
Abstract
Objectives:
To better understand the sleep–wake cycle characteristics in the female Neotropical rodent Proechimys guyannensis related to comparative neurobiology.
Methods:
Surface neocortical and hippocampal electrodes were chronically implanted in the brains of female Wistar and Proechimys animals. In addition, electrodes for the study of muscle activity were implanted into the neck muscle of both species. After surgical recovery and a period of adaptation, animals were continuously registered for periods as long as 48 h.
Results:
In both the light and dark phases of the cycle, significant differences in some electrographic patterns were observed between the Proechimys and Wistar animals. Although Proechimys has nocturnal activities and a pattern of polyphasic sleep similar to Wistar rats, the analysis of its sleep-wakefulness cycle indicates that the Neotropical rodent sleeps less with consequent longer periods of wakefulness when compared to Wistar rats.
Conclusions:
Together with previous findings of different neuroanatomical, neurophysiologic and behavioral characteristics, this study allow us to better understand adaptive differences of the Neotropical rodent Proechimys.
Introduction
Organisms as distant from humans as fruit flies have been used to provide knowledge about the development and complexities of the sleep cycle. The use of simple organisms has the advantage of the lower complexity of the brain, since their central nervous system has fewer neurons and consequently a neural network that allows, to certain extent; recognize the activity of each single element. Furthermore, and associated with genomic approaches, these studies have helped to unravel the mechanisms related to the sleep–wake cycle and to provide an evolutionary perspective of the biological rhythms.1–3
To our knowledge, Weir 4 was the first to carefully describe the behavioral and morphological characteristics of the rodents of the genus Proechimys, of family Echimydae, suborder Hystricomorpha (named casiragua in the Brazilian Amazonia) maintained in captivity after being captured in the tropical rainforest since 1969, although this species has been used in scientific research since the 1950s.4–10
Our laboratory has shown that although Proechimys guyannensis are able to show convulsive behavior and status epilepticus of long duration under certain treatments (intrahippocampal kainic acid or systemic pilocarpine), these animals are resistant to the further development of chronic epilepsy suggesting the possibility of endogenous antiepileptogenic mechanisms.11,12 This finding prompted us to further investigate morphological and physiological characteristics of the P. guyannensis brain in an attempt to reveal the mechanisms underlying the so-called epileptogenic resistance. In summary, these studies have revealed that P. guyannensis’ brain presents striking differences both morphologically and physiologically when compared to the laboratory Wistar rats (WRs). These changes include, among others, the different distribution of γ-aminobutyric acid (GABA), opioid and cannabinoid binding, 13 an anatomically distinctive hippocampal cornu ammonis 2 (CA2) subfield 14 and the presence of large pyramidal-like neurons throughout the stratum oriens from CA2 to CA1 area with strikingly different electrophysiological properties. 15
Accordingly, this article describes and compares the main characteristics of the sleep–wake cycle in the P. guyannensis to those classically reported for the WRs.
Methods
Animals
Eight female P. guyannensis and eight female WRs, weighing between 240 and 280 g, were used in this study. Animals were housed under environmentally controlled conditions (22°C ± 1°C), a 12/12 h light–dark cycle (lights on at 7 a.m.), with water and food ad libitum. All procedures involving the animals and their care were performed in accordance with the Ethical and Practical Principles of the Use of Laboratory Animals16,17 and the experimental protocol was approved by the Ethical Committee of UNIFESP (CEP 2132/11). All precautions were taken for reducing the number of animals.
Electrode implantation
Animals were deeply anesthetized with ketamine (40 mg/kg intraperitoneal (i.p.)) and xilazine (5 mg/kg, i.p.). For deep electrographic recordings (EGRs), bipolar stainless steel electrodes were implanted in the right hippocampus under stereotaxic guidance. For surface EGRs, two screw electrodes were bilaterally fixed in the frontal area of each hemisphere. Electrodes for electromyography (EMG) were surgically implanted into the neck muscle. The electrodes were then soldered to a 6-pin socket and fixed to the skull of animals with acrylic cement. All recordings were registered in the individual home-cages of each animal and lasted up to 48 h. Recordings were made using a Nihon Koden Co apparatus model QP 223A (Tokyo, Japan). Signals were acquired with rate sampling of 256 Hz, scale of 100 µV and 1 s. Brain recordings were amplified and filtered at 0.1 s (1.6 Hz) and 35 Hz, high-pass and low-pass, respectively, while EMG was filtered at 0.3 s (0.53 Hz) and 70 Hz. EGRs were analyzed using Neurotronics™ Inc program (Neurotronics®, FL, USA). Sleep–wake phases were characterized according to Timo-Iaria et al. 18 The following parameters were analyzed: the percentage of total time of wakefulness (TTWS), total time of slow wave sleep (TTSWS) and total time of paradoxical sleep (TTPS) based on changes in the EGRs and EMG in both species.
Statistical analysis
Comparisons between animal groups, and between the light and dark phases in the same group were analyzed using the independent and dependent “t” test, respectively. Data are presented as mean ± standard error (SE) and the significance level was established at 5%.
Results
In the rodent P. guyannensis, the EGRs obtained both in the brain and in the muscles during the different stages of sleep and wakefulness were similar to those observed and extensively published for the WR (Figure 1(a)–(f)). In summary, the wakefulness of these animals was characterized by fast (desynchronized) activity in the neocortex, and theta activity in the hippocampal region accompanied by fast electromyographic activity during exploratory behavior. Slow wave sleep (SWS) was characterized by accentuated reduction in muscle tonus concomitantly with the prevalence of delta waves and other graph elements in cortical and hippocampal areas. During the paradoxical sleep (PS) period, the cortex expressed desynchronized activity and theta rhythm in the hippocampus with almost complete disappearance of the muscle activity. 18

Electrographic recordings of sleep-wakefulness cycle in (a–c) Proechimys guyannensis and (d–f) Wistar rat. (a and d) Alertness with desynchronized recordings and increased muscle tone, (b and e) slow waves sleep with delta waves and paradoxical sleep with cortical and hippocampal desynchronization and (c and f) suppressed muscle activity.
P. guyannensis presents a pattern of sleep and nocturnal activities that can be considered polyphasic (Figure 2(a) and (b)). These animals slept for about 45% of the time during the light phase and 28% of the time during the dark phase of the cycle (Figure 3).

Hypnogram showing the polyphasic pattern obtained during the light period in (a) Proechimys guyannensis and (b) Wistar rat.
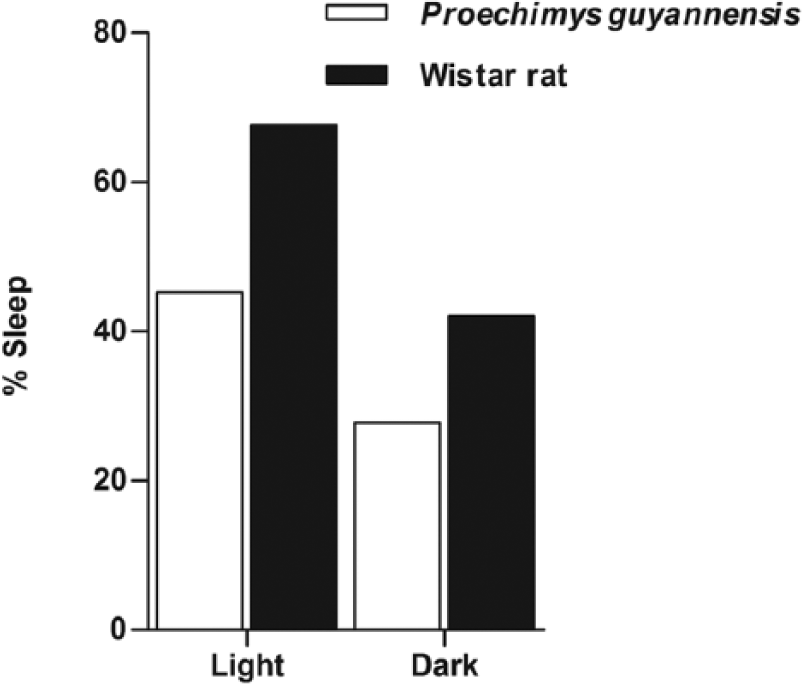
Percentage of the sleeping time during the light and the dark periods of the cycle in the Proechimys guyannensis and Wistar rat.
In comparison, WR slept 68% and 42% in the light and dark phases of the cycle, respectively. In addition, wakefulness periods in P. guyannensis represented 55% in the light and 72% in the dark. In WR, these periods represented around 26% and 56% in the light and dark phases, respectively.
Several aspects of the cycle were significantly different between P. guyannensis and WR animals during the light period. These differences included the reduced sleeping time, increased wakefulness, increased duration of SWS, increased latency and decreased duration of the PS and increased awakenings in P. guyannensis when compared to the WRs (Figure 4(a)–(f)).
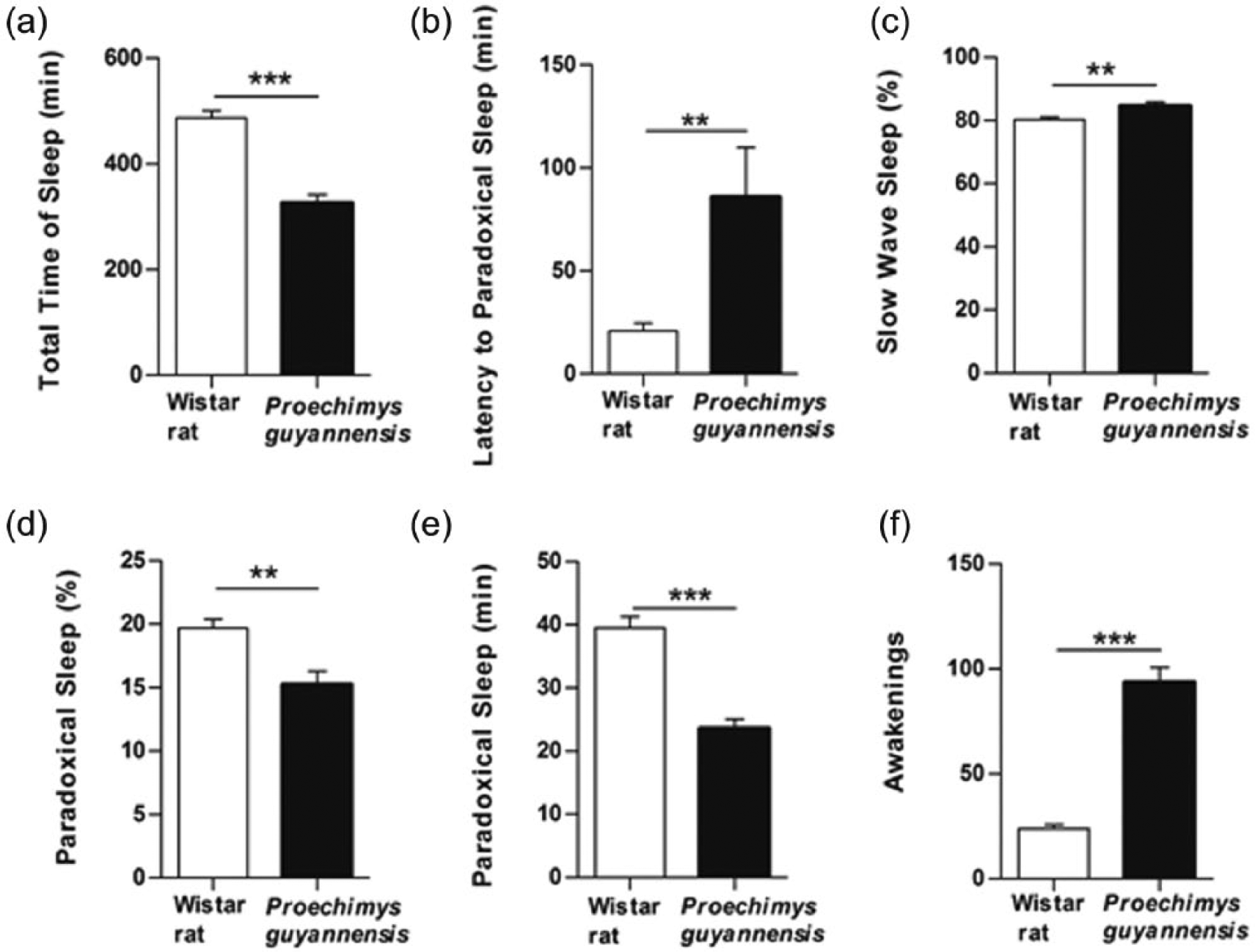
(a–f) Stage parameters in the light period of sleep-wakefulness cycle in Proechimys guyannensis and Wistar rat. **p < 0.001; ***p < 0.0001.
One aspect that could be considered similar for both species was the latency for the initiation of the SWS. During the dark period, differences in the sleep-wakefulness pattern were less pronounced between the two species (Figure 5(a) and (b)).

(a) Sleep time and (b) awakenings episodes in the dark period observed in Proechimys guyannensis and Wistar rat. Mean ± SE; ***p < 0.0001.
Lower total sleep duration, higher waking percent and increased awakenings continued to characterize the pattern of P. guyannensis in comparison with WR. Important aspects of the sleep-wakefulness cycle were significantly different between the light and dark period in P. guyannensis animals such as time of sleep and wakefulness, latency to PS, awakenings in PS (Figure 6(a)–(d)).
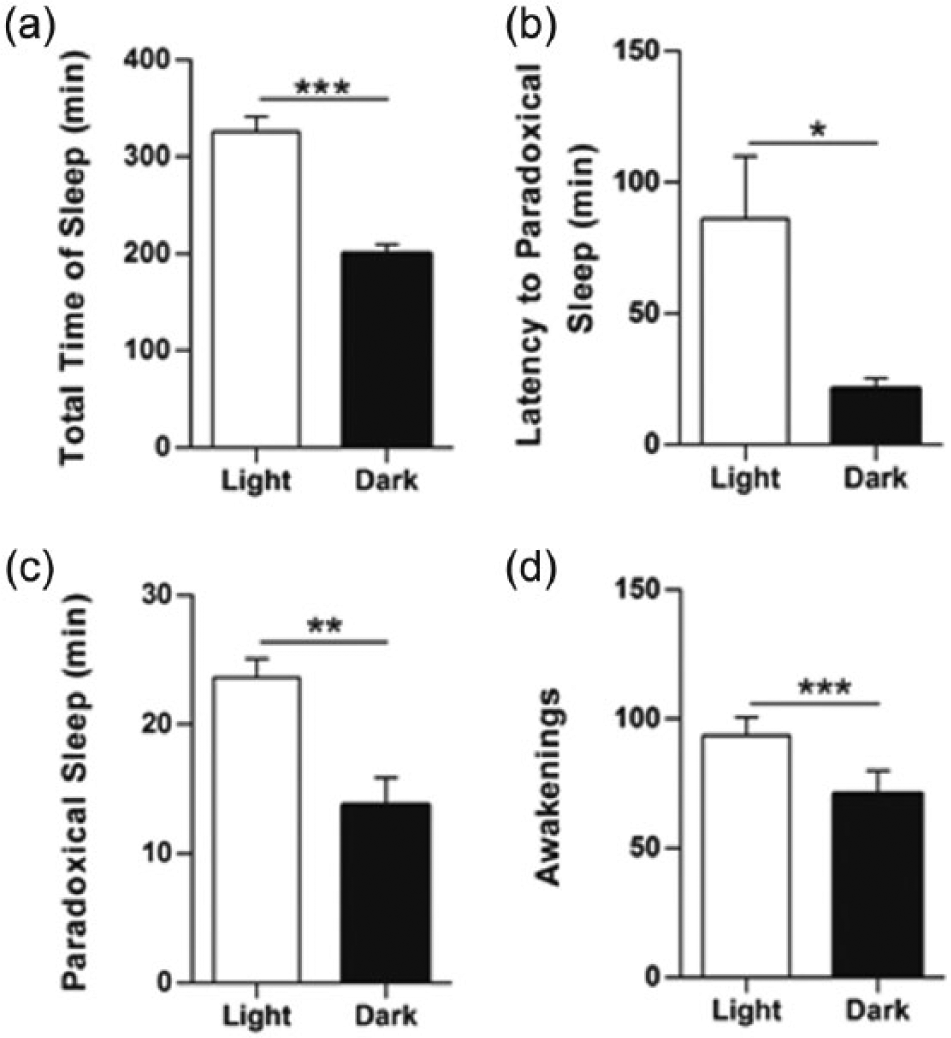
Sleep-wakefulness cycle in the Proechimys guyannensis. Notice in (a) that sleep duration during the light period is longer than in the dark period. In (b–d), it is shown that although the latency to PS is smaller during the dark period, the total duration of PS and number of awakenings are reduced in the same part of the cycle. Mean ± SE; *p < 0.01; **p < 0.001; ***p < 0.0001.
Another important difference between the animal species used in this study relates to the behavior during sleep. While the WR presents ventroflexion of the head towards the trunk, P. guyannensis remains lying on the belly with stretched head.
Discussion
The pattern of sleep-wakefulness cycle in the Neotropical rodent P. guyannensis as in the WRs is polyphasic.18,19 Besides, our data showed that P. guyannensis compared to WR, sleep less, which results in more and longer lasting periods of wakefulness during the entire cycle.
The first hypothesis that could be used to explain the differences found in the pattern of sleep–wake cycle of the P. guyannensis in relation to WR is related to the fact that these animals belong to phylogenetically distant branches. While this hypothesis may not be readily discarded, several authors have considered this shortening of the sleep period as a necessity for survival in the wild. It is very likely that this adaptation in response to the presence of predators is already incorporated into the molecular and genetic pattern of these species.5,20 However, one cannot rule out, as reported by some authors, that these changes, predominantly those related to the dark period, as being related to environmental exploration, searching for food, social interaction, procreation and so on.4,6,8,10,21,22
The methodology used in this study does not allow a more detailed discussion of the mechanisms underlying the differences in the sleep patterns presented by P. guyannensis compared to WRs. Furthermore, this study was conducted in female animals and any more accurate digression could only be made if these findings are confirmed for male animals. However, this study, taken together with other already conducted by our group in the Neotropical rodent P. guyannensis, may contribute for the understanding of the important morphological and physiological differences present in the brain of this species when compared to other species commonly used in laboratory.
Footnotes
Acknowledgements
The authors would like to express their thanks to T.A.F. Alvarenga.
Declaration of conflicting interests
None of the authors have any conflict of interest to disclose.
Funding
This work was supported by grants from the Associação Fundo de Incentivo à Psicofarmacologia, Fundação de Amparo à Pesquisa do Estado de São Paulo (#10/15110-8 to G.M.), CInAPCe-FAPESP, CNPq and FAPESP/CNPq/MCT-Instituto Nacional de Neurociência Translacional. J.C. Silva is fellow from CNPq (Brazil).