
Abstract
Curcumin was previously reported to accelerate neutrophil apoptosis, but the mechanism is unclear. Herein, we confirmed that curcumin induces human neutrophil apoptosis as assessed by cytology and by increase in the cell surface expression of annexin-V and CD16 shedding. Curcumin activated caspase-3 and the cleavage of the two cytoskeletal proteins lamin B1 and vimentin. In addition, curcumin activated protein kinase RNA-like endoplasmic reticulum kinase and eukaryotic initiation factor 2 alpha and reduced de novo protein synthesis and the protein expression of the two chaperone proteins, HSP70 and HSP90. We conclude that curcumin acts as an endoplasmic reticulum stressor in human neutrophils. The ability of curcumin to activate the endoplasmic reticulum stress-induced cell apoptotic pathway is part of its mode of action in primary cells like mature neutrophils.
Introduction
Polymorphonuclear neutrophil cells (PMNs) are the most abundant cell type of the immune system. They are phagocytes that undergo apoptosis spontaneously and play a key role in inflammation, being among the first cells to arrive at an inflammatory site. In addition, PMNs are the main source of preformed inflammatory mediators and proteolytic enzymes contained in the granules.1,2 The rate of apoptosis of PMNs is of great importance in the regulation of inflammation since this rate may vary under different activation states, resulting in recurrent infections (high or increased apoptotic rate) or autoimmune disorders and persistent chronic inflammation (low or decreased apoptotic rate).2,3 Therefore, identifying new apoptotic regulators and understanding the mode of action of some previously identified factors represent important potential therapeutic targets for control of inflammation.
For centuries, curcumin (Cur) has been used to treat many human autoimmune and chronic inflammatory diseases, including type I diabetes, 4 arthritis, 5 psoriasis 6 and inflammatory bowel disease. 7 Cur is, in fact, used as a general anti-inflammatory agent, but its precise mechanism of action is not completely understood. There are many molecular targets of Cur, including transcription factors, protein kinases, enzymes, growth factors, anti-apoptotic proteins and inflammatory mediators.8,9 Cur exerts antioxidant, antibacterial, antifungal, antiviral, anti-inflammatory, antiproliferative and proapoptotic effects.8,10 Mostly, cell death induced by Cur treatment is called apoptosis. 11 It was previously reported that the proapoptotic activity of Cur involves activation of caspases and mitochondrial dysfunction triggered by enhanced Bax expressions in cell lines.12,13 Cur was identified recently as a PMN proapoptotic molecule acting by a p38-dependent mechanism involving caspase-3 activation. 14 Interestingly, involvement of the endoplasmic reticulum (ER) stress-induced apoptosis pathway has been recently demonstrated in Cur-induced promyelocyte HL-60 cells. 11 Since we have recently documented that the ER stress-induced cell apoptosis pathway is functional in arsenic trioxide (ATO)–induced human PMNs, 15 and since the mode of action of Cur is unclear, we decided to investigate the possibility that Cur induced PMN apoptosis by altering the ER stress-induced cell apoptotic pathway.
Materials and methods
Chemicals
Cur, ATO, cycloheximide and dimethyl sulphoxide (DMSO) were purchased from Sigma Chemical Company (St Louis, MO, USA). RPMI 1640 and Hank’s balanced salt solution (HBSS) were purchased from Calbiochem International (La Jolla, CA, USA).
Human neutrophil isolation
PMNs were isolated from the venous blood of consenting healthy individuals by dextran sedimentation followed by centrifugation over Ficoll-Hypaque (Amersham Pharmacia Biotech Inc., Baie-d’Urfé, QC, Canada) according to our institutionally approved procedures.16,17 Cell viability (>99%) was monitored by trypan blue exclusion, and the purity (>98%) was verified by cytology from cytocentrifuged preparations coloured by the Hema 3 Stain Set (Biochemical Sciences Inc., Swedesboro, NJ, USA).
Assessment of neutrophil apoptosis by cytology
Briefly, freshly isolated human neutrophils (used at 1, 5 or 10 × 106 cells/mL in RPMI 1640 supplemented with 10% autologous serum) were incubated for different periods of time in the presence or absence of various concentrations of Cur (1–100 µM). The proapoptotic drug ATO (5 µM) was used as a positive control. 18 Cytocentrifuged preparations of neutrophils were performed and processed as documented previously.16,17
Assessment of apoptosis by flow cytometry
CD16 expression and annexin-V binding were used to confirm the ability of Cur to induce PMN apoptosis. After 24 h of incubation with various concentrations of Cur, the cells were suspended at concentrations of 10 × 106 cells/mL, washed and pre-incubated for 30 min at 4°C (light protected) with 20% autologous serum to prevent non-specific binding via Fc receptors (FcRs). The cells were then washed, resuspended at 1 × 106 cells/mL and incubated with 2 µL of the fluorescein isothiocyanate (FITC)–mouse anti-human CD16 mAb for 30 min at 4°C (light protected) or with 2 µL of FITC-annexin-V for 15 min at room temperature (light protected) prior to fluorescence-activated cell sorting (FACS) analysis. 17
Processing of caspases in Cur-induced human PMNs
Cells were treated with cur for different periods of time (15 min to 4 h), and processing of caspase-3 was assessed by monitoring the degradation/cleavage of the procaspase-3 form by immunoblotting, using a rabbit anti-caspase-3 antibody (BD PharMingen, Mississauga, ONT, Canada). 18
Western blot
Neutrophils (10 × 106 cells/mL in a 96-well plate) were incubated in the presence or absence of 10 or 50 µM Cur for 24 h and then harvested for the preparation of cell lysates in 4X Laemmli sample buffer. Aliquots corresponding to 500,000 cells were loaded onto 10% sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred from gel to polyvinylidene difluoride (PVDF) membranes. Non-specific sites were blocked with 3% bovine serum albumin (BSA) in Tris-buffered saline (TBS)-Tween (0.1%) for 1 h at room temperature. Membranes were incubated with monoclonal anti-human lamin B1 (1:1000, clone C-20; Santa Cruz Biotechnology, Inc., Santa Cruz, MA), anti-human vimentin (1:1000, clone V9; Santa Cruz Biotechnology, Inc.), anti-heat shock proteins (HSPs – HSP70 (SPA-810) and HSP90 (SPA-830)), purchased from Stressgen (Ann Arbor, MI, USA), overnight at 4°C. The membranes were then washed with TBS-Tween and incubated with appropriate horseradish peroxidase (HRP)-conjugated secondary antibodies directed against the respective primary antibodies for 1 h at room temperature in TBS-Tween. Protein expression was revealed using enhanced chemiluminescence (ECL)-Western blotting detection system. Protein loading was verified by staining the membranes with Coomassie blue at the end of the experiments. Densitometry analysis was performed using the FluorChem2 software.
Metabolic labelling and de novo protein synthesis
Cells (10 × 106 cells/mL in RPMI 1640 medium without methionine and supplemented with 10% autologous serum) were metabolically labelled with 4625 MBq of the Redivue Pro-Mix L-[ 35 S] in vitro cell labelling mix (PerkinElmer, Boston, MA) in the presence or absence of agonists, as indicated in the appropriate figure legends, for 24 h, as previously described.18,19 Newly synthesized polypeptide was then visualized by autoradiography after separation of proteins by electrophoresis.
Phosphorylation events
Neutrophils (40 × 106 cells/mL in RPMI 1640–4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES)–penicillin/streptomycin (P/S)) were stimulated from 1 to 5 h with the agonists. Cells were lysed in 4X Laemmli sample buffer, and aliquots corresponding to 1 × 106 cells were loaded onto 10% SDS-PAGE and transferred to PVDF or nitrocellulose membranes. Membranes were blocked for 1 h at room temperature in TBS-Tween containing 5% non-fat dry milk (Carnation, Don Mills, ON, Canada). After washing, the anti-phosphospecific eukaryotic initiation factor 2 alpha (eIF2α) (1:1000; Cell Signaling Technology, Inc., Danvers, MA) or protein kinase RNA-like endoplasmic reticulum kinase (PERK) (1:1000; Santa Cruz Biotechnology, Inc.) antibody was added at a final concentration of 1:1000 in TBS-Tween. The membranes were incubated overnight at 4°C and then washed with 0.1% of TBS-Tween and incubated for 1 h at room temperature with a goat anti-rabbit HRP-conjugated secondary antibody (Jackson ImmunoResearch Laboratories, West Grove, PA, USA) at a final concentration of 1:25,000 in TBS-Tween. Actin expression was assessed after stripping of membranes and staining with an anti-actin antibody (clone AC-40; Sigma-Aldrich Canada Co., Oakville, ONT). Protein expression was revealed using an ECL-Western blotting analysis detection system.
Statistical analysis
The data are reported as mean ± standard error of mean (SEM). Analysis of variance was used to compare the means of different treatments. If significance was identified, individual comparisons were subsequently made by Bonferroni test for unpaired values. Statistical analyses were performed using GraphPad Prism version 5.00 for Windows (GraphPad Software, San Diego, CA, USA). Statistical significance was set at p < 0.05.
Results
Cur induces apoptosis of PMNs in a cell- and concentration-dependent manner
As illustrated in Figure 1(a), the rapid PMN proapoptotic activity observed after 6 h of treatment with 50 µM Cur, as assessed by cytology, was significantly increased relative to spontaneous apoptosis (SA), but only when the cell concentration was adjusted to 1 × 106 cells/mL. As the concentration of cells was increased, the proapoptotic activity of Cur decreased; this was particularly significant when PMNs were plated at 10 × 106 cells/mL. However, as illustrated in Figure 1(b), after 24 h of treatment, Cur significantly increased the apoptotic rate when cells were plated at 1, 5 and 10 × 106 cells/mL. We then determined the concentration-dependent proapoptotic activity of Cur when cells were plated at 10 × 106 cells/mL, a cell concentration routinely used to obtain 30%–50% of apoptotic PMNs during SA after 24 h. As illustrated in Figure 2(a), Cur significantly increased apoptosis in a dose-dependent manner, except at the lowest concentration of 1 µM. As expected, ATO induced PMN apoptosis.15,18,20 In parallel, we tested the potential necrotic effect of Cur, curiously undocumented before this study, and observed that necrosis never exceeded ~5% for all tested concentrations, except for 100 µM Cur, where about 20% of cells were necrotic, as assessed by trypan blue exclusion assay (inset, Figure 2(a)). Therefore, except where indicated, Cur was used at a concentration of 50 µM (Cur50) in subsequent experiments. We then confirmed the above-mentioned results by flow cytometry. As expected, the number of annexin-V positive cells was increased by Cur treatment when compared to SA and, conversely, was decreased when cells were treated with the anti-apoptotic cytokine granulocyte-macrophage colony-stimulating factor (GM-CSF) (Figure 2(b)). As expected, the number of CD16 negative cells 21 was increased by Cur treatment and decreased by GM-CSF versus SA (Figure 2(c)).

Modulation of the human PMN apoptotic rate by curcumin. Fresh human neutrophils were isolated and incubated at 1, 5, or 10 × 106 cells/mL for (a) 6 h and (b) 24 h in the presence or absence of 50 µM curcumin (Cur), and apoptosis was assessed by cytology as described in the ‘Materials and methods’ section. Results are means ± SEM (n ≥ 3).
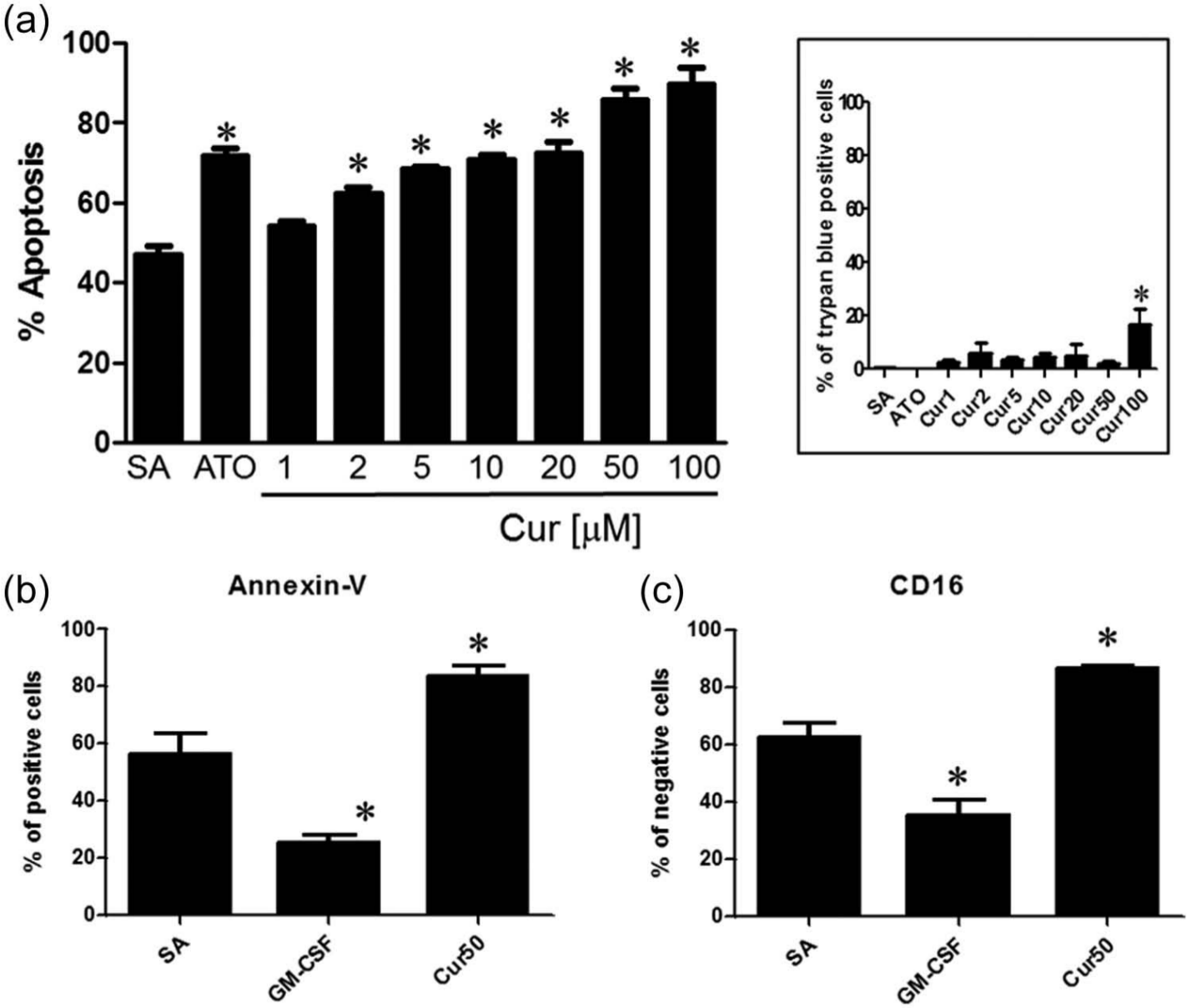
Curcumin accelerates human PMN apoptosis in a concentration-dependent manner. Cells (10 × 106 cells/mL) were incubated for 24 h with buffer (SA), 5 µM ATO (positive control) or an increasing concentration of Cur, and apoptosis was assessed by (a) cytology or (b and c) flow cytometry after staining with (b) annexin-V or (c) CD16 as described in the ‘Materials and methods’ section. Inset in (a) – cell viability was evaluated by trypan blue exclusion.
Cur activated caspases and the degradation of cytoskeletal proteins but inhibited de novo protein synthesis in human PMNs
As illustrated in Figure 3(a), Cur induced the process of procaspase-3 within 2 h of treatment. We then determined whether or not Cur could induce degradation of lamin B1 and vimentin, two proteins known to be cleaved by caspases during PMN apoptosis. Cur10 slightly to moderately induced the degradation of these two proteins (Figure 3(b)), whereas Cur50, like ATO, drastically degraded both lamin B1 and vimentin when compared to SA, indicating that apoptosis was accelerated by Cur in human PMNs. Because the atypical ER stressor ATO is known to induce de novo protein synthesis in PMNs,15,20 and since Cur had been found recently to induce ER stress-induced cell apoptosis in promyelocytic HL-60 cells, 11 we then examined whether or not Cur could act as a more typical ER stressor, for example, by inhibiting protein synthesis, an important step known to alleviate ER stress.23–25 As illustrated in Figure 4(a), Cur, in contrast to ATO, inhibited de novo protein synthesis, suggesting that Cur acts via a different mechanism than ATO. To support this, we investigated how Cur could alter the levels of protein ubiquitination known to be increased by ATO in PMNs. 20 As illustrated in Figure 4(b), Cur did not increase ubiquitination of proteins in the presence or absence of the MG132 proteasome inhibitor, and, in fact, it slightly decreased this process.

Curcumin induces the processing of caspase-3 and the cleavage of cytoskeletal proteins. PMNs (10 × 106 cells/mL) were incubated for the indicated periods of time with 10 or 50 µM curcumin (Cur10 and Cur50, respectively), and (a) the processing of procaspase-3 (proCASP3) or (b) the cleavage of lamin B1 or vimentin (Vim) were assessed by Western blot as described in the ‘Materials and methods’ section. Results are from one representative experiment out of at least three. Arrowhead indicated the 47-kDa fragment of lamin B1 observed in HL-60, PLB-985 and human PMNs. 22
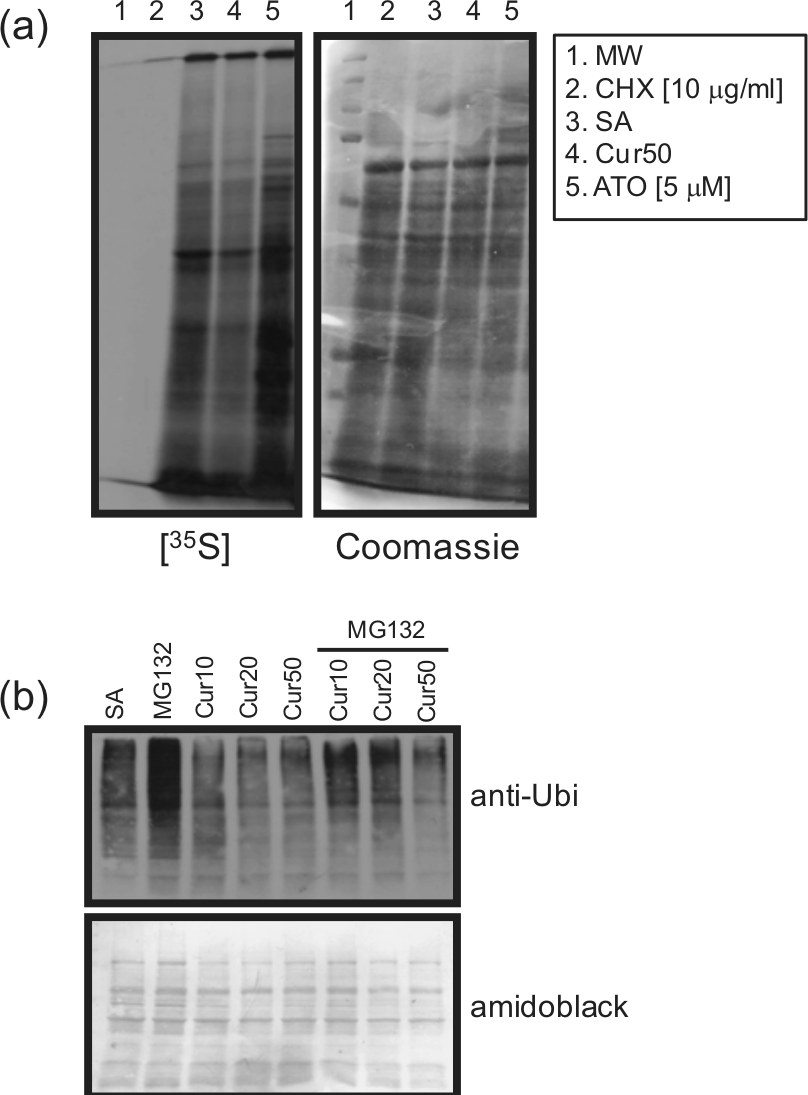
Curcumin inhibits de novo protein synthesis and protein polyubiquitination in human neutrophils. PMNs (10 × 106 cells/mL) were incubated for (a) 24 h or (b) 3 h with the indicated agonists and de novo protein synthesis ((a)), or polyubiquitination was performed as described in the ‘Materials and methods’ section. Results are from one representative experiment out of three.
Activation of the ER unfolded protein response by Cur during PMN apoptosis
The above-mentioned results prompted us to determine whether or not an ER stress occurs in response to Cur in PMNs by investigating the activation of PERK/eIF2α, an important marker of the ER unfolded protein response (UPR), leading to inhibition of protein synthesis. After 1 h of treatment, Cur increased the phosphorylation of PERK but not as strongly as thapsigargin (Tp), a classical ER stressor (Figure 5(a)). As illustrated in Figure 5(b), Cur also activated eIF2α, as evidenced by its increased phosphorylation when compared to control (or SA). Interestingly, after 1 h, eIF2α appeared to be more activated by Cur versus Tp, but in contrast to Tp, the effect of Cur was not maintained over time. The above-mentioned results indicated that Cur could activate the ER stress-induced cell apoptotic pathway in human PMNs. Knowing that unfolded proteins are first recognized by molecular chaperones, including HSP90 and HSP70, we then decided to verify whether Cur could decrease levels of expression of these two proteins. As expected, both HSP90 and HSP70 protein expressions were decreased in response to Cur, especially HSP90 (Figure 5(c)).

Curcumin activates PERK and eIF2α but decreases the expression of HSP90 and HSP70 in neutrophils. PMNs were incubated at 37°C with the diluent (1% DMSO, Ctrl or SA), 50 µM Cur or 10 µM thapsigargin (Tp) for the indicated periods of time. Activation of (a) PERK and (b) eIF2α and (c) the expression of HSP90 and HSP70 were performed as described in the ‘Materials and methods’ section. (a) and (b) – results are from one representative experiment out of four and (c) – results are from one representative experiment out of three.
Discussion
Accumulating evidence indicates that apoptosis of neutrophils is one of the critical determinants of the outcome of the inflammatory response. This process represents, therefore, a potential target for therapeutic intervention.1–3 The effects of Cur on the intrinsic and extrinsic apoptotic pathways have been well described, 26 but are still not fully understood, and have become more complex as novel molecular targets are discovered. Cur modulates numerous proapoptotic and anti-apoptotic targets, and more recently, it has been demonstrated that Cur can act as an ER stressor in HL-60 cells, 11 rabbit skeletal muscle cells 27 and in mouse melanoma cells. 28 In fact, research on apoptosis and Cur has suggested that Cur may induce apoptosis through all the main apoptotic pathways in cancer cells. The major apoptotic pathway targeted by Cur may vary between cell type, differentiation state and Cur concentration, and therefore, the precise mechanism is difficult to determine because of crosstalk between the different apoptotic pathways. 26 Although the initial study (and probably the only one yet) reported that Cur induced apoptosis in human PMNs via p38 and caspase-3 activation, the authors did not recognize or describe the rapidity with which this molecule can act. 14 Here, we present new insight into the mechanisms involved in Cur-induced apoptosis in human PMNs. The effectiveness of Cur on PMNs apoptosis is dependent on the cell concentrations at which PMNs are plated. Although we confirmed that Cur could effectively induced apoptosis rapidly after 6 h, the apoptotic rate did not reach more than 20% at a cell concentration of 1 × 106 cells/mL, as opposed to 60% when cells were plated at 2 × 106 cells/mL. 14 The other important experimental condition that differs in this study relates to temperature. The authors of the initial study kept the PMNs on ice before increasing the temperature, to that of the assay conditions, probably causing a thermal shock. There is no information regarding whether or not cells were incubated in the presence or absence of serum. In this study, cells were incubated without any thermal shock, in the presence of 10% autologous serum and at 10 × 106 cells/mL (more representative of an inflammatory response when the number of PMNs is considerably increased). Despite this, when cells were incubated under these experimental conditions, Cur still exerts potent proapoptotic properties, and here, we show new mechanisms related to its mode of action during PMN apoptosis. We confirmed the activation of caspase-3 in Cur-induced PMNs, but in addition, we show that lamin B1 and vimentin are cleaved in Cur-induced PMN apoptosis, supporting the role of caspases in Cur-induced cells,12,29–31 including PMNs. 14
Activation of the sensors of unfolded proteins localized in the lumen of the ER, such as PERK leading to activation of eIF2α, suggests that unfolded proteins accumulate in the ER of Cur-induced PMNs. Normally, accumulation of unfolded protein in the ER forces cells to respond by one mechanism devoted to stop protein translation, avoiding overloading of the ER by nascent proteins. Herein, as assessed by metabolic labelling and de novo protein synthesis, Cur reduced general protein translation, probably due to the resulting activation of eIF2α, correlating with its role in attenuating protein translation. 24 In the ER, protein folding is achieved with the help of several molecular chaperones and folding enzymes. Among these proteins, HSP70 and HSP90 are important intracellular molecular chaperones with protective effects that are known to be induced in response to a variety of physiological and environmental insults.32–34 The observation that Cur induced a slight reduction in the expression of both HSP70 and HSP90 in human PMNs suggests that the proapoptotic activity of Cur is related to ER stress. Interestingly, unfolded proteins are known to be rapidly degraded in cells, and one important mechanism associated with this degradation is the ubiquitin–proteasome pathway. Therefore, chaperones like HSP70 and HSP90 may recruit E3 ubiquitin ligase in order to polyubiquitylate such proteins. 35 However, our results show that Cur decreased polyubiquitination of proteins in PMNs, suggesting a role for this mechanism probably related to its proapoptotic activity.
From the data obtained in this study, it can be concluded that the proapoptotic activity of Cur in human neutrophils is related to its ability to act as an ER stressor and to induce the ER stress-induced cell apoptotic pathway. Therefore, in addition to the results previously reported by others demonstrating that Cur is able to induce both receptor-mediated and mitochondria-mediated proteolytic pathways for apoptosis in human glioblastoma T98G cells, 12 two pathways known to be activated in human neutrophils, 15 Cur should be now considered as an agent able to activate the three major pathways of cell apoptosis known to be operational in human neutrophils, namely, the extrinsic, the intrinsic and the most recently identified ER stress-mediated pathways.15,20
Footnotes
Acknowledgements
We thank Mary Gregory for reading this manuscript.
Declaration of conflicting interests
The authors declare that there are no conflicts of interest.
Funding
This study was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC).