
Abstract
When asking spider-phobic individuals what they find most frightening about spiders, many report spiders’ unique movement characteristics. Here, we experimentally tested whether spider-specific movements are indeed a major cue in triggering spider fear. One-hundred-and-twenty participants (55 spider-fearful) viewed realistic 3D-animated videos of spiders and ladybugs moving on different paths and with different speeds. We found that spider-fearful individuals perceived spiders as more uncontrollable and unpredictable than ladybugs and non-fearful individuals. This held true to a certain degree even when spiders moved on predictable movement paths. Spiders also elicited more fear and disgust in spider-fearful individuals, which was amplified by curvy movement paths. Finally, spider-fearful individuals reported higher intolerance of uncertainty (IU), which may potentially increase distress in subjectively uncontrollable and unpredictable spider encounters. As far as we are aware, our study provides first experimental evidence of the effects of spider movement on cognition and emotion in spider fear, expanding the current knowledge about fear-related triggers.
Introduction
Spider fear is commonly regarded as a prototypical form of animal phobia, not least due to its high prevalence in the general population (Becker et al., 2007; Jacobi et al., 2004). A classical associative account to explain the acquisition and maintenance of animal phobias is preparedness theory (Seligman, 1971). With respect to spider phobia, preparedness theory states that (a) fear reactions help to avoid bodily harm (e.g., envenomation) and (b) disgust reactions help to avoid diseases carried by spiders (Rozin & Fallon, 1987). From this view, it is evolutionarily reasonable for human beings to quickly learn aversive emotional reactions to spiders. Indeed, spiders have been shown to evoke higher self-reported fear and disgust in the general population compared to other small animals (Gerdes et al., 2009). However, only very few spiders pose a real threat to humans (an estimated 0.5% of known spider species; Hauke & Herzig, 2017). Also, the probability of contracting serious diseases from a spider bite is very low (Vetter et al., 2015). Furthermore, being based on classical conditioning, preparedness theory implies the occurrence of initial aversive encounters (i.e. conditioning events) with spiders to acquire phobic reactions. Yet only a minority of spider-phobic individuals report such initial aversive encounters (<10%, Davey, 1991). A more recent, non-associative account of fear acquisition states that certain innate, evolutionary-relevant fears can also emerge without any conditioning events (Poulton & Menzies, 2002). Importantly though, this account still cannot sufficiently explain why spiders as a largely harmless species are so frequently feared compared to other small animals (see Armfield, 2006, for more details). Therefore, additional aspects, such as cognitive factors, have to be considered to explain the acquisition and maintenance of spider phobia.
Cognitive Factors: Uncontrollability, Unpredictability, and Biased Perceptions
Armfield (2006) proposed a model of cognitive vulnerability to explain the etiology and characteristics of specific phobias which relies on four cognitive factors: perceived dangerousness, disgustingness, uncontrollability and unpredictability. In the model, both perceived uncontrollability and unpredictability are inherently linked to the movement of spiders. This is the case because the model states that spider-phobic individuals develop cognitive vulnerability schemas that lead them to perceive spider movements as highly (a) unpredictable (i.e. they feel incapable of predicting where a spider might move to), and (b) uncontrollable (i.e. they feel a lack of agency over the spider’s behaviour). Indeed, individuals who were informed that a spider might behave in an uncontrollable und unpredictable manner reported higher fear ratings before and during an encounter with a fake spider (Armfield, 2008; note though that this was a non-phobic sample). Importantly, these perceptions of spider movements as highly uncontrollable und unpredictable are thought to be distorted, such that spider-phobic individuals perceive them (and, consequently, the behaviour of the spider in general) as more uncontrollable und unpredictable than is the case in reality. This may in turn lead to an overestimation of the probability of a negative outcome when encountering a spider (e.g. being suddenly attacked by the spider or involuntarily coming into contact with it). Such an overestimation of negative outcome probabilities in fear-relevant encounters has been previously referred to as consequence expectancy bias and previous research found compelling evidence for its presence in spider-phobic individuals (see Hengen & Alpers, 2019 for a recent study or Aue & Okon-Singer, 2015 for a review). Additionally, spider-phobic individuals also tend to overestimate the probability of encountering spiders in general (= encounter expectancy bias, e.g. Aue & Hoeppli, 2012; De Jong & Muris, 2002). Moreover, when faced with ambiguous images varying in their degrees of similarity to spiders, spider-fearful individuals were more likely to perceive such images as spiders compared to non-fearful individuals (= interpretation bias; Haberkamp et al., 2019). Lastly, evidence also points towards attentional biases in spider phobia which modulate perceptions. Spider-phobic individuals display an initial bias towards threatening stimuli (i.e. spiders), have difficulties in disengaging attention from these stimuli and avoid these stimuli more in later stages of visual processing (see Aue & Okon-Singer, 2015 for more details). In sum, evidence strongly suggests that several biases in perception exist in spider phobia and that they are relevant to the acquisition and maintenance of the disorder.
However, the specific movement characteristics of spiders and their relations to biased perceptions were hardly investigated in clinical research. This is somewhat surprising given that spider-phobic individuals report spider-specific movements as an important trigger of their fear (Mcnally & Steketee, 1985). Also, in a more recent study, spider-phobic individuals related their fear largely to the unique movement characteristics of spiders and provided mostly movement-related answers in free text questions such as ‘fast’, ‘legs’ and ‘move’ (Lindner et al., 2019). Again, this suggests that movement-related aspects of spiders (i.e. movement pattern and speed) might play an important role in the acquisition and maintenance of animal phobias.
In stark contrast to this evidence, hardly any experimental studies that investigated fear-evoking characteristics of spiders have used moving spiders. For example, the spider used in the study by Armfield (2008) was fake and unable to move. Other studies used in sensu encounters (e.g. Armfield, 2007; De Jong & Muris, 2002) or static images (e.g. Basanovic et al., 2019; Haberkamp et al., 2018; Vrijsen et al., 2009), making it hard or even impossible to study the role of movement-related attributes. Thus, to learn about the acquisition and maintenance of spider phobia, it is important to take into account movement characteristics of spiders and related cognitive factors, like perceived uncontrollability and unpredictability, for which it is important to use realistically moving spiders in the experimental setup.
Trait Factors: Intolerance of Uncertainty
Besides cognitive factors, trait factors related to spider movements might also influence the acquisition and maintenance of spider phobia. One such trait is intolerance of uncertainty (IU; Carleton, 2016), which is defined as ‘an individual’s dispositional incapacity to endure the aversive response triggered by the perceived absence of salient, key, or sufficient information, and sustained by the associated perception of uncertainty’ (Carleton, 2016, p. 31). IU has been shown to be a transdiagnostic risk factor for several anxiety disorders (e.g. generalized anxiety disorder, social anxiety disorder and panic disorder; Mahoney & McEvoy, 2012; Shihata et al., 2016, but see Rosser, 2019 for a critical review). In spider phobia, elevated levels of IU may increase perceived uncontrollability and unpredictability, which in turn may increase phobic reactions. However, to our knowledge, no study has examined the relationship of spider phobia and IU yet.
Taken together, even though there is ample evidence that movement-related factors play an important role in the acquisition and maintenance of spider phobia, there is little research to clarify that role. Specifically, previous studies did not use realistically moving stimuli; did not test how spider movement patterns or speeds relate to perceived uncontrollability, unpredictability and phobic reactions; and did not test the potential relationship of IU and spider fear.
Study and Hypotheses
To address these research gaps, we created novel 3D-animated videos of realistically moving spiders and ladybugs (as a non-aversive control stimulus), and conducted an online experiment with spider-fearful and non-fearful individuals. We varied animal movements in terms of their movement path (straight vs. curvy) and speed (slow vs. medium vs. fast). The straight path was designed to represent a predictable and controllable movement pattern while the curvy path was designed to represent an unpredictable and uncontrollable movement pattern. Participants responded to each video on self-report scales of perceived uncontrollability and unpredictability as well as fear, disgust, arousal and valence (cf. Haberkamp et al., 2019). We tested the following hypotheses:
Emotional Ratings of Animal Movements
H-1: Realistic 3D-animated spiders will elicit more negative emotions (fear, disgust, arousal and valence) in spider-fearful individuals compared to ladybugs and non-fearful controls. Also, the movement path and speed of realistic 3D-animated spiders will affect emotional ratings of spiders in spider-fearful individuals. Specifically, spiders moving on a curvy path will elicit more negative emotions compared to spiders on a straight path, and faster spiders will elicit more negative emotions compared to slower spiders.
Perceived Uncontrollability and Unpredictability of Animal Movements
H-2: The movement path of realistic 3D-animated animals will affect perceived uncontrollability and unpredictability of animal movements in spider-fearful and non-fearful individuals. Specifically, spiders and ladybugs moving on a curvy path will be perceived as more uncontrollable and unpredictable compared to animals on a straight path.
Intolerance of Uncertainty
H-3: Spider-fearful individuals will score significantly higher on IU measures than non-fearful individuals. We expect that IU levels in spider-fearful individuals will be correlated with emotional ratings (i.e. fear, disgust and arousal ratings) when spiders move on a curvy path. Also, IU levels will be correlated with perceived unpredictability and uncontrollability.
Method
The present study was preregistered on OSF (https://osf.io/czrpn). We provide access to data and analysis code (https://osf.io/5zwbk/) as well as the stimuli we used in our study and the script which controlled the procedure of the experiment (https://osf.io/zfj78/). The study was approved by the Ethical Committee of the Faculty of Psychology (Philipps-University Marburg, case number 2021-09k) and carried out in accordance with the provisions of the World Medical Association Declaration of Helsinki.
Participants
Note that we refer to participants in our study as ‘spider-fearful’ rather than ‘spider-phobic’ because we also included individuals not fulfilling all diagnostic criteria for specific phobia (APA, 2013). Previous research has shown that such individuals react similarly to spiders compared to individuals that fulfill all diagnostic criteria for specific phobia (Landová et al., 2021).
Before the study, we conducted a power analysis using data simulation in R. The power analysis incorporated the factors group (spider-fearful vs. non-fearful) × movement path (straight vs. curvy) × animal (spider vs. ladybug) x speed (slow vs. medium vs. fast). We mainly focused on the three-way interactions of our design in our power analysis. To the best of our knowledge, no previous study used realistically moving spiders to study movement-related aspects of spider phobia. However, studies that manipulated uncontrollability and unpredictability produced only small differences in outcome measures (Armfield, 2007, 2008). Therefore, we assumed small effect sizes of η p 2 ≥ 0.01 for the three-way interactions in accordance with the conventions by Cohen (1988). To detect such effects with a power of .80 and α-error rate of .05, the power analysis for our design determined that 110 individuals should participate in our experiment (55 spider-fearful, 55 non-fearful).
The study was advertised via mailing-lists and the online research participation platform Sona Systems (https://www.sona-systems.com). All individuals gave written informed consent. Individuals were eligible for participation if they were classified as spider-fearful or non-fearful (determined by scoring above the 75th percentile to be deemed spider-fearful or below the 25th percentile to be deemed non-fearful, using gender specific norm values of the Spider Phobia Questionnaire, SPQ; Hamm, 2006). Individuals were excluded if they were (a) undergoing psychotherapeutic or (b) psychopharmacological treatment or (c) reported elevated psychological distress symptoms (determined by exceeding cutoff values of >10 for depressive and stress symptoms and >6 for anxiety symptoms on the Depression, Anxiety and Stress Scales-21, DASS-21; Nilges & Essau, 2015). 260 individuals were screened for eligibility. 56 did not meet the spider-fearful/non-fearful criterion, 45 quit the screening, 29 reported elevated distress symptoms, and 10 were currently undergoing psychological or psychopharmacological treatment. Thus, our final sample consisted of 120 individuals (55 spider-fearful). They were reimbursed by course credit or participation in a raffle for one of 10 gift vouchers worth 30€ each.
Materials
Self-Report Measures
Depression-Anxiety-Stress-Scale-21
The DASS-21 is a 21-item scale measuring depression, anxiety and stress symptoms during a one-week period (German version by Nilges & Essau, 2015; original by Lovibond & Lovibond, 1995). Items are rated on a 4-point Likert scale ranging from 0 = ‘Does not apply to me at all’ to 3 = ‘Applies strongly to me or most of the time’. The DASS showed good sensitivity and specificity to detect clinically relevant symptoms in a German sample (Nilges & Essau, 2015). Proposed cutoff values are 10 for depressive, 6 for anxiety and 10 for stress symptoms. The overall scale showed good internal consistency in our sample (α = .82).
Spider Phobia Questionnaire
The SPQ is a 31-item scale measuring spider phobia symptom severity via a dichotomous response format (‘Yes’ vs. ‘No’; German version by Hamm, 2006; original by Klorman et al., 1974). It is widely used to measure spider phobia severity and has been shown to be reliable and valid (Hamm, 2006). The internal consistency in our sample was excellent (α = .94).
Fear of Spiders Questionnaire
FSQ is an 18-item scale also measuring spider phobia symptom severity (German version by Rinck et al., 2002; original by Szymanski & O’Donohue, 1995). Items are rated on a 7-point Likert scale ranging from 0 = ‘not at all’ to 6 = ‘very much’. It is frequently used to corroborate findings from the SPQ (e.g. Haberkamp et al., 2019) and has shown good psychometric qualities (Rinck et al., 2002). The internal consistency in our sample was excellent (α = .97).
State-Trait-Anxiety-Inventory–State
The STAI-S is a widely used 20-item scale measuring state anxiety (German version by Laux et al., 1981; original by Spielberger, 1970). Responses are given on a 4-point Likert scale ranging from 0 = ‘not at all’ to 3 = ‘very much’. Psychometric qualities of the scale are good (Laux et al., 1981) and the internal consistency in our sample was excellent (α = .93).
Intolerance of Uncertainty Scale Short Form
IUS-12 measures trait IU via 12 items responded to on a 5-point Likert scale ranging from 0 = ‘not at all characteristic of me’ to 4 = ‘entirely characteristic of me’ (German version by Dietmaier et al., 2008; original by Carleton et al., 2007). Although it contains two subscales, we were interested in the overall scale. Its psychometrics have been shown to be adequate (Dietmaier et al., 2008) and the internal consistency in our sample was good (α = .83).
3D-Animated Videos
We created novel 3D-animated videos of spiders and ladybugs, as non-aversive control stimuli. This allowed us to keep the movement characteristics and environment invariant, in contrast to real-life animal videos where animal behaviour and environmental contexts might be highly heterogeneous. Animations were created and rendered using the open-source software Blender (Community, 2018). The animals entered the screen from four different entry points (top left, top right, bottom left and bottom right) and followed a straight or one of two curvy paths. Animals moved on the straight or curvy paths with varying speed, that is, slow, medium or fast. The variation in movement speed was created by altering the playback speed of the videos (i.e. the videos varied in how long their overall playback time was). Animals walking on a curvy path by necessity had to cover a longer distance than animals walking on a straight path, resulting in overall longer videos in the curvy compared to the straight path condition. In the curvy path condition, the video durations were 14 seconds for the slow speed, 11.5 seconds for the medium speed and 9 seconds for the fast speed. In the straight path condition, the durations were 12 seconds for the slow speed, 9.5 seconds for the medium speed and 7 seconds for the fast speed. Initially, the speeds were chosen with the overall duration of the experiment in mind. When first piloting the stimuli, participants in this pilot indicated that they perceived the animals as moving in three distinct speeds. Those variations resulted in 12 different conditions (2 animals × 2 paths × 3 speeds). Each condition was presented twice using different entry points, resulting in 24 trials per participant. The frequency of each entry point was equal in the straight and curvy path conditions. All videos are accessible on OSF (https://osf.io/zfj78/).
Procedure
The experiment was conducted online using SoSci Survey (Leiner, 2021). First, individuals provided informed consent and entered demographic data. Then, they indicated whether they currently receive psychological and/or psychopharmacological treatment and filled out the DASS-21 as well as the SPQ. When meeting the inclusion criteria, they also filled out the FSQ, the STAI-S and the IUS-12.
Following that, participants were instructed to rate all 24 videos with respect to their emotional response (fear, disgust, arousal and valence) and perceived uncontrollability and unpredictability of the animal movements. We also informed participants that they should avoid looking away from the screen during video presentation. Ratings were obtained immediately after each video. Participants were asked to ‘Please rate the video you have just seen in terms of the following characteristics’. They responded on separate 7-point Likert scales ranging from 0 to 6. The characteristics represented our outcome measures and were ‘Fear-Inducing’ (0 = ‘Not at all’, 6 = ‘Extremely) for fear, ‘Disgusting’ (0 = ‘Not at all’, 6 = ‘Extremely) for disgust, ‘Arousing’ (0 = ‘Not at all’, 6 = ‘Extremely) for physiological arousal, ‘The video was…’ (0 = ‘Very pleasant’, 6 = ‘Very unpleasant’) for valence and ‘The movements appeared to be…’ (0 = ‘very controllable’/‘very predictable’, 6 = ‘very uncontrollable’/‘very unpredictable’) for perceived uncontrollability and perceived unpredictability of the movements, respectively. Note that the response scales for perceived uncontrollability and unpredictability were intended to capture fear-related aspects as proposed in the cognitive vulnerability model by Armfield (2006). Thus, the scale for perceived unpredictability was intended to measure the extent to which participants thought they were able to predict the movements of the animal. The scale for perceived uncontrollability was intended to measure the perceived ‘inability to exert influence over the movement, approach, or behavior of an animal’ (Armfield, 2006, p. 756).
During video presentation, the experiment was displayed in fullscreen mode and the cursor was removed. Video presentation was fully randomized with the constraint that no animal was shown more than twice in a row. After completing the rating of the last video, participants received either course credit or participated in the raffle.
Statistical Analyses
All data were preprocessed and analyzed using R version 4.05 (R Core Team, 2021). Deviations from the preregistered analysis plan are explained on OSF (https://osf.io/5zwbk/). We calculated linear mixed effects models (LMMs) to analyze fear, disgust, arousal and valence as well as our main outcome measures (uncontrollability and unpredictability) using the mixed function of the afex package (Singmann et al., 2021). All factors of our 2 (spider fear: spider-fearful vs. non-fearful) × 2 (animal: spider vs. ladybug) × 2 (movement path: straight vs. curvy) × 3 (speed: slow vs. medium vs. fast) design were entered as fixed effects into each model. The design thus consisted of one between-subjects factor (spider-fear), and three within-subject factors (movement path, movement speed and animal). We will not report four-way interactions. STAI scores of each participant were included as covariates to control for baseline anxiety, as well as the repeated measurement factor as each condition was presented twice. Intercepts were allowed to vary between individuals as a random effect to account for the non-independence of our within-subject factors. Interaction contrasts were analyzed using the emmeans package (Lenth, 2021). We used restricted maximum likelihood estimation (REML) and an unstructured covariance matrix to estimate parameters. We report unstandardized coefficients for all models. Differences between fearful and non-fearful participants in questionnaire scores (i.e. SPQ, FSQ, STAI, DASS-21 and IUS-12) and demographics were analyzed via t-tests. We analyzed the relationship between IUS-12 scores and outcome measures via partial correlations, to assess the specificity of the influence of IU.
Results
Sample Characteristics
Spider-fearful and non-fearful group sample characteristics.

Note. DASS – D/A/S = Depression-Anxiety-Stress-Scales–Depression/Anxiety/Stress, SPQ = Spider Phobia Questionnaire, FSQ = Fear of Spiders Questionnaire, STAI = State-Trait-Anxiety-Inventory – State, IUS-12 = Intolerance of Uncertainty Scale 12. Parentheses contain standard deviations. Bold t-values denote results with p < .05.
Emotional Ratings of Phobic Stimuli
If our videos of 3D-animated spiders are sufficiently realistic, they should elicit elevated emotional responses in spider-fearful individuals. Thus, analyses of the main effects of group and animal serve as a general manipulation check. Indeed, we found a main effect of group on all four emotional ratings (spider-fearful vs. non-fearful: all Bs > 0.49, ps < .001), that is, spider-fearful participants reported higher fear, disgust, arousal and more negative valence ratings compared to non-fearful participants. We also observed a main effect of animal on all four emotional ratings (spider vs. ladybug: all Bs > 0.70, ps < .001), that is, spiders elicited more negative emotions compared to ladybugs. In addition, the group × animal interaction was significant (all Bs > 0.53, p < .001), that is, the negative emotions elicited by spiders were more pronounced in the spider-fearful compared to the non-fearful group (for detailed results, see Supplementary Table S1 in the Supplemental Material available online).
Influence of Movement Path on Emotional Ratings
We predicted that the movement path of the animated spiders would affect the emotional ratings of spider-fearful individuals; specifically, spiders moving on a curvy path should elicit more negative emotions in spider-fearful individuals because these movements might appear more uncontrollable and unpredictable. Indeed, we found a main effect of path on fear, disgust, arousal and valence ratings (all Bs > 0.07, ps < .001) and significant two-way interaction effects of group × path on all emotional ratings (all Bs > 0.04, ps < .035; for detailed results, see Supplementary Table S1 in the Supplemental Material available online), indicating that the effect of path on emotional ratings differed between spider-fearful and non-fearful participants. Also, we observed significant two-way interaction effects of animal × path on all emotional ratings (all Bs > 0.04, ps < .015, see S1), indicating that the effect of path also differed depending on animal type.
For arousal and valence, we did not find any significant three-way interaction effects. Their significant two-way interaction effects were explained by higher arousal and valence ratings for curvy paths in spider-fearful (Bs > 0.34, ps < .001) compared to non-fearful individuals (Bs < 0.22, ps < .001), and higher ratings when observing spiders moving on a curvy path (Bs > 0.35, ps < .001) compared to ladybugs (Bs < 0.22, ps < .001). See Supplementary Table S2 in the Supplemental Material available online for details of the two-way interactions.
For fear and disgust, we found significant three-way interaction effects of group × animal × path (B = 0.07 for fear and B = 0.03 for disgust, both ps < .034); differences in fear and disgust ratings between spider-fearful and non-fearful individuals did not only depend on the observed animal but also on the path it was moving on. Specifically, spider-fearful individuals reported higher fear and disgust when observing spiders moving on a curvy compared to a straight path (both Bs > 0.42, ps < .001). This effect was not present in non-fearful individuals (|Bs| < 0.09, ps > .145). Also, movement path did not affect emotional ratings when observing ladybugs, both in spider-fearful (Bs < 0.07, p > .530) and non-fearful individuals (Bs < 0.02, p > .802; for detailed results see Supplementary Table S3 in the Supplemental Material available online, for an illustration, see Figure 1). This suggests that spider-fearful individuals experience more fear and disgust when spiders, but not other non-phobic small animals, move in an apparently more uncontrollable and unpredictable manner. Differences in mean fear and disgust ratings between the curvy and straight path condition, separated by animal (spider vs. ladybug) and group (spider-fearful vs. non-fearful individuals). Note. Error bars represent 95% CIs. *** = p < .001, n.s. = not significant.
Influence of Movement Speed on Emotional Ratings
We predicted that speed would influence emotional ratings of spider-fearful participants with more negative responses to faster in contrast to slower moving spiders, to ladybugs and ratings of non-fearful participants. In contrast to this hypothesis, we found no effect of movement speed on emotional ratings. Rather, we observed a main effect of speed on arousal ratings (B = 0.10, SE = 0.03, CI [0.04; 0.16], t (2728.2) = 3.39, p < .001), with increasing arousal with faster movements. All other analyses were non-significant (cf. Supplementary Table S1 in the Supplemental Material available online).
Perceived Uncontrollability and Unpredictability of Animal Movements
We predicted that the movement path of animated spiders would affect perceived uncontrollability and unpredictability of animal movements in spider-fearful participants. More specifically, we expected that spiders and ladybugs moving on a curvy rather than a straight path would be perceived as more uncontrollable and unpredictable. First, we observed significant main effects for group on both perceived uncontrollability (unc) and unpredictability (unp) (unc: B = 0.34, SE = 0.1, CI [0.14; 0.54], t (117) = 3.55, p < .001; unp: B = 0.28, SE = 0.09, CI [0.11; 0.45], t (117) = 3.16, p = .002), with spider-fearful individuals reporting higher uncontrollability and unpredictability compared to non-fearful individuals. We also found a main effect of animal (unc: B = 0.4, SE = 0.02, CI [0.35; 0.45], t (2728.1) = 16.57, p < .001; unp: B = 0.33, SE = 0.02, CI [0.28; 0.38], t (2728.1) = 13.37, p < .001), showing that spiders were rated as moving more uncontrollably and unpredictably compared to ladybugs. Also, main effects of path (unc: B = 1.03, SE = 0.02, CI [0.98; 1.08], t (2728.1) = 42.83, p < .001; unp: B = 1.34, SE = 0.02, CI [1.29; 1.39], t (2728.1) = 54.36, p < .001) and speed (unc: B = 0.17, SE = 0.04, CI [0.09; 0.24], t (2728.1) = 4.07, p < .001; unp: B = 0.09, SE = 0.04, CI [0.01; 0.17], t (2728.2) = 2.09, p = .037) demonstrated that animals were rated as more uncontrollable and unpredictable when moving on a curvy path or when moving faster, compared to a straight path or when moving slower.
We also found significant two-way interactions of group × animal (unc: B = 0.25, SE = 0.02, CI [0.21; 0.30], t (2728.1) = 10.52, p < .001; unp: B = 0.18, SE = 0.02, CI [0.14; 0.24], t (2728.1) = 7.46, p < .001) and group × path (unc: B = 0.13, SE = 0.02, CI [0.08; 0.17], t (2728.1) = 5.22, p < .001; unp: B = 0.13, SE = 0.02, CI [0.08; 0.18], t (2728.1) = 5.14, p < .001), plus significant three-way interactions of group × animal × path (unc: B = −0.06, SE = 0.02, CI [-0.11; −0.01], t (2728.1) = −2.51, p = .012; unp: B = −0.05, SE = 0.02, CI [-0.10; −0.01], t (2728.1) = −2.15, p = .032). Thus, the effect of the movement path on uncontrollability and unpredictability ratings varied between spider-fearful and non-fearful individuals and also depended on the observed animal.
This significant interaction revealed a specific pattern of responses (see Figure 2 for an illustration). Again, spider-fearful individuals perceived spider movement as more uncontrollable and unpredictable compared to ladybugs and non-fearful individuals. However, these evaluations of spiders as moving uncontrollably and unpredictably were, to a lesser degree, also present when spiders moved on a straight path, resulting in a smaller effect of path when observing spiders (unc: B = 2.15, SE = 0.1, CI [1.96; 2.35], t (2728) = 21.52, p < .001; unp: B = 2.78, SE = 0.1, CI [2.58; 2.98], t (2728) = 27.11, p < .001) compared to ladybugs (unc: B = 2.47, SE = 0.1, CI [2.27; 2.67], t (2728) = 24.69, p < .001; unp: B = 3.09, SE = 0.1, CI [2.89; 3.29], t (2728) = 30.1, p < .001). In contrast, in non-fearful individuals, the effect of movement path was similar when observing spiders (unc: B = 1.89, SE = 0.09, CI [1.71; 2.07], t (2728) = 20.58, p < .001 unp: B = 2.49, SE = 0.09, CI [2.3; 2.67], t (2728) = 26.36, p < .001) and ladybugs (unc: B = 1.73, SE = 0.09, CI [1.55; 1.91], t (2728.3) = 18.68, p < .001 unp: B = 2.37, SE = 0.09, CI [2.18; 2.56], t (2728.4) = 25, p < .001). This suggests a potential perception bias in spider-fearful individuals which leads to elevated perceived uncontrollability and unpredictability of spider movement compared to ladybugs, even when spiders moved along a straight, arguably more controllable and predictable path. Differences in mean uncontrollability and unpredictability ratings between the curvy and straight path condition, separated by animal (spider vs. ladybug) and group (spider-fearful vs. non-fearful individuals). Note. Error bars represent 95% CIs. *** = p < .001.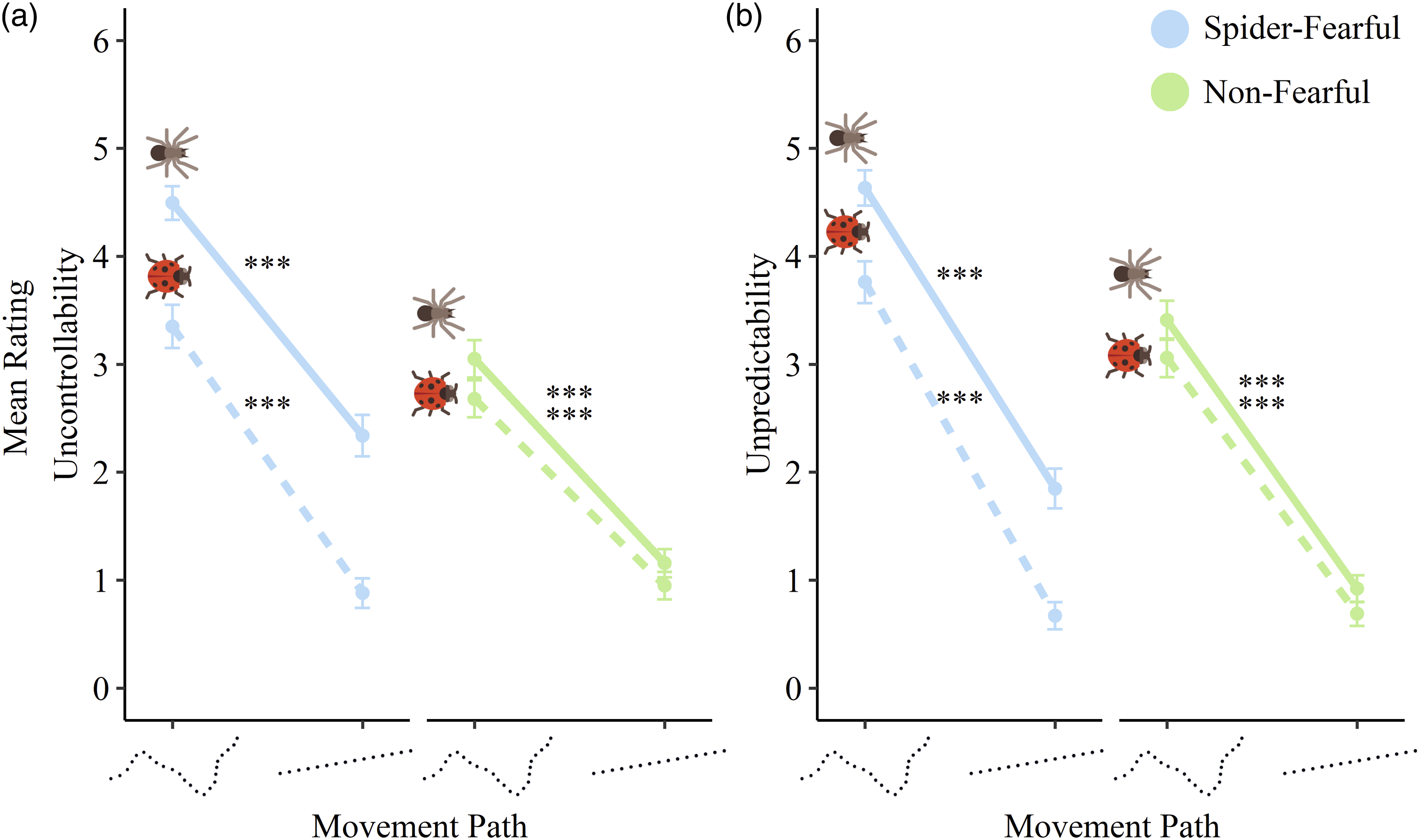
IU in Spider-Fearful and Non-Fearful Individuals and Associations of IU with Emotional Ratings
Partial Correlations between emotional, uncontrollability, unpredictability and intolerance of uncertainty scores of participants, controlling for spider fear measured via SPQ.
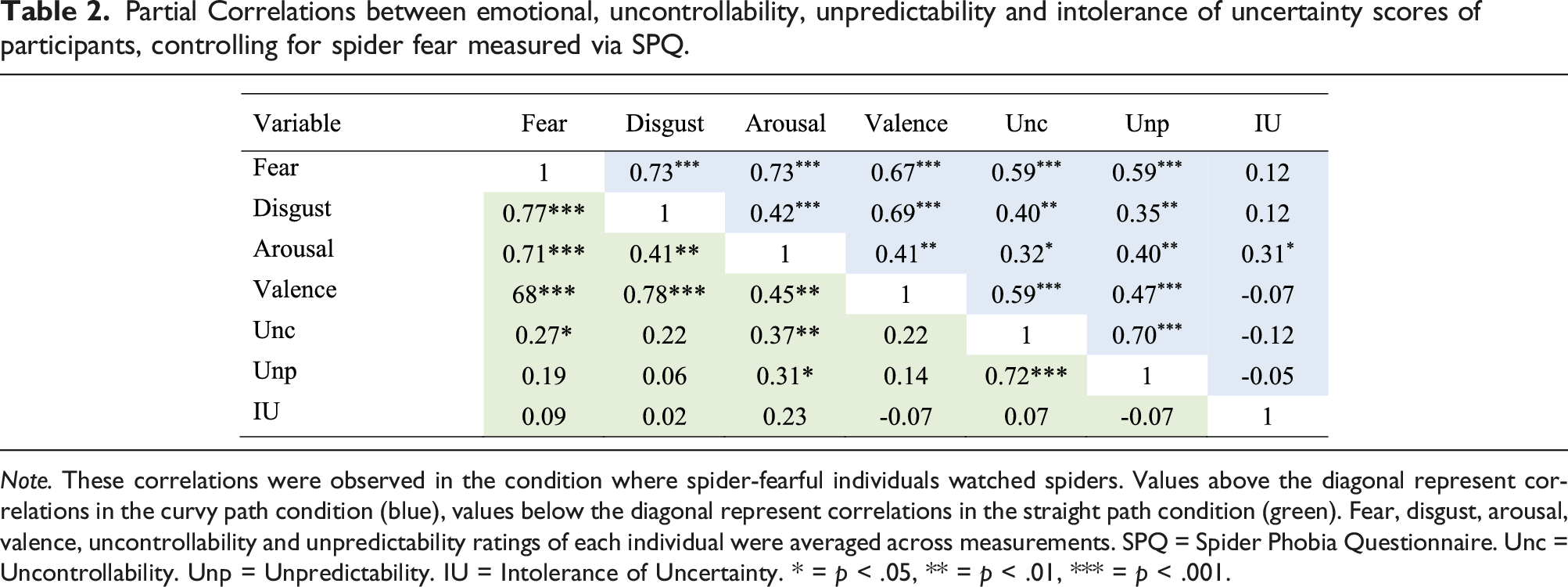

Note. These correlations were observed in the condition where spider-fearful individuals watched spiders. Values above the diagonal represent correlations in the curvy path condition (blue), values below the diagonal represent correlations in the straight path condition (green). Fear, disgust, arousal, valence, uncontrollability and unpredictability ratings of each individual were averaged across measurements. SPQ = Spider Phobia Questionnaire. Unc = Uncontrollability. Unp = Unpredictability. IU = Intolerance of Uncertainty. * = p < .05, ** = p < .01, *** = p < .001.

Partial correlation between arousal rating and IU for spider-fearful individuals observing spiders moving on a curvy path, controlled for spider fear measured via SPQ. Note. The x- and y-axis display the residuals of the measures after controlling for the influence of SPQ. The grey area displays CIs. SPQ = Spider Phobia Questionnaire. IU = Intolerance of Uncertainty.
Discussion
The present study investigated the effects of spider movement characteristics (i.e. movement path and speed) on emotional reactions and perceived uncontrollability and unpredictability in spider-fearful and non-fearful individuals. Also, we tested the relationship between these variables and the transdiagnostic risk factor intolerance of uncertainty (IU). Spider-fearful individuals rated spiders moving on a curvy path as more fear inducing and disgusting than when they moved on a straight path, which was not the case for non-fearful individuals and also not when a ladybug was observed. Moreover, spider-fearful individuals perceived the spiders as moving uncontrollably and unpredictably, (1) especially when they moved on a curvy path, (2) to a lesser degree also when they moved on a straight path and (3) more than ladybugs and non-fearful individuals. Lastly, spider-fearful individuals displayed elevated levels of IU compared to non-fearful individuals, and IU levels were positively correlated to the arousal ratings of spider-fearful individuals when they observed spiders moving on a curvy (but not on a straight) path, even after controlling for levels of spider fear.
We used realistic 3D-animated animal videos which were specifically created for the purpose of our study and allowed us to combine ecological validity with highly controlled experimental conditions. For example, we designed the different movement path conditions to maximize perceived randomness of the animal movements in one condition, and minimize it in the other. To our knowledge, our study is the first that explicitly measured the influence of spider movements on emotional reactions and perceived uncontrollability and unpredictability, using realistic 3D-animated videos in an experimental design.
By definition, spider phobia is described as increased negative emotional reactions (i.e. fear, disgust, arousal and valence) to spiders (APA, 2013). These reactions have been well-documented (Cisler et al., 2009; Matchett & Davey, 1991; Tolin et al., 1997) and do also occur in our study. Spider-fearful individuals reported more negative emotional reactions to spiders compared to ladybugs and to non-fearful individuals. However, we also observed specific effects of spider movement paths on emotional ratings in spider-fearful individuals. In line with our hypothesis, spider-fearful individuals rated spiders as more fear-inducing and disgusting when moving on a curvy compared to a straight path. The curvy movement path also elicited higher arousal and more negative valence ratings compared to the straight path, even though this effect was also observed in non-fearful individuals and when observing ladybugs, albeit weaker (i.e. ratings in non-fearful individuals were lower, and ladybugs were rated lower in arousal and negative valence). This is in line with previous studies reporting higher physiological arousal in response to a fear-irrelevant stimulus, for example, in non-acrophobic individuals confronted with heights in virtual reality (Diemer et al., 2016), potentially representing a general response to situations calling for caution.
In contrast to our expectations, speed only affected arousal ratings and was limited to higher arousal ratings with higher speed, irrespective of spider fear or observed animal. In other words, the effect we found for speed in our study seemed unrelated to any fear-relevant mechanisms. This was surprising, since ‘fast’ was one of the most frequent words that spider-phobic individuals associated with fear-inducing aspects of spiders in the study by Lindner et al. (2019). One potential explanation for this finding might be that the fear-relevant aspects of spider movement speed are triggered differently in real-life encounters. Specifically, spiders in real-life encounters tend to either sit still or, when disturbed, dart away in a sudden fashion, whereas the spiders in our stimuli moved with consistent speeds. The inconsistencies of our study results with those of Lindner et al. (2019) highlight that further research is needed to clarify the specific role that movement speed plays in spider fear.
Consistent with our hypotheses, however, animals that moved on a curvy path lead to higher ratings of uncontrollability and unpredictability in both groups. Also, spider-fearful individuals perceived spiders on a curvy path to move in a more uncontrollable and unpredictable manner compared to ladybugs and non-fearful individuals. Notably, this elevated perception of uncontrollability and unpredictability was also observed when spiders moved in a straight line (albeit to a lesser degree compared to them moving on a curvy path). This unexpected finding suggests a potential bias in the perception of spider movements, which may to a certain degree be irrespective of actual movement path.
Within the cognitive vulnerability model of fear (Armfield, 2006), perceived unpredictability and uncontrollability are crucial vulnerability schemas in individuals with specific fears. In spider fear, this should result in internal representations of spiders as highly uncontrollable and unpredictable, which is supported by our findings. This biased perception of spider behaviour (i.e. their movements) might be inherently aversive for spider-fearful individuals and increase the subjective likelihood of negative outcomes when encountering a spider, with respect to dangerousness (i.e. being attacked by the spider; Armfield, 2006), disgust (i.e. coming into contact with the spider; Knowles et al., 2019) or both. This is in line with findings that spider-fearful individuals generally overestimate the probability of negative outcomes when confronted with a spider (Hengen & Alpers, 2019; Van Overveld et al., 2006). These vulnerability schemas with respect to spider movements may also be partially resistant to change, even in light of new evidence (i.e. the spider being harmless) and may therefore play a role in maintaining the fear. Such a partial resistance to change is also supported by our observation of biased perceptions in a ‘virtual’ situation where spiders cannot pose a real threat to participants. Relatedly, a growing body of research suggests that individuals suffering from anxiety disorders (e.g. social anxiety, panic disorder, etc.) fail to change their fear-related beliefs even in light of contradicting evidence (Kube & Rozenkrantz, 2021). In spider fear, this may prevent spider-fearful individuals from learning that spider behaviour is not as uncontrollable and unpredictable as their schemas suggest, thereby limiting safety learning (Duits et al., 2015).
As expected, spider-fearful individuals displayed higher IU levels compared to non-fearful individuals, suggesting elevated IU levels in the spider-fearful compared to the general population. However, previous studies in patients with other anxiety disorders (e.g. social anxiety, generalized anxiety disorder and obsessive compulsive disorder) found IU levels markedly above those of our spider-fearful participants (Carleton et al., 2012). Consequently, IU may play a less important role in spider fear compared to other anxiety disorders. This notion is supported further by our finding that after controlling for spider fear, IU was only positively correlated to arousal ratings in spider-fearful individuals watching spiders move on a curvy path (but not on a straight path). However, it has also been shown previously that fear responses to fear relevant-animals (even when presented in virtuo) are especially pronounced when both perceptual (i.e. an actually visible feared animal) and contextual (i.e. the knowledge that in the current environment, a feared animal might be present/appear) cues are present (Peperkorn et al., 2013). It seems possible that any perceived uncertainty directly related to IU could have been tied to the question whether a spider or a ladybug might appear on the grass plane. In our study, the chance of a spider appearing was 50%, as the amount of spider and ladybug videos was equal. In other words, our participants never knew which animal to expect. The display of the grass plane could have served as an ambiguous cue signaling a threatening (spider) or neutral (ladybug) animal, thereby increasing the perceived uncertainty. Relatedly, a study by Abado et al. (2020) manipulated the expectancies of spider-fearful individuals to decrease the uncertainty about whether a threatening (spider) or neutral (bird) animal would appear. The authors found that such a decrease in uncertainty led to a decrease in attention bias towards spiders and concluded that an increase in certainty regarding the appearance of a threatening stimulus may be helpful in reducing fear. This study further supports the notion that expectancies play a role in spider fear. Taken together, these notions support that the interactions of expectancies, IU and emotional responses to threatening stimuli should be further investigated in future studies.
With the uncertainty of which animal would appear being removed when a spider did actually appear (i.e. the dreaded negative outcome came to pass), IU may not have had an influence above and beyond the triggered fear response, resulting in absent correlations. Moreover, IU has predominantly been shown to influence physiological arousal in conditioning experiments, with evidence suggesting that individuals high in IU experience heightened arousal (particularly skin conductance) when information about threat contingencies is absent and the unconditioned stimulus does not occur (Morriss et al., 2021). This ties well into our research, where, as mentioned, participants did also not know about the threat contingencies. In sum, this may explain the heightened arousal in those spider-fearful individuals which reported comparably higher IU levels and the lack of associations with other fear-relevant outcomes in our study.
Surprisingly, we did not find a correlation between IU and uncontrollability or unpredictability. This is in contrast to our assumption that the interpretation of situations (here, spider encounters) as uncontrollable and unpredictable would be directly affected by IU. We speculate that IU might not increase the perceived uncontrollability and unpredictability of the spider movements, but instead might render these perceptions more aversive. In other words, the degree to which spider-fearful individuals interpret spider movement as uncontrollable and unpredictable might depend more on disorder-specific beliefs about spider behaviour, rather than IU. In turn, the resulting increase in perceived uncontrollability and unpredictability would make encounters with spiders less tolerable for spider-fearful individuals, specifically for those with high levels of IU (defined as an “inability to tolerate the aversive reaction triggered by a perceived lack of sufficient or salient information”; Carleton, 2016, p. 31). Along these lines, Shihata et al. (2017) conducted a study investigating disorder-specific and context related components of IU in their study and found that these components explained variance above and beyond trait IU. Similarly, as spider fear is defined by a narrow area in which IU might play a role (i.e. only in situations involving spiders), the IUS-12 used in this study may not have adequately captured these facets of IU tied to the fear-relevant situations of our sample. Moreover, it is possible that the participants’ responses on the scales for perceived uncontrollability and, potentially to a lesser extent, unpredictability were influenced by the fact that we used videos as stimuli. More specifically, participants never had any actual control over the animals’ movements because no real interaction with them was possible (as opposed to real-life encounters, where it is possible to e.g. put a jar over a spider). Therefore, participants had to infer how much control they would have over the animals’ movement in hypothetical real-life scenarios to be able to rate the scales according to our theoretical considerations. In our study, we had no way to ascertain whether, if at all, our participants imagined such hypothetical scenarios and to what extent they informed their uncontrollability ratings. It seems plausible that such mental processes could have influenced the ratings depending on how the participants interpreted the stimuli and the perceived uncontrollability rating scale. For instance, some participants may have created imaginary scenarios of real-life spider encounters mimicking the situation presented in our videos to varying degrees. These scenarios would likely be influenced by participants’ previous experiences with spider encounters and could have plausibly influenced their ratings of perceived uncontrollability depending on the imagined scenario. Such processes might have obscured any correlations of uncontrollability (and possibly also unpredictability) ratings with IU due to the cognitive elaborations necessary to give the ratings, whereas IU can be seen as a construct more directly related to the emotional reactions triggered by situations perceived as uncertain in terms of a potentially negative outcome (Carleton, 2016). Future studies using virtual stimuli could therefore attempt to give participants actual control over the spider’s movements by, for example, providing them with the ability to move a jar across the screen with which they may trap the spider. Alternatively, one could also provide clear instructions to participants that they should base their ratings of uncontrollability/unpredictability on pre-specified hypothetical real-life scenarios.
Limitations
First, we collected our experimental data online. Although the accuracy of computerized and internet based self-reports has been demonstrated (Gwaltney et al., 2008; Weigold et al., 2013), confounding factors may still have influenced the experimental procedure. For example, we do not know whether participants performed the experiments alone or were distracted, or which screen they used to watch the videos. Although we believe that participants in our study correctly understood the intended meaning of our response scales, we acknowledge that we cannot exclude that some participants may have misunderstood their meaning due to no experimenter being present to respond to questions. Additionally, the online format of our experiment prohibited us from collecting potentially insightful measurements aside from self-report data, such as physiological (e.g. SCR) or eye-tracking measures. This seems especially important to draw a complete picture of the influences of IU on emotional reactions in spider fear, since spider fear and IU have been shown to differently influence attentional processes measured via eye-tracking (Spider Fear: McSorley et al., 2017; IU: Morriss & McSorley, 2019) and physiological responding (IU: Morriss, 2019; Spider Fear: Peperkorn et al., 2013). Second, our sample consisted of mostly female participants, limiting generalizability to male individuals suffering from spider fear. Third, our study is cross-sectional and therefore does not allow to draw causal inferences, for example, about whether elevated IU levels in spider-fearful individuals precipitated their spider fear. Fourth, our findings with 3D-animated videos might not entirely generalize to real-world encounters with spiders (specifically regarding movement speed). While we took an effort to create realistic displays of moving spiders, video presentations still differ in crucial aspects from real encounters, for example, by making physical contact with the spider impossible. Therefore, we would expect our findings to be even more pronounced when testing real-world situations. Finally, using a ladybug as a control stimulus could also be viewed as a limitation because ladybugs differ on several levels from spiders (e.g. number of legs, size, visibility of legs, movement speed in real life, etc.). However, we chose ladybugs as control stimuli since these animals have a neutral (or positive) valence and are at least somewhat comparable in size compared to other arachnids (such as scorpions, mites, ticks, etc.) – although we do acknowledge that many spider species are larger than ladybugs. One other possible alternative might be to use certain ant species as a control stimulus, although ant species large enough to be at least somewhat comparable to spiders in size are potentially not neutral in valence.
Conclusion
In conclusion, our study provides, as far as we are aware, first experimental evidence that spider-fearful individuals perceive the movement of spiders as disproportionately uncontrollable and unpredictable. Furthermore, this higher perceived uncontrollability and unpredictability is associated with aversive emotional reactions. This belief of spiders behaving erratically seems to be, at least to a certain degree, independent of the spider’s actual behaviour, although additional studies are needed to clarify this. Spider-fearful individuals also reported markedly higher IU levels than non-fearful individuals, which were also correlated to arousal ratings when observing spiders. Our findings highlight the importance of using moving stimuli to investigate spider phobia to capture real-world behaviour as accurately as possible. Specifically, our results support the notion that biased perceptions of uncontrollability and unpredictability of spider movements are important cognitive factors in spider fear. In contrast, using static spider stimuli would not allow to observe these biases, and also increase certainty about spider behaviour (especially in experimental lab situations) so that other factors such as IU might not take effect. Finally, our findings also have implications for clinical practice. Specifically, we argue that an important component in exposure therapy is that spider-fearful individuals learn that they have more control over spider movements (learning e.g. that they can react before a spider crawls underneath their clothes) and that movements are more predictable than assumed. This learning might be instrumental in changing the belief that a spider might suddenly attack, jump or make physical contact, with nothing that could be done about it. Consequently, a reduction in perceived uncontrollability and unpredictability after exposure treatment could be regarded as a relevant treatment outcome measure.
Supplemental Material
Supplemental Material - Out of control: The role of spider movement and intolerance of uncertainty in spider fear
Supplemental Material for The role of spider movement and intolerance of uncertainty in spider fear by Markus Grill and Anke Haberkamp in Journal of Experimental Psychopathology
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Deutsche Forschungsgemeinschaft (DFG; German Research Foundation) [grant number 450096725] awarded to AH.
Supplemental Material
Supplemental material for this article is available online.
Author Biographies
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.