumph of the Gaia hypothesis was to spot the extraordinary influence of Life on the Earth. ‘Life’ is the clade including all extant living beings, as distinct from ‘life’ the class of properties common to all living beings. ‘Gaia’ is Life plus its effects on habitability. Life’s influence on the Earth was hard to spot for several reasons: biologists missed it because they focused on life not Life; climatologists missed it because Life is hard to see in the Earth’s energy balance; Earth system scientists opted instead for abiotic or human-centred approaches to the Earth system; Scientists in general were repelled by teleological arguments that Life acts to maintain habitable conditions. Instead, we reason from organisms’ metabolisms outwards, showing how Life’s coupling to its environment has led to profound effects on Earth’s habitability. Recognising Life’s impact on Earth and learning from it could be critical to understanding and successfully navigating the Anthropocene.
Keywords
Introduction
Before being the name of a hypothesis, Gaia was the name of a new entity (Lovelock, 1972), defined as ‘the biosphere and all of those parts of the Earth with which it actively interacts’ (Lovelock and Margulis, 1974a) – where the ‘biosphere’ meant ‘the total ensemble of living organisms’ and ‘the Earth’ refers to the entire planet as an object in the solar system. The Gaia hypothesis proposed that living beings could collectively regulate aspects of their global abiotic environment: the chemical composition of the atmosphere and oceans, and potentially also the climate (Lovelock and Margulis, 1974a, 1974b; Margulis and Lovelock, 1974). The idea of ‘regulation’ often led Lovelock to compare Gaia with an organism, and sometimes with a thermostat, since both are common examples of regulated phenomena.
Gaia fed into the emergence of Earth system science (ESS; Lenton, 2016; Steffen et al., 2020) – where ‘the Earth system’ has been defined in different ways that we outline in the following. It is often taken to refer to the life-supporting entity at the surface of the planet as distinct from much of the inner Earth and its heat source (Lenton, 2016). Subsequently, Lovelock has often said ‘the Earth system’ is just an alternative, scientifically correct name for Gaia – suggesting there is a single global entity that should be a focus of scientific attention. Many Earth system scientists have followed suit, treating Gaia and the Earth system as synonymous. Biologists, however, seem to recognise that Gaia is a distinctly biological proposition, but tend to dismiss it as unworkable in theory (Dawkins, 1982; Doolittle, 1981). Here, we argue that Gaia and the Earth system are distinct and that we are still in the process of discovering Gaia – not least because different scientific disciplines have persistently missed the extraordinary and variable influence of Life on the Earth.
In the following, we examine four key historical reasons why Gaia was hard to spot or hard to appreciate for many scientists. First, we examine the failure of biology to spot the largest living entity – which following recent work we label ‘Life’ (with a capital L) to denote the clade of all living beings stemming from the last universal common ancestor (Dutreuil, 2016, 2018; Mariscal and Doolittle, 2018). Second, we highlight the paradox that Life is hard to spot in the Earth’s energy balance, yet in terms of information and order (negative entropy) it is everywhere and radically changing things. Third, we examine the failure of ESS to study Gaia, highlighting how Gaia is distinct from various definitions of the Earth system (that were in part inspired by the Gaia concept).
As well as seeking to explain why Life was hard to spot and Gaia hard to appreciate, we aim to show a path forwards. Rather than asserting Gaia as an entity, we reason from a different starting point: We review the varying influences of Life on the Earth. When combined with the constraints that non-living conditions impose on Life, this inevitably leads to feedback between Life and its environment, which includes Life’s habitability requirements. A range of resulting ‘complex system’ dynamics are possible, including stabilisation, destabilisation and regulation of habitable conditions.
This leads us to the fourth and scientifically most challenging aspect of Gaia; that a global scale entity was invoked exhibiting goal-directed behaviour – self-regulation of habitable conditions. This seemingly required an engineer or designer where none exists. Such teleological reasoning immediately repelled many scientists (although some began to research feedbacks involving life). For biologists, it was compounded by the absence of a workable evolutionary mechanism to account for it. But as we review, theoretical progress is showing that there is an effective filtering mechanism whereby stabilising feedback regimes tend to persist, increasing the chance of acquiring further stabilising mechanisms. This can explain how Life has become involved in the regulation of habitable conditions – that is, how Gaia has come about.
Such a filtering mechanism necessarily involves making errors – and current human activities, which are globally destabilising, appear to be a case in point. Thus, the occurrence of the Anthropocene does not disprove the existence of Gaia. However, ‘Gaia’ and the ‘Anthropocene’ represent different frameworks for understanding global changes, from contrasting perspectives. It is no coincidence that Paul Crutzen (2002, 2004) – who has known Lovelock since the 1970s – chose to critique Gaia only after introducing the Anthropocene. As such, Gaia offers something distinct from the Anthropocene version of ESS – it provides a different understanding of the world and our place in it and something that we could usefully learn from (Lenton and Latour, 2018).
From life to Life – the failure of biology to see the largest living entity
To understand Gaia, one needs to see that Lovelock was interested in a biological entity that was not studied before by biologists: the biota or ‘Life’ (capital L). ‘Life’ designates a new biological entity, localised in space and time comprising the total ensemble of all the living beings stemming from their last common ancestor, which biologists designate as a ‘clade’ 1 (Doolittle, 2019; Dutreuil, 2016, 2018; Mariscal and Doolittle, 2018). This is distinct from ‘life’ (small l), the class designating the properties common to all living beings. While biologists study organisms, populations, species and so on, until very recently they had never considered the long-lived entity Life as a phenomenon to be studied. While the environment of living things includes other organisms as well as abiotic aspects, Life’s environment is purely abiotic because Life includes all living things.
Gaia is more than just Life in that it includes the effects of Life on habitability. Indeed, the Gaia hypothesis proposed that Life would not have survived without affecting its own habitability conditions. Biologists lack of interest in Life, in its large-scale effects, and in what allows it to persist in the long run, is visible, for instance, in the parallel development of Gaia and ‘niche construction theory’ (Laland et al., 2016). This theory, developed after Lewontin (1983), also emphasised the importance of the effects living beings have on their environment – and the potential adaptive effects which can ensue – but the empirical examples worked upon are mostly local (beaver dams, bird’s nests, etc.; Dutreuil and Pocheville, 2015).
Alongside failing to spot Life and Gaia, evolutionary biologists misread and misunderstood the idea of ‘regulation’ or ‘homeostasis’ in Gaia as something akin to a common good. In their vocabulary, it was seen as something increasing the fitness of all individual living beings – which ought to be explained by altruism – that is, behaviours which are costly (in fitness) for the individuals performing them but beneficial for the collective (Kerr et al., 2004). Yet, the entire absence of the vocabulary and theoretical framework of evolutionary biologists (e.g. fitness, cost-benefit analysis, cheater, altruist) – in favour of cybernetic language in Lovelock’s (1979) prose – along with Margulis having strongly criticised the cost-benefit analysis underlying all the discussions about cooperation in biology – should have alerted biologists to the fact that Lovelock and Margulis were after something else than explaining a putative global altruism.
These two discrepancies – regarding the scale of the living entities considered and the theoretical apparatus to study regulation – are neatly encapsulated in this quote from Doolittle (1981): … the rewards for good (Gaian) behaviors are as remote as penalties for bad behaviour. It is difficult to accept that behaviors whose effects on atmospheric or oceanic composition or global temperature will not be felt for thousands of generations can be selected for, especially when the first beneficiaries of those effects may be organisms which are not themselves responsible for them. (p. 61)
The paradox of invisible Gaia
Life on Earth was hard to spot for a simple reason: it is nearly invisible in the energy balance of the Earth system. Life is powered by only a tiny fraction of the total absorbed solar energy. Yet, in terms of negative entropy and information, Life is everywhere and radically changing things. With that tiny fraction of available energy, Life has profoundly altered atmospheric composition and hence the Earth’s longwave energy fluxes and has significantly changed planetary albedo and hence the Earth’s shortwave energy fluxes, thus exerting considerable leverage on the overall energy balance. This agency must come from the informational quality of Life. Thus, the energetic and entropic views of Life are very different.
Photosynthetic organisms manage to convert on average only ∼0.5 W m–2 or ∼0.3% of the ∼180 W m–2 solar radiation incident on the Earth’s surface into the chemical energy of sugars in gross primary production. This total free energy input of ∼264 TW (Dyke et al., 2011) is the power supply for Life. Around half of it is respired by the photosynthesising organisms, and the other half providing net primary production (food supply) to the rest of Life. This power supply is tiny compared to the radiative energy balance of the planet (which absorbs ∼120,000 and ∼80,000 TW at the surface) and modest compared to the ∼900 TW power supply available from atmospheric circulation alone (Dyke et al., 2011). However, it is considerably larger than the surface power supply from Earth’s internal heat source (Dyke et al., 2011; Rosing et al., 2006).
Life has a somewhat larger hand in altering the Earth’s surface albedo and hence the total ∼160 W m–2 of solar radiation absorbed at the surface. Only a small fraction of the photons absorbed by photosynthetic organisms are successfully converted to chemical energy; hence, the total absorption of sunlight used to fix carbon in photosynthesis is larger at ∼1.8 W m–2 (Dyke et al., 2011). Correspondingly, vegetation generally lowers land surface albedo, particularly in the boreal high latitudes (Betts, 2000), although grasslands can increase albedo relative to bare soil. Phytoplankton also generally lower ocean surface albedo resulting in an extra ∼0.25 W m–2 absorption (Frouin and Iacobellis, 2002), although coccolithophore blooms with their calcareous liths increase surface albedo. These biological effects on surface albedo can clearly be seen in the visible spectrum, including from space (Sagan et al., 1993), and some were very familiar long before the Gaia hypothesis, but their modest magnitude barely hinted at Life’s existence.
Life on land increases evapotranspiration, cooling the surface and affecting cloud cover. This has long been appreciated, but largely viewed as a local or regional effect. The majority of the ∼80 W m–2 lost from the Earth’s surface in evaporating water comes from the ocean.
We now know that Life has increased both atmospheric and cloud albedo through the production of aerosols and cloud condensation nuclei (CCN) that scatter solar radiation, affecting the ∼80 W m–2 of incident solar radiation absorbed by the atmosphere and ∼80 W m–2 reflected by clouds. However, the discovery of these effects largely postdates (and was inspired by) the Gaia hypothesis. When simply viewing the physics of atmosphere and cloud albedo (and even the aerosol chemistry leading to it), underlying biological drivers (where present) are well hidden.
The largest effects of Life on the Earth’s energy balance come via altering atmospheric composition and hence longwave fluxes. The atmosphere absorbs ∼360 W m–2 of the ∼400 W m–2 radiated by Earth’s surface and re-radiates ∼330 W m–2 back to the surface – a phenomenon commonly termed the ‘greenhouse effect’. Life has had a major impact on the ‘greenhouse effect’ by altering atmospheric composition – but this is well hidden when just viewing the energy balance. Its discovery marked the start of the Gaia hypothesis.
The fact that Life is hard to spot in the energy balance of the Earth system – both in terms of the small direct power supply to Life and the largely indirect and thus invisible nature of biological influences on radiative fluxes – may be a key reason why climatologists have often ignored the Gaia hypothesis. There are notable exceptions (McGuffie and Henderson-Sellers, 2014). However, many prefer to see the climate system as a gigantic ‘heat engine’ that just redistributes and degrades the free energy (low-entropy, shortwave radiation) coming from the Sun, using that to do work such as the movement of the atmosphere, oceans and hydrological cycle.
From Gaia to the Earth systems – losing sight of Life
Gaia started off with the recognition of the new biological object – Life – and its effects on atmospheric composition and hence its own conditions of habitability. It studied Life’s expansion and connections with its surroundings, crystallising new scientific problems – such as understanding its maintenance through time. The focus of Gaia on the long-lived entity Life also marks out the difference between Gaia and various conceptions and definitions of ‘the’ Earth system – particularly what we will term the ‘NASA’ and ‘IGBP’ Earth systems. From a historical point of view, Gaia influenced these conceptions of the Earth system, but despite a shared recognition of the interactions between living things and their environment, what constitutes the fascinating core of the Gaia idea – the centrality of Life – was never adopted by them. Here, we contrast the different conceptions of Gaia and the Earth system, touching on their historical context and focusing on the normative framework from which their definitions followed. Table 1 summarises the different way in which their boundaries are delineated.
Different recipes for identifying Gaia and variants of the Earth system.

NASA: National Aeronautics and Space Administration; IGBP: International Geosphere-Biosphere Programme.
Gaia
The recognition and definition of Gaia stemmed from two different sources. The first one, emphasised by Lovelock, was the detection of life on other planets. Thinking about this issue, Lovelock (1965) came to recognise, and then strongly emphasise (Lovelock, 1972), that Life’s imprint on the Earth is visible in the constitution of the atmosphere, maintained in chemical disequilibrium by living beings’ metabolism producing oxygen and methane (and other reduced gases). The second source of reflection for Lovelock stemmed from his daily activities as a consultant both for major scientific institutions of the Earth sciences (such as the National Oceanic and Atmospheric Administration (NOAA) or the National Center for Atmospheric Research (NCAR)) and for chemical and petroleum industries. Armed with his Electron Capture Detector, Lovelock crossed the oceans and atmosphere and found chemical entities produced by living beings, for example, dimethyl sulphide (DMS; Lovelock et al., 1972), and by human activities, for example, chlorofluorocarbons (CFCs; Lovelock et al., 1973).
Thinking about Life viewed from space, on the one hand, and tracing the delicate chemical connections between Life (including humans) and its environment, on the other hand, led to the recognition of a new entity and to the tracing of its boundaries, redefining our conceptions of Life and the environment. If living beings produce aspects of their environment, then their material boundaries no longer stop at their membranes and epidermis, and they expand as far as their influence on the environment can reach. Thus, what was beforehand thought of as ‘abiotic’ such as the atmosphere can now be seen as a living part ‘like the fur of a mink or the shell of a snail’ (Lovelock, 1972).
To trace Gaia’s boundaries thus requires (1) identifying all living beings (bacteria, horses, etc.) on Earth, (2) tracing carefully the chemical and material interactions and connections between these living beings and what is outside their membranes, (3) establishing which of those connections are relevant to habitability (or to Life’s persistence) and (4) recognising Gaia as the entity isolated by the resulting network of connections. Recognising and measuring living beings’ massive influence on their environment, on the one hand, and thinking about the puzzle of Earth’s long-term stability, on the other hand, led to the hypothesis that it is precisely the former which accounts for the latter.
NASA
NASA’s definition of the Earth system was guided by other imperatives during the 1980s. NASA needed a bold and ambitious research programme to prevent the massive cuts in funding promised by the Reagan administration arguing that we have already visited the solar system (Goldstein, 2009). Hence, the space agency turned towards the Earth by developing a billion dollars programme of new satellites (Conway, 2008). The global changes (climate change and the ozone hole) and the recent emergence of plate tectonics gave a new vision of the Earth, more dynamic and interconnected than recognised beforehand. The two massive outcomes of Cold War geophysics – plate tectonics and climatology – along with its programmes and institutions (Doel, 2003; Edwards, 2010; Hamblin, 2013; Turchetti and Roberts, 2014), thus paved the way for the realisation that the Earth system is a real object comprising ‘physical, chemical, biological and human components’ and seen as ‘a related set of interacting processes operating on a wide range of spatial and temporal scales, rather than as a collection of individual components’ (NASA ESSC, 1986). If studied in its entirety, it thus ought to be studied in an interdisciplinary manner, linking biogeochemistry, climatology, oceanography, solid Earth geophysics and so on. NASA’s ESS programme needed the support of both the fluid Earth community and of solid Earth geophysics: thus, it included all the timescales of Earth’s history (NASA ESSC, 1986). In order to organise the scientific work in an interdisciplinary fashion, NASA proposed to structure the programme according to the timescales of the processes studied (rather than according to their material nature – biological, physical, chemical – as traditionally done by disciplines).
To delineate the boundaries of NASA’s Earth system thus requires identifying all processes occurring on Earth and the way they interact. Notice here the main difference with Gaia; one does not start with Life. Consequently, there can be a Mars system just as there can be an Earth system (and a study of Mars’ climatology, past tectonics, etc.) even if there was no life on Mars. Whereas in the absence of life, a ‘Martian Gaia’ would be meaningless. Correspondingly, NASA’s ESS focuses on the presence of liquid water as the sine qua non of habitability – which is really an abiotic definition of the potential for life. This leads to the definition of a ‘habitable zone’ (around a given star) in terms of the range of surface temperatures at which a planet can retain liquid water. Thus, NASA chose an abiotic definition of habitability and lost sight of Life in the Earth system.
IGBP
IGBP was the institution that did the most to put forward the idea of ‘global change’ and to expand it beyond a pure physics of climate to include biogeochemical cycles as well as human aspects (Dutreuil, 2016; Kwa, 2005a, 2005b, 2006, Kwa and Rector, 2010; Uhrqvist, 2014). At the inception of the IGBP, there was internal tension between the proponents of a research programme which would aim at understanding and predicting the future ‘guided by socioeconomic motivation’ and those advocating for IGBP’s research programme to be an ‘intellectual challenge, guided by the scientific need to enhance the understanding of the total earth system and its biota and the response of this system to external inputs and anthropogenic influences’ (Roederer, 1986). This tension disappeared at the end of the 1990s, during the great synthesis of the IGBP (Steffen et al., 2004), when the ‘Earth system’ expression was put to the forefront. Now, it was clear that the imperative was to study the global changes affecting human existence. Schellnhuber proposed a formalisation of the idea of a sustainable development maintaining Earth’s habitability (Schellnhuber, 1999; Schellnhuber and Wenzel, 1998). Crutzen and Stoermer (2000) then introduced the Anthropocene concept, before the elaboration of other key concepts such as ‘tipping elements’ (Lenton et al., 2008) and ‘planetary boundaries’ (Rockström et al., 2009).
Whereas habitability in Gaia meant conditions sufficient for Life to persist, habitability within the IGBP context meant habitability for human beings. To identify IGBP’s Earth system and its boundaries thus requires (1) identifying human conditions of existence and (2) connecting these conditions of existence to all known material and processes affecting it. IGBP’s research programme was guided by the imperative to understand how global changes will affect human conditions of existence in the future. For instance, the study of the past of the Earth system (championed by the PAGES project) is largely restricted to recent Earth history, because it can give us quantitative data useful to informing future projections. Thus, IGBP lost sight of Life by focusing more narrowly on what is required for human life.
Other contexts
The ‘Earth system’ label has spread in other contexts, but none of these have put Life at its centre. The Intergovernmental Panel on Climate Change (IPCC) eventually included the ‘Earth system’ label after IGBP’s important lobby within the World Climate Research Programme (WCRP). Here, ‘Earth system model’ is used to designate models including a dynamic carbon cycle (Dahan, 2010; Heymann and Dahan Dalmedico, 2019). This is far from representing the complexity and richness of what the Earth system is for IGBP, NASA or Gaia. In other fields, also connected to Gaia and Lovelock, ‘Earth system’ gets sporadic re-use either simply to change the label of a discipline or to highlight a slight inflection of what is going on in a discipline. For example, ‘Earth system’ is sometimes used to describe long-term geosphere–biosphere interactions (Lenton et al., 2004). In geochemistry, ‘Earth system’ was used to stress the importance of feedback vis-à-vis a simpler model of circulation of matter (fluxes and reservoirs) first elaborated by Bob Garrels (Berner, 1999; Kump, 1988; Lenton and Watson, 2011; Lovelock, 1986). Solid Earth physics also sometimes uses the ‘Earth system’ label to stress the important dynamic character of the Earth (Condie, 2005; Rollinson, 2007).
Similarities and distinctions
One important novelty, common to all the Earth systems here, is the interdisciplinary study of a global object (crossing biological, chemical and physical entities), from a particular perspective. A ‘perspective’ (Wimsatt, 1994) is what constrains one decomposition of an object into parts or processes and the scientific problems to be solved within this perspective. This perspective differs for each ‘system’ (Table 1): Gaia starts with Life and follows its material connections with the environment and the way in which they affect Earth’s habitability during the whole of Earth’s history; NASA tries to identify all the processes linked together at a given timescale; IGBP strives to disentangle the processes affecting humans thriving in the future. These differences have consequences for how a particular aspect of ‘the’ Earth system is treated: for example, the IPCC conception usually treats humans as if they were outside the Earth system (in that scenarios of human activity act as an external forcing factor to Earth system models, the output of which then drives impacts models of the consequences), whereas IGBP recognised that humans are clearly inside the Earth system.
The different Earth systems carry different notions of habitability. In particular, NASA sees the planetary frame (the Earth system) as ‘habitable’, whereas Gaia sees Life profoundly affecting its own conditions for survival and flourishing, and IGBP focuses on what is habitable for humans and how humans are affecting that.
Cybernetic concepts – which would now be called complex systems concepts – while widely used by Lovelock in his formulation of Gaia, are largely absent from the formulation of ESS by NASA, which only stressed the interactions between components of the Earth system, although it actually showed many feedback loops in the famous ‘Bretherton diagram’ (NASA ESSC, 1986). Subsequent incarnations of ESS have placed more emphasis on feedback, and the IGBP’s Amsterdam Declaration 2001 has gone as far as to suggest that ‘the Earth System behaves as a single self-regulating system’. However, it stopped short of saying what it regulates towards – that is, it stopped short of assigning the system a goal – which would be habitable conditions in the cybernetic Gaia formulation – that is, homeostasis. This is arguably a consequence of not having centred the definition of the system on Life. This imputing of a goal function is one of the central objections to Gaia, which we return to in the section ‘From the assertion of goal functions to a workable theory’.
The varying influences of Life on its material environment
Having highlighted why Life was hard to spot or side-lined from the perspectives of biology, climatology and ESS, we now briefly review the influence of Life on the Earth. This retains the procedural spirit apparent in the heydays of Gaia’s elaboration – captured by Latour (2017): We strive to render Gaia visible by tracking down the influence of Life on its environment.
Atmospheric composition
Lovelock originally noted the profound impact of Life on the balance of gases in Earth’s atmosphere, creating an extraordinary state of thermodynamic disequilibrium (Hitchcock and Lovelock, 1967; Lovelock, 1965; Lovelock and Giffin, 1969).
Oxygen at ∼21% of today’s atmosphere is almost entirely a biological product (of oxygenic photosynthesis by cyanobacteria, algae and plants), which would have a surface concentration of only ∼10−12 atm in the absence of Life (Haqq-Misra et al., 2011). The concentration of methane in this oxygen-rich atmosphere is a factor of ∼1030 greater than expected at equilibrium, as are the concentrations of hydrogen and ammonia (Lovelock, 1975b). This extraordinary order (negative entropy) in atmospheric composition can only be explained by an input of free energy via photosynthetic Life. An estimated ∼0.7 TW is required just to maintain the O2–CH4 coexistence (Simoncini et al., 2013).
Carbon dioxide (CO2) is extraordinarily scarce in the Earth’s atmosphere when compared to Mars and Venus (which have atmospheres dominated by CO2; Lovelock, 1972). Current human activities notwithstanding, Life has turned CO2 from a dominant component of the atmosphere to a trace gas, by locking up organic carbon in sedimentary rocks and by accelerating the weathering of continental silicate rocks and the resultant deposition of carbonate sediments. In the absence of Life, atmospheric CO2 would be up to ∼10- to 100-fold higher producing a radiative forcing of ∼10–20 W m–2 (Schwartzman and Volk, 1989).
Recent work suggests that Life has also lowered atmospheric pressure by transferring nitrogen (the largest constituent of the atmosphere) to the crust, by fixing atmospheric N2 and sequestering some of the resulting organic nitrogen in sedimentary rocks (Johnson and Goldblatt, 2018). This weakens the greenhouse effect by reducing the ‘pressure broadening’ of longwave absorption lines of other gases, an effect which outweighs the reduced Rayleigh scattering of solar radiation by N2 (Goldblatt et al., 2009).
Other biogenic gases generate aerosols and sometimes CCN, principally affecting Earth’s shortwave radiation balance. Biogenic DMS (Lovelock et al., 1972) produced by a range of marine phytoplankton oxidises to form biogenic CCN, an increased density of which increases the albedo of marine stratocumulus clouds (Charlson et al., 1987), generating a radiative forcing of around –1 W m–2 (Boucher et al., 2003). Nearshore macro-algae also produce a range of aerosol and CCN precursors, including methyl iodide (Lovelock, 1975a). Vegetation emits large amounts of isoprene, terpenes and other biogenic volatile organic carbon compounds (BVOCs) that can also be a source of aerosols and CCN (Kesselmeier and Staudt, 1999).
Biogeochemical cycling
Living organisms are made out of a range of elements in stoichiometric proportions that contrast sometimes significantly from their natural abundance at Earth’s surface. Importantly, the inputs of those essential elements in bio-available form to the Earth’s surface are generally meagre compared to the total requirements of Life. Thus, for Life to flourish, it must increase the inputs and/or (re)cycle the essential elements it needs (Volk, 1998). Furthermore, input and/or recycling needs to be greater for elements that are scarcer relative to their biological requirements than those that are less scarce relative to biological requirements. Table 2 summarises some of the effects of Life on the cycling of materials.
Effects of life on material cycling.
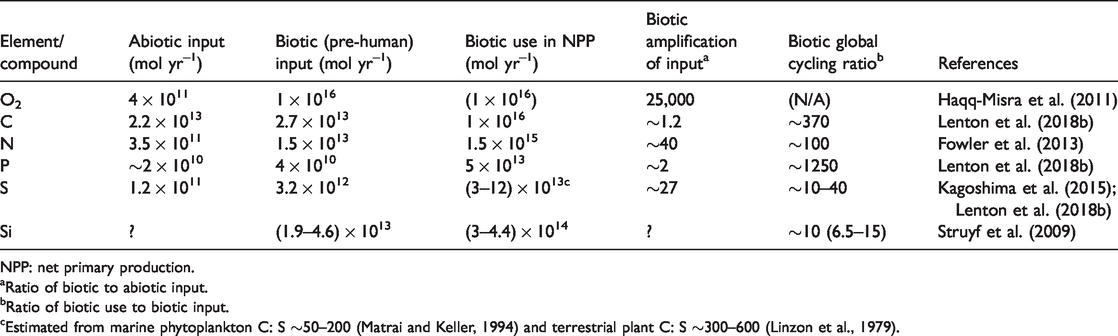
NPP: net primary production.
aRatio of biotic to abiotic input.
bRatio of biotic use to biotic input.
cEstimated from marine phytoplankton C: S ∼50–200 (Matrai and Keller, 1994) and terrestrial plant C: S ∼300–600 (Linzon et al., 1979).
Carbon is the backbone of organic chemistry and photosynthetic and respiratory fluxes of carbon exceed inputs to the surface Earth from the mantle and the tectonic recycling of sedimentary rocks by a global ‘cycling ratio’ of around 400. Carbon input to the surface from tectonic recycling of sediments is also increased somewhat by Life, because the sedimentary organic carbon reservoir is entirely due to Life and its oxidation depends on a biogenic oxygen-rich atmosphere (Lenton et al., 2018b). Furthermore, plankton have shifted the locus of carbonate deposition somewhat from shelf seas to the deep ocean, increasing the tectonic recycling of carbonate carbon (Volk, 1989). Even the supposedly ‘mantle’ input of carbon is partly from what was once subducted organic carbon (Foley and Fischer, 2017).
Nitrogen has an essentially biological cycle in which organisms are responsible for all the key transformations. The input of bio-available nitrogen (
Phosphorus has no significant gaseous form and all ultimately derives from continental chemical weathering. Phosphorus weathering is amplified by Life, by an uncertain factor, through innovations including organic acid production and selective dissolution of P-rich apatite inclusions in rocks. Also, around 20% of the P weathering flux derives from sedimentary rocks whose deposition depended on Life (Lenton et al., 2018b). Phosphorus uptake in net primary production exceeds weathering by a global cycling ratio of ∼1250, higher than nitrogen or carbon, consistent with phosphorus being the ‘ultimate limiting nutrient’.
Sulphur is widely used by Life in varying (and remarkably poorly known) proportions (Linzon et al., 1979; Matrai and Keller, 1994). Life amplifies sulphur input by a factor of ∼30 in that inputs of sulphur are dominated by sediment recycling (Lenton et al., 2018b) rather than mantle input (Kagoshima et al., 2015) and the two major forms of sedimentary sulphur are both attributable to Life: pyrite is a biogenic product and gypsum required biogenic oxygen to produce the sulphate it contains. Sulphur uptake in net primary production further exceeds sulphur input by a global cycling ratio of ∼10–40.
Silicon is required by plants and diatoms (although not all primary producers) with a weathering input that is biologically enhanced by an uncertain amount (Struyf et al., 2009). Plants take up ∼10 times the weathering input and immobilise Si in organic matter, reducing the river input to the ocean, where diatoms take up ∼40 times the river input, indicating marine recycling exceeds the global cycling ratio of ∼10.
A wide range of micro-nutrients are necessary for primary production, or for other key metabolisms, such as nitrogen fixation. The abundance of trace metals, relative to Life’s requirements, varies and has changed with the changing oxidation state of the Earth’s surface environments (Saito et al., 2003; Whitfield, 1981; Williams and Fraústo da Silva, 2006). Life correspondingly exerts varying influence on the cycling of trace metals today, particularly through the production of organic ligands of varying strength (Benner, 2011; Sunda, 2012).
Despite iodine being the heaviest element commonly used by Life, volatile organic iodine compounds, notably methyl iodide, are made in abundance by marine organisms. They play an important role in atmospheric chemistry and aerosol formation and cycle some iodine back from the ocean to the land (Lovelock et al., 1973). There is also considerably terrestrial iodine cycling by Life (Amachi, 2008).
Some elements are not biologically required; hence, there is no incentive for Life to alter their cycling. However, cycling may happen as an accidental consequence of chemical incorporation into biological products, or the chemical consequences of a biological product. A famous example of the latter is the natural nuclear fission reactors from Oklo in Gabon dating to ∼1.7 Ga, which required the prior rise of atmospheric oxygen in the Great Oxidation to concentrate fissionable 235U (Gauthier-Lafaye et al., 1996).
Rock cycling, continent formation and water retention
A large fraction of the diversity of Earth’s minerals are either biologically precipitated or require oxygen (a biological product) in their creation and hence would not exist without Life (Hazen et al., 2008). More fundamentally, it has been proposed that Life has altered the planet’s rock cycle and played a role in continent formation. Life clearly has the energetic potential to do so – its current power supply (264 TW) exceeds the power driving mantle convection (∼12 TW) by a factor >20 and that driving crust cycling (26 TW) by a factor of ∼10 (Dyke et al., 2011). It also exceeds the work done by the atmospheric heat engine in physical weathering (<50 TW) and sediment transport (<13 TW; Dyke et al., 2011).
The production of continental crust as granites requires hydration of the oceanic crust (Campbell and Taylor, 1983). Recently, it has been hypothesised that early Life by permeating the seafloor crust hydrated it and thus started the formation of micro-continents ∼4.0–3.5 Ga (Grosch and Hazen, 2015). Conceivably granite formation was energetically limited prior to the evolution of oxygenic photosynthesis, which greatly increased the power supply to Life (Rosing et al., 2006). The timing of onset of modern-style plate tectonics ∼3.0 Ga is at least consistent with current evidence for the timing of origin of oxygenic photosynthesis. If a link to continent formation is corroborated, this would represent an extraordinary consequence of Life.
Whether Earth would have lost its water in the absence of Life, making the planet uninhabitable, is an open question. In the absence of Life, without biotic enhancement of weathering and possibly without continents, atmospheric CO2 and temperature would be much higher, risking passing the runaway threshold to a ‘moist greenhouse’ atmosphere that supports higher rates of hydrogen loss to space (Popp et al., 2016). Furthermore, by producing an oxygen-rich atmosphere, an ozone layer, and the strong thermal stratification of the stratosphere, Life has created an effective ‘cold trap’ at the tropopause that prevents water reaching the upper atmosphere where it can be split apart and hydrogen lost to space. That said, before causing the rise of atmospheric oxygen, early methanogens likely enhanced hydrogen loss to space by increasing atmospheric methane concentration (Catling et al., 2001).
Distinguishing Earth system and Gaia perspectives
Biologists have had little interest in Life’s influence on the global environment – which is understandable given that climate or ocean composition are not their major object of inquiry. Earth system scientists have increasingly been studying the influence of living beings on the environment – partly thanks to Gaia. But these ‘Earth system’ studies, important as their discoveries have been, have tended to consider biological influences on earthly processes on a par with other abiotic processes. Gaia scientists in contrast have stubbornly tracked down Life’s influence to make it more visible. To render Gaia visible, an important counterfactual – or what-if – question has been asked: how would Earth look had Life had not influenced its environment? (Dietrich and Perron, 2006; Dutreuil, 2014; Lovelock, 1965).
From effects on to regulation of the environment
Lovelock’s (1965) realisation that Life massively alters its environment soon led him to suggest that a new entity is formed by the interactions between Life and its environment – Gaia – and that this ‘influence’ may not be ‘for nothing’: it can account for the very survival and thriving of Life itself.
Blurring the boundaries between life and its environment
Organisms can only survive under particular conditions. As well as liquid water, a source of free energy, and all the materials they need to build their bodies, organisms also require particular physical conditions of temperature, pH, pressure, redox state and so on. More complex organisms typically have more stringent habitability requirements than simpler ones.
Studying the mechanisms by which organisms deal with their material environments has led ecologists and physiologists – and occasionally evolutionary biologists – to realise how fuzzy the boundaries are between organisms and their environment. Hence, the notion of ecosystem, or more recently, that of ‘extended phenotype’, ‘niche construction’ or ‘extended organism’. For example, if one is interested in how a single termite deals with its vital requirement in terms of oxygen, CO2 and temperature, one has to follow all the material links involved in these processes. In so doing, one will cross the termite’s cuticles so many times that the relevant biological entity to be studied – and so, in a sense, the material boundary of this single termite – will be the whole termitary (Turner, 2000).
Early on, Lovelock and Margulis scaled up these two arguments. If organisms have conditions of existence, then aggregating them all together necessary leads to the conclusion that Life also has habitability bounds. Below a given minima and above a given maxima – which may change with evolution – Life would cease to exist. Also, given Life massively influences its environment, then, just as the argument goes for the termite (or for any ecosystem), the boundaries between Life and its environment are blurred and their interactions constitute a complicated entity: Gaia.
Recognising the existence of this entity, Lovelock and Margulis scaled up familiar biological questions and in so doing, introduced new biological puzzles. In particular, Has Life maintained Earth’s habitability?
Habitability
The habitability discussion has focused on climate, after Lovelock and Margulis (1974) proposed that the long-term stability of climate in spite of the Sun’s rising luminosity (the ‘faint young Sun paradox’) may be explained by Life’s influence on the climate. More concrete mechanisms were suggested after the proposal of an abiotic climate regulator (Walker et al., 1981) in the form of a negative feedback involving silicate weathering and the associated uptake of CO2 that can buffer variations in planetary surface temperature and thus maintain liquid water over a wider range of solar luminosity than in the absence of the feedback. This abiotic feedback can broaden the habitable zone bounded by 0°C at which runaway freezing occurs and ∼70°C (rather than the boiling point of 100°C) at which a ‘moist greenhouse’ occurs (the oceans evaporate to create a pressure-cooker atmosphere).
The crucial difference with Life becoming intertwined in feedback loops is that it brings, by definition, habitability constraints (as well as biotic effects) into those feedbacks loops: Life affects environmental variables, just like other abiotic processes; but Life is sensitive to what it does within habitable conditions, whereas abiotic processes are not. This can make negative feedback stronger (a more effective regulator) than in the abiotic case. For example, Life strongly amplifies the silicate weathering feedback on Earth today (Lovelock and Watson, 1982; Lovelock and Whitfield, 1982) and by having organisms such as plants with narrower habitability bounds than 0°C–70°C entwined in the feedback loop, gives rise to narrower stabilising ranges. Furthermore, the biogenic weathering effect with plants may now be so strong (Schwartzman and Volk, 1989) that it could be maintaining Earth in a habitable state for plants, when without them it would by now have become (or could soon become) uninhabitably hot for them (Lenton and von Bloh, 2001). This is supported by recent modelling capturing the interacting effects of water vapour, CO2 and N2, which suggests the ‘habitable zone’ would be disappearing in the absence of Life, which has broadened it considerably (Goldblatt, 2016).
Regulation
Alongside the habitability issue, Gaia scientists looked for examples where the activity of certain living beings (certain parts of Life) regulate an environmental variable and thus produce and maintain their own (and sometimes others’) condition of existence at a regional or global scale. 2
Climate, at various timescales, has again been an important focus of study. For example, the famous Charlson–Lovelock–Andreae–Warren (CLAW) hypothesis highlighted the possibility for climate regulation via biogenic DMS production (Charlson et al., 1987). This drew widespread attention from climatologists, biogeochemists and Earth system scientists. Another important example is the large-scale effects of land plants on the hydrological cycle: for instance, the increase of precipitation enabled by the evapotranspiration of the Amazon rainforest is necessary for the rainforest to persist (Betts, 1999). More generally, evapotranspiration by plants increases the latent heat flux over land up to threefold relative to a desert world (Kleidon et al., 2000). The resulting cooling effect is generally strongest in the tropics, where it outweighs the effect of vegetation lowering surface albedo (Betts, 1999). On longer timescales, it has also been argued that planktonic calcifying organisms, by buffering carbonate-ion concentration in the oceans and thus stabilising the carbon cycle, may have prevented the return of catastrophic ‘snowball Earth’ events during the Phanerozoic (Ridgwell et al., 2003).
The regulation of chemical components has also been scrutinised, such as the long-term regulation of atmospheric oxygen, constrained within bounds not too high to allow sustained vegetation and not too low to enable multicellular animal life using respiration. Atmospheric O2 regulation mechanisms were originally proposed to involve biogenic methane production and fires (Watson et al., 1978). Subsequent suggestions hinge on excess O2 causing fires that suppress land vegetation and make plant photosynthesis less efficient. In one feedback, fires transfer phosphorus from the land to the ocean where less oxygen is produced per unit of phosphorus (Kump, 1988). In another, suppressing vegetation suppresses phosphorus weathering thus limiting oxygen production (Lenton and Watson, 2000b). Both contribute to the long-term stabilisation of atmospheric oxygen (Lenton et al., 2018b).
Proposals for what regulates bio-available nitrogen in the ocean actually predate the Gaia hypothesis (Redfield, 1934, 1958) and are understood to involve a negative feedback on the abundance of nitrogen fixers (that supply available nitrogen): Nitrogen fixation is highly energy-intensive and only advantageous to perform where nitrogen is scarce, but it increases nitrogen levels – limiting its own activity (Lenton and Watson, 2000a). This regulator results in available nitrogen levels in the ocean tracking fluctuations in phosphorus levels – which may be driven by varying input from the continents. Phosphorus levels in the ocean in turn are regulated by the dependence of its biogenic burial (removal) flux on its concentration. For example, an increase in phosphorus levels (and corresponding increase in nitrogen fixation and nitrogen levels) is counteracted by greater productivity driving increased burial of phosphorus (Lenton and Watson, 2000a).
Gaia, Earth system and biology
The empirical description of mechanisms by which Life can maintain habitable conditions or certain living beings can regulate global/regional environmental variables have been the hallmark of Gaia research. This research has fallen through the cracks between ESS and biology: Earth system scientists had little interest in (non-human) living entities maintaining their own conditions of existence; biologists had little interest in empirical material on climate and the global environment and thought the theoretical issues had long been dismissed.
From the assertion of goal functions to a workable theory
Now we turn to the final – and most often quoted – reason why many scientists rejected Gaia: Lovelock’s invocation of goal functions and the apparent purposiveness that comes with that. This was initially compounded by a lack of theoretical mechanisms that could provide an explanation of how ‘global adaptation’ – especially a predominance of regulation – could come about.
The teleology provocation
At the very moment Lovelock discovered Gaia, he attributed it functions, goals and norms. Lovelock was looking for the ‘role’ of the various influences of Life on the environment: What is the function of methanogens within Gaia? Of the Amazonian rainforest?
Lovelock introduced the ideas of feedback, self-regulation, homeostasis and goal-seeking behaviour from cybernetics. Functional talk has not generally hurt engineers or scientists trained in cybernetics: it is very common in systems analysis – where the function of an entity within a larger system studied by the scientist is reduced to the causal effects this entity has on other parts of the system (Cummins, 1975). This type of functional talk does not imply norms: it does not specify what the entity should do in this particular system. But Lovelock, even though appealing to this cybernetic tradition, meant something more when he talked about the ‘function’ of living parts of Gaia. He was also using a biological meaning of function.
Lovelock attributed function to Gaian parts just as we do spontaneously with any other living forms when we speak about (for example) the function of organs within a body. In this context, attributing a function to an entity is normative: when we say that it is the function of the heart to pump blood, we mean that the heart is supposed to pump blood. This triggered Dawkins’ (1982) principal objection to Gaia: functional talk is only scientifically sound when it applies to entities subject to natural selection – and in his view, being a population of one with little sign of reproduction, Gaia was not subject to natural selection.
But of course the issue of teleology, even for organisms (i.e. life) – is a very difficult one. It is embedded within discussions from 18th century natural theology, where the functions of organs within organisms or of species at the surface of the Earth were designed by God or where the apparent design of a biological entity was used to prove the existence of God. It is also linked with the recognition of the specificity of life (small l) as a particular class of entity radically different from inorganic entities. Reflections about the specificity and status of organisms led to the constitution of biology as a discipline and were accompanied by broader reflections about the order of nature – most famously deployed by Kant (Huneman, 2008). Even though Darwin is often said to have solved the biological issue of organisms’ design, philosophers were, at the time Dawkins was writing, still struggling hard to make sense of biological teleology, even for supposedly ‘easy’ cases where natural selection occurs (Millikan, 1989; Neander, 1991; Wright, 1973). Hence, the dismissal of this issue when an entirely new biological entity had been discovered was at best premature (Latour and Lenton, 2019).
Biologists plead for mechanism, Earth system scientists miss the need for theory
The teleological critique was linked to a theoretical one: biologists argued that a mechanism producing global regulation was lacking. Such a mechanism would need to do the same explanatory work as natural selection does in explaining the adaptation of organisms and legitimising functional talk in biology. But they could not see what such a mechanism be, if natural selection cannot operate on Gaia because Gaia is not part of a population and does not reproduce (Dawkins, 1982). Instead one or two, such as W. D. Hamilton, realised there might be something somewhat akin to natural selection (Lenton, 2005) – that is, a filtering of variation in which regulatory variants came to persist and therefore predominate (as opposed to members of a population that leave the most descendants coming to predominate).
Over in ESS, there was never this recognition of the need for an ‘evolutionary’ mechanism to explain Gaia. Instead critics of the Gaia hypothesis within ESS put forward examples of ‘destabilisation’ by Life (Kirchner, 1989; Tyrrell, 2013), for example, oxygenic photosynthesis introduced a poison for most of the biosphere at the time; the spread of oxygenic photosynthesis has initiated glaciations, possibly including ‘Snowball Earth’ events (Kopp et al., 2005); and disruption of the biological sulphur cycle may have been responsible for the largest Phanerozoic extinction (Ward, 2009). But one need not wait for Kirchner (1989) or Tyrrell (2013) to find examples refuting the idea that living beings always regulate their environment – the originators of Gaia had already provided them (Margulis and Lovelock, 1974).
As Lovelock was well aware from the start, presenting cases of destabilisation as a refutation of Gaia misses a basic understanding both of biology and of cybernetics. In biology, just because we can suffer cancer does not mean we are not self-regulating organisms (Doolittle, 2019). In cybernetics, any ‘hat function’ (e.g. inverted parabola) response of Life to an environmental variable coupled with an effect of Life on that variable can give rise to both positive feedback and negative feedback regimes. Lovelock enshrined this in the formulation of the Daisyworld model (Watson and Lovelock, 1983) and has subsequently emphasised, for example, that the DMS feedback can be positive or negative depending on the circumstances (Lovelock and Kump, 1994). Lovelock’s second book ‘The Ages of Gaia’ (Lovelock, 1988) hinges on this recognition – intervals where destabilisation predominates mark the boundaries between the ‘Ages’ in which regulation predominates.
‘If Gaia stabilizes and destabilises, is there any possible behaviour that is not Gaian’ asked Kirchner (1989), in response. Others have suggested that Gaia’s validity could be resolved by simply tallying up stabilising and destabilising feedback mechanisms (Ward, 2009). This recurring argument highlights the fact that the majority of Earth system scientists discussed Gaia at an empirical level and not a theoretical one. Of course, there was theoretical work on other aspects of the Earth system – including how humans could maintain their own conditions of planetary habitability (Schellnhuber, 1999; Schellnhuber and Wenzel, 1998) – but little theoretical engagement with Gaia. Meanwhile empirical examples of global regulation were never going to be sufficient to answer biologists – because their true interest lay in abstract properties of living entities. The only way out of this impasse was to theorise about Gaia.
Models and theory in response
After biologists had abandoned the stage, and Earth system scientists had largely declined to enter it, a small band took up the challenge of theorising about Gaia.
The Daisyworld model (or ‘parable’) was formulated to demonstrate an abstract mechanism by which living entities, could, in principle regulate the global environment without any teleology or inconsistency with natural selection (Watson and Lovelock, 1983). This model rapidly became the centre of theoretical discussion about Gaia (Betts and Lenton, 2007; Lenton et al., 2018a; Lenton and Lovelock, 2001; McDonald-Gibson et al., 2008; Volk, 2002; Watson and Lovelock, 1983). After it was pointed out that the first version of the model represented a special case of coupling between life and its environment (Kirchner, 1989), theoreticians focused their efforts on understanding the conditions under which a mechanism is regulatory or destabilising. Hence, they analysed the diversity of dynamic behaviours in a Gaia system (oscillations, catastrophes, tipping points between regimes of stability, etc.), how to shift from one behaviour to another, how frequent and likely is such behaviour and so on (Lenton et al., 2018a; Wood et al., 2008). This could now be described as a complex systems approach (Scheffer, 2009) – although Daisyworld significantly predates the rise of ‘complexity science’.
Answering the persistent confusion of Gaia with altruism, it was recognised that large-scale and long timescale effects of Life ought to be based on by-products of selection (e.g. the production of oxygen is a by-product of oxygenic photosynthesis, which is not selected for oxygen production but for energy capture; Lenton, 1998; Volk, 1998; Wilkinson, 1999). Constraints were also put on the conditions for regulation to emerge: for example, the evolutionary dynamics of the trait responsible for the modification of the environment ought to be faster than that of the environmental preferences of the living entities (Lenton and Lovelock, 2001; McDonald-Gibson et al., 2008; Robertson and Robinson, 1998; Wood et al., 2006).
At smaller space scales and shorter timescales, elements of a Gaia theory at least ‘compatible’ with natural selection were sought. This meant either extending or hinging on standard selection mechanisms such as group selection (Williams and Lenton, 2008); introducing concepts foreign to evolutionary thought such as ‘feedback on growth’ (a trait benefitting indifferently to all living entities without any differential – i.e. selective – effects; Lenton, 1998; Williams and Lenton, 2008) or extending the conditions of application of selection mechanisms.
Recognising that critics of Gaia may have presumed too narrow a definition of how evolution can occur – requiring variation within populations and replication with heritability – two independent arguments have been made for cruder filtering mechanism not requiring reproduction: the idea of ‘sequential selection’ (Betts and Lenton, 2007; Lenton et al., 2018a), and that of ‘selection by survival alone’ that is, based on persistence through time alone – which marked the return of Doolittle (2014), hinging on Bouchard (2014). In the sequential selection mechanism, the central idea is that through a series of trials/experiments over time, Life’s effects on its environment could have stumbled upon stable attractors, which by definition then tend to persist. Selection based on persistence alone (Doolittle, 2014) then gives the opportunity for the resulting Gaia phenomenon to acquire further persistence-enhancing (stabilising) mechanisms. Subsequently, Doolittle has offered either biogeochemical cycles (Doolittle, 2017) or clades including Life (Doolittle, 2019) as relevant units of selection based on differential persistence – noting that both form populations of a sort.
Following a tradition of theoretical biology, all these researches put forward some specific properties of life, which are both proper to living entities and necessary for regulatory mechanisms to emerge: (1) the peaked response of growth to the variation of the environment; (2) the inevitable effect of metabolism on the environment and (3) the capacity for extinction and the ability to restart.
The ongoing need for synthesis
Retrospectively, one can look at these modelling and theoretical endeavours as descriptions of the abstract properties of Life and of Gaia – that is, Life plus its effects on habitability. The decades encompassing the constitution of biology at the turn of the late 18th century were a time of active reflection about the abstract properties of life (from the specificity of ‘organisms’ to the nature of causality at stake in biology) and a time of intense empirical observations and experiments (e.g. the advent of comparative and experimental embryology; of comparative anatomy and palaeontology and of physiology and modern medicine). Similarly, here, the recognition of the existence of Life and of Gaia required and still requires empirical, theoretical and philosophical efforts.
Summary
Gaia has been famously criticised by biologists and by Earth system scientists. Biologists pointed out the lack of theoretical mechanisms for regulation, tied in with a critique of Lovelock’s teleological language when he took too seriously the idea that Gaia is alive. Empirical examples of global regulation were of little interest to them, and having missed the novelty of the entities Life and Gaia, they were perhaps too prompt to dismiss the theoretical issues as unsolvable. By contrast, Earth system scientists, having granted that living beings are part of the Earth system, largely missed the theoretical and philosophical challenges raised by the centrality of Life in Gaia. In so doing, they largely restricted themselves to empirical discussions about how the Earth system actually works. If they deployed theoretical efforts at all – of the kind at stake in Daisyworld, that is, playing with toy models to explore the possible space of behaviours – these efforts, even though influenced in some way by Gaian research, were tied to the understanding of the Earth system humans inhabit and to consequences of this understanding for future global changes (Schellnhuber and Wenzel, 1998).
The response to these critiques has generally been: (1) to repent from teleological language and (2) to point to one important success of Gaia with the constitution of ESS. This article has taken these two arguments backwards: (1) the teleological dimension of Gaia must be confronted directly, rather than being avoided to please biologists; (2) ESS, though influenced by Gaia in many ways, has lost sight of the central position of Life.
Lovelock’s central contribution, aided by Margulis, was to discover a new living entity; Life. Too big and too foreign to be studied by biologists; too hard to spot in the energy balance to be detected and taken into account by early climatologists; too living to be fully embraced by Earth system scientists. Yet Life massively alters its global environment. So much so that it becomes hard to separate Life’s boundaries from its environment, hence the name given to the complicated entity which results from these interactions: Gaia. Studying the physiological and ecological behaviour of this entity has been the central Gaia research programme: could Life maintain its own condition of existence? What are the general properties and behaviour of a global entity constituted by the interaction of Life and its environment?
The genius of Lovelock and Margulis was to detect the hard to spot Gaia amid the Earth system. Two issues merit further discussion which we leave for further papers: a more detailed history of ESS and its relationship with Gaia and a serious discussion of Gaia’s teleology, linking the theoretical efforts developed by the Gaian scientific community with philosophical debates on causality and on the way Gaia has changed what we mean by ‘life’.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Bruno Latour benefitted from a fellowship from Zentrum für Kunst und Medien, Karlsruhe.