
Abstract
Intolerance of uncertainty (IU) is a transdiagnostic risk factor for psychiatric disorders, and plays an important role in fear and threat learning under uncertainty. The ‘reinstatement of fear/threat’ is an understudied phenomenon thought to represent clinical symptom relapse. Reinstatement of conditioned responding can be captured in the laboratory by presenting unsignaled presentations of an aversive unconditional stimulus. The present study investigated IU as a predictor of reinstatement effects, such that individuals higher in IU, relative to lower IU, would show larger reinstatement. Sixty-two participants completed the Intolerance of Uncertainty Scale and provided threat expectancy ratings (i.e. certainty of receiving a shock) during a differential threat conditioning and extinction paradigm with reinstatement. Findings suggested a differential increase in threat expectancy ratings to both the threat and safety cue following reinstatement, although this effect was small and did not survive follow-up tests. Nevertheless, IU was a significant predictor of reinstatement to the threat cue but not the safety cue, although this effect was not in the expected direction. Specifically, higher IU was associated with reduced threat expectancy ratings post-reinstatement. These findings provide support for the limited literature demonstrating an important role for IU in reinstatement effects and should be investigated further.
Intolerance of uncertainty (IU) refers to elevated anxiety and arousal to uncertain or ambiguous events and situations and is considered a core mechanism in problematic coping styles (Rosen et al., 2014). Moreover, IU has been well-studied as a transdiagnostic mechanism of a number of psychiatric disorders, including generalized anxiety disorder, obsessive-compulsive disorder (Holaway et al., 2006) and anorexia nervosa (Brown et al., 2017). In accounting for this association, associative learning theories describe a transdiagnostic intermediate phenotype, underpinned by aberrant threat conditioning and impaired extinction learning, which manifests as tendencies towards (i) rapid threat acquisition, (ii) delayed threat extinction in non-threatening contexts and (iii) rapid reinstatement of threat (Briscione et al., 2014; Murray et al., 2016; Zuj & Norrholm, 2019). To that end, laboratory-based paradigms investigating the return of threat, including reinstatement, can be useful in understanding clinical symptom relapse following appropriate therapies. Reinstatement, in particular, is poorly understood and has received relatively little attention in comparison to other aspects of threat learning (Sjouwerman & Lonsdorf, 2020).
During Pavlovian threat conditioning, a neutral stimulus is repeatedly paired with an aversive unconditional stimulus (US; e.g. a mild electric shock). After repeated pairings, the neutral stimulus becomes a conditional stimulus (CS), producing a conditional response (CR) of increased threat detection. This process is considered a theoretical and experimental analogue for the development of intense fear and anxiety reactions to reminders of a traumatic event (Zuj & Norrholm, 2019). However, in the context of repeated exposure to the CS without the US, a process termed extinction typically occurs, whereby a new memory trace is formed to indicate that the CS no longer predicts threat (Bouton, 2002). Extinction learning, however, is not considered to override or erase the original conditioning memory as the conditioned responses can return due to a number of mechanisms (e.g. Bouton & Moody, 2004; Myers & Davis, 2007).
Reinstatement is the process of threat return due to unsignaled encounters with the US following extinction learning. In clinical situations, reinstatement might occur when an individual, who has previously completed a course of prolonged exposure-based therapy, suddenly experiences an unsignaled panic attack whilst in a safe situation, such as shopping. The cognitive and emotional experiences, as well as the physiological sensations, associated with re-experiencing the US can ‘reinstate’ the conditioned emotional reactions to triggers previously associated with the treated psychiatric disorder. In experimental paradigms, reinstatement involves delivery of the US in the absence of the CS following extinction learning, and can result in significant return of threat responding to the original threat cue (Norrholm et al., 2006), as well as generalised (non-differential) return of threat responses to both threatening and safe cues (Dirikx et al., 2009; Kull et al., 2012; Milad et al., 2005; Zuj et al., 2018). More research is needed to understand the effects of individual differences on reinstatement of threat responses to complement recent research investigating experimental boundary conditions of reinstatement (e.g. the number of reinstatement USs, same/different sensory US for reinstatement; Sjouwerman & Lonsdorf, 2020).
Since few learned threat-related associations in real-life settings are universally confirmed, the presence of uncertainty is inherently intertwined with determining whether the US will follow a CS. This uncertainty can be amplified following an unplanned encounter with a US. Reinstatement paradigms therefore introduce significant uncertainty regarding the relationship between the CS+ and US and the overall conditioned context (Haaker et al., 2014; Haaker et al., 2013; Lonsdorf et al., 2014). Further, IU has been positively associated with larger differential conditioned electrodermal responding following a reinstatement manipulation (Dunsmoor et al., 2015; Lucas et al., 2018). Recent research suggests that individuals with high IU display significantly greater skin conductance response magnitude to threat cues compared to safety cues throughout extinction learning (see recent meta-analysis and review, see Morriss, Wake, et al., 2021; Morriss, Zuj, & Mertens, 2021).
The aim of the current study was to investigate the role of IU in the reinstatement of threat expectancy ratings and build on the limited research conducted thus far. Based on the uncertainty regarding the CS-US contingency that may be caused by a reinstatement manipulation, and in line with previous findings using psychophysiological measurements (Dunsmoor et al., 2015; Lucas et al., 2018), we hypothesised that IU would be a significant predictor of reinstated threat expectancy ratings. Specifically, we predicted that elevated IU would be associated with greater threat expectancy ratings following a reinstatement manipulation. Due to past inconsistencies regarding CS-specific versus non-differential reinstatement effects, we made no predictions regarding CS-specific reinstatement.
Method
Participants
Eighty-six participants were recruited from Swansea University and the surrounding community and were aged between 18–57 (M = 26.8 years, SD = 7.6 years), with 37 males and 49 females. Additional demographic information can be found in Supplementary Materials. A sensitivity analysis was conducted using G*Power v3.1 (Faul et al., 2007), showing that for a linear regression assessing R 2 deviation from zero with 1 predictor, Power (1 – β) = 0.80, and ⍺ = .05, the critical F-value is 3.95. Twenty-four participants were excluded 1 due to rating greater threat expectancy of the CS- than the CS+ on the final trial of the threat conditioning phase, suggesting a failure to learn the CS-US contingency, resulting in a total sample size for analyses of N = 62 (25 males and 37 females). A revised sensitivity analysis with the same input parameters as above and a sample size of N = 62 indicates a critical F-value of 4.00.
Exclusion criteria included the presence of a neurological condition (e.g. epilepsy), pregnancy, a history of cardiovascular issues or a pacemaker, or any health issue that may be impacted by the delivery of a mild electric shock. Participants were reimbursed £11.25 for their time; psychology undergraduate participants received research credits in lieu of a monetary reward. The study protocol was approved by the Department of Psychology Research Ethics Committee at Swansea University, and all participants gave full written informed consent.
Materials
Intolerance of Uncertainty Scale (IUS) – short form
The IUS-short form (Carleton et al., 2007) is a 12-item questionnaire assessing intolerance of uncertainty on a 5-point Likert scale ranging from 1 (‘Not at all characteristic of me’) to 5 (‘Entirely characteristic of me’). The 12-item IUS has strong psychometric properties (Khawaja & Yu, 2010).
Depression anxiety stress scale (DASS-21)
The DASS-21 (Lovibond & Lovibond, 1995) is a 21-item questionnaire assessing symptoms of depression, anxiety and stress over the past week on a 4-point Likert scale ranging from 0 (‘Never’) to 3 (‘Almost always’). The DASS-21 provides subscales of depression, anxiety and stress, and each subscale score is multiplied by 2 to be equivalent with the DASS-42 in accordance with scoring instructions. The DASS-21 demonstrates strong reliability and validity in non-clinical UK adults (Henry & Crawford, 2005).
Threat conditioning and extinction paradigm
The US was a 250 ms mild electric shock generated using an STM200 stimulator (BIOPAC Systems, Santa Barbara, USA) and administered through an MLADDF30 bar electrode with two 9 mm contacts spaced 30 mm apart. The CSs were red and blue circles presented on a computer screen and a fixation cross was presented during the inter-trial intervals (ITI). CSs were presented onscreen on a white background for 10 s each, with the ITI ranging from 8 to 12 s (M = 10 s). The CS presentation order was pseudo-random with no more than two consecutive presentations of the same CS. Six seconds into each CS presentation, a visual analogue scale appeared below the CS until a response was made or until the remainder of the trial, and asked participants to ‘Please indicate how certain you are that you will receive a shock after this circle?’ with responses ranging from 0 (‘certain no electric shock’) to 100 (‘certain electric shock’) (e.g. Zuj et al., 2018, 2020). The US occurred immediately following CS offset on relevant trials as used previously by the authors (e.g. Xia et al., 2019; Zuj et al., 2016, 2020) with CS colour counterbalanced.
The experiment had four phases: habituation, threat conditioning, threat extinction and reinstatement. During habituation, participants received two presentations of each of the CS+ and CS- in the absence of the US. During the threat conditioning phase, participants experienced two blocks of eight trials, with four of each CS presented per block. During the first block, the US was delivered pseudo-randomly on 50% of CS+ trials (with the first CS+ trial always being followed by the US) and 100% of CS+ trials during the second block (with a total US reinforcement rate of 75%). This reinforcement rate was adopted from Grady et al. (2016), who found that a partial reinforcement rate followed by a continuous reinforcement rate during conditioning produced extinction-resistant conditioned responses, maximising the sensitivity to identify effects during extinction. The threat extinction phase involved 24 trials (with 12 trials of each CS) and the US was not presented on any trial. Reinstatement stimuli were presented following the final CS of threat extinction. There were three unsignaled shocks during reinstatement that began 12 s after the offset of the final CS of threat extinction and were each separated by 5 s intervals. The reinstatement phase began 30 seconds after the last reinstatement US, and involved 16 trials (eight of each CS). Each experimental phase proceeded continuously from one to the next, with no perceptible change to participants.
Procedure
After providing informed consent, participants gave background demographic information and completed the IUS-short form and the DASS-21. The US electrode was then attached to the palm side of the wrist of the dominant hand and participants underwent shock calibration. The electric current was set to 35 mV as a baseline and increased or decreased in 2.5 mV increments according to the participants’ rating of intensity. Participants were provided with verbal instructions to rate the intensity of each shock on a scale from 1 (‘not at all uncomfortable’) to 5 (‘very uncomfortable’). When a shock was rated as a 4, the participant received a second delivery of this intensity and, if considered ‘uncomfortable but not painful’ by the participant, this shock intensity was used for that participant for the duration of the task (Xia et al., 2019; Zuj et al., 2020). Participants then underwent the conditioning, extinction and reinstatement task described above, after which the participants were debriefed and compensated for their time.
Statistical analyses
Threat expectancy ratings were analysed using a 2 (CS) × k (trial) repeated measures ANOVA for each experimental phase, where k equals the number of trials in each phase. Reinstatement effects were analysed using a 2 (CS) × 2 (time) repeated measures ANOVA comparing the average threat expectancy for the final two trials of the extinction learning phase with the first trial in the reinstatement phase (Haaker et al., 2014; Zuj et al., 2018). Greenhouse-Geisser corrected degrees of freedom and epsilon values (ε) are reported where the assumption of sphericity was violated. Post-hoc comparisons were Sidak-corrected for multiple comparisons. Effect sizes are reported as partial eta-squared (ηp2), and Cohen’s d using the criteria of 0.2, 0.5 and 0.8 for small, medium and large effects, respectively (Cohen, 1988). All models were repeated with DASS depression and stress scores entered as covariates. There were no changes to statistical significance or effect direction, with only minor changes in effect size. As such, the original analyses without covariates are reported (analyses including covariates can be found in Supplemental Materials). Stepwise multiple regressions were conducted to investigate IUS, over and above DASS depression and stress scores, as a predictor of (1) reinstatement of differential responding; (2) reinstatement to the CS+ and (3) reinstatement to the CS-. Reinstatement of differential responding was operationalised as the difference between CS+ and CS- threat expectancy ratings across the final two trials of the extinction phase subtracted from the difference between the CS+ and CS- expectancy ratings for the first trial of the reinstatement phase. Reinstatement to the CS+/− is operationalised as the average of the final two trials of the extinction phase subtracted from the first trial of the reinstatement phase, individually for each respective CS. Regression models were assessed for normal distribution of errors and homoscedasticity of residuals. These analyses were conducted in IBM SPSS v26 for Mac with the alpha set to ⍺ = 0.05.
To compliment Frequentist statistics, Bayesian repeated measures ANOVAs were also conducted using JASP v0.13.1 for Mac (JASP Team, 2020). We evaluated the weight of evidence for the alternative hypothesis over the null (BF10). Here, values greater than 1 represent increasing evidence for the alternative hypothesis, values less than 1 represent increasing evidence for the null hypothesis over the alternative, and values of 1 represent evidence for neither hypothesis (Lee & Wagenmakers, 2013). The BF10 is reported for main effects and the inclusion Bayes Factor across matched models (BFincl) is reported for interactions.
Results
Participant characteristics
Total sample means and standard deviations for demographic measures.

Habituation
The 2 (CS) × 2 (trial) repeated measures ANOVA found no significant main effects or interactions (F’s < 1.33, p’s > .254).
Threat conditioning
The 2 (CS) × 8 (trial) repeated measures ANOVA found, during conditioning, a significant main effect of CS, F (1, 61) = 109.43, p < .001, d = 1.81, BF10 = 1.282e-30, with higher threat expectancy ratings to the CS+ (M = 58.78, 95% CI [53.54, 64.03], SD = 20.66) than the CS− (M = 24.58 [20.25, 28.91], SD = 17.06). Further, there was a significant main effect of trial, F (5.71, 348.47) = 7.99, p < .001, ηp2 = .116, ε = .816, BF10 = 7.518e-113. These main effects were superseded by a significant CS × trial interaction, F (4.78, 291.42) = 35.93, p < .001, ηp2 = .371, ε = .682, BFincl = 4.311e+27. As shown in Figure 1, there is an increasing differential response pattern throughout the conditioning phase, with increasing threat expectancy ratings to the CS+ and decreasing ratings to the CS-. This pattern of effects demonstrates successful acquisition of the CS-US contingency. Trial-by-trial threat expectancy ratings for the CS+ and CS−. HAB: habituation; TC: threat conditioning; EXT: extinction learning; TEST: reinstatement. Yellow lightning symbols indicate unsignaled US presentations. Error bars represent 95% confidence intervals.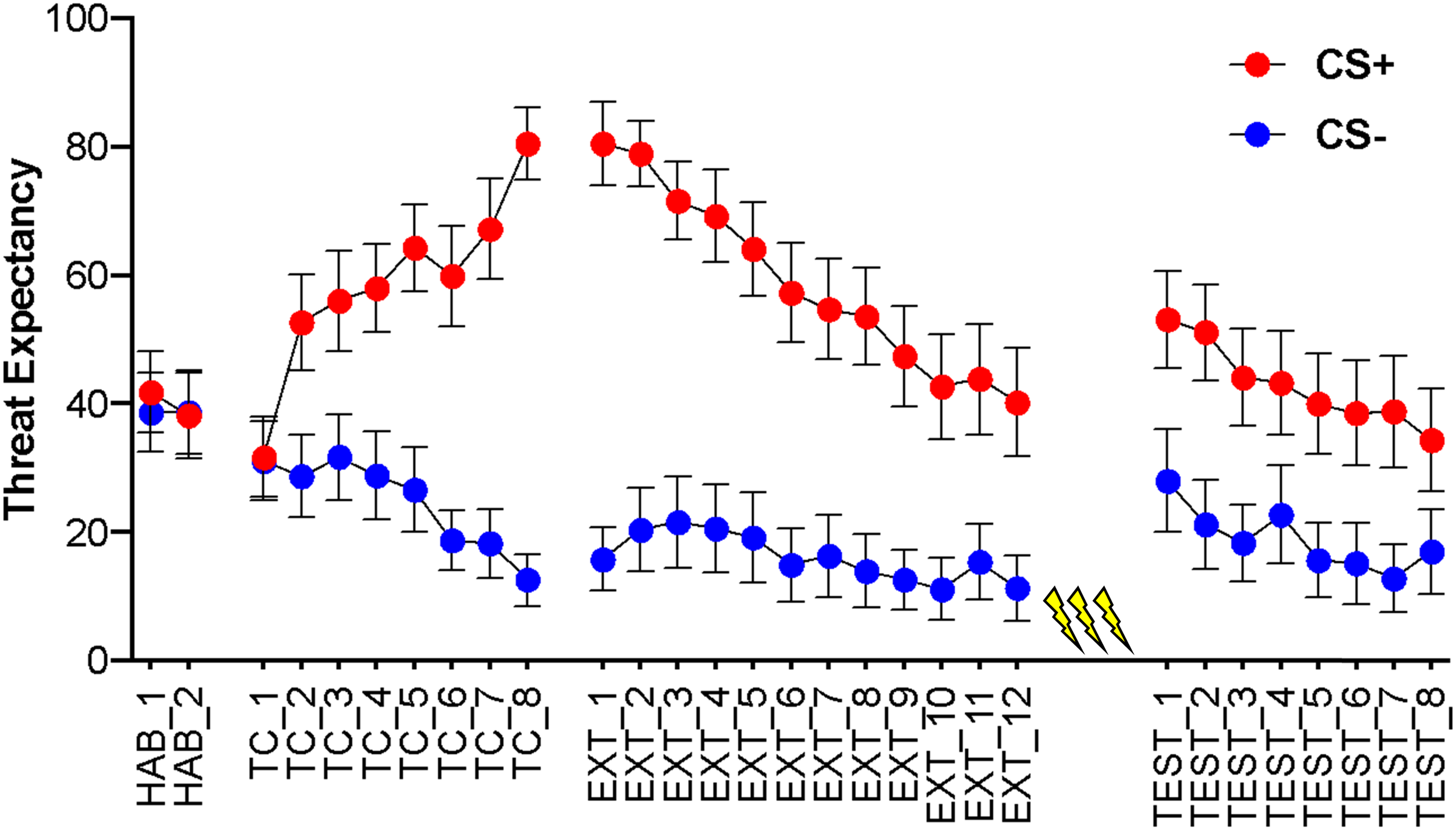
Threat extinction
During the extinction learning phase, the 2 (CS) × 12 (CS) repeated measures ANOVA found a significant main effect of CS, F (1, 61) = 154.02, p < .001, d = 2.02, BF10 = 2.192e-45, with higher threat expectancy ratings for the CS+ (M = 58.69 [52.86, 64.53], SD = 22.97) compared to the CS− (M = 16.10 [11.25, 20.95], SD = 19.10). There was also a significant trial main effect, F (4.40, 268.30) = 27.26, p < .001, ηp2 = .309, ε = .400, BF10 = 8.810e-223. As with the conditioning phase, these main effects were superseded by a significant CS × trial interaction, F (4.39, 267.92) = 18.41, p < .001, ηp2 = .232, ε = .399, BFincl = 1.908e+12. Here, threat expectancy ratings to the CS- remained relatively low whilst CS+ ratings show a significant decline throughout extinction learning (Figure 1). A paired-samples t-test identified significantly higher responding to the CS+ compared to the CS- on the final trial of extinction learning, t (61) = 7.36, p < .001, d = 0.94, BF10 = 1.915e+7, suggesting incomplete extinction.
Reinstatement
After the unsignaled reinstatement stimuli, the 2 (CS) × 8 (trial) repeated measures ANOVA revealed a significant main effect of CS, F (1,61) = 43.71, p < .001, d = 0.98, BF10 = 5.2757, with threat expectancy ratings remaining higher for the CS+ (M = 42.93 [36.08, 49.79], SD = 26.98) than the CS− (M = 18.89 [13.32, 24.45], SD = 21.92). Further, there was a significant main effect of trial, F (3.30, 201.05) = 12.60, p < .001, ηp2 = .171, ε = .471, BF10 = 1.10,157. Here, threat expectancy ratings showed a general downward trend that was relatively uniform across the CS+ and CS− (Figure 1), as indicated by a non-significant CS × trial interaction, F (4.21, 256.69) = 1.95, p = .100, ηp2 = .031, ε = .601, BFincl = 1.90,812.
To assess the change in threat expectancy ratings to the CS+ and CS− as a result of the reinstatement stimuli, a 2 (CS) × 2 (time) repeated measures ANOVA was conducted. As above, there was a significant main effect of CS, F (1, 61) = 48.94, p < .001, d = 1.11, BF10 = 5.0484, with higher threat expectancy ratings to the CS+ (M = 58.55 [49.80, 67.31], SD = 34.49) compared to the CS− (M = 24.53 [17.91, 31.16], SD = 26.09). There was no significant main effect of time, F (1, 61) = 0.19, p = .661, ηp2 = .003, BF10 = 1.33,712. There was, however, a significant CS × time interaction, F (1, 61) = 12.37, p = .001, ηp2 = .169, BFincl = 0.234. Sidak-corrected test of simple main effects showed a greater difference between the CS+ and CS− before reinstatement (M DIFF = 42.94, t (61) = 6.96, p < .001, d = 0.88) compared to post-reinstatement (M DIFF = 25.10, t (61) = 5.34, p < .001, d = 0.68). This suggests that reinstatement introduced an increase in uncertainty regarding the predictive properties of the CS+ compared to the CS−, as reflected by smaller differential threat expectancy ratings in the reinstatement phase. An additional Sidak-corrected test of simple main effects was conducted to assess the difference between pre- and post-reinstatement separately for the CS+ and CS−, however, there was no significant change for either the CS+ (M DIFF = 10.86, t (61) = 1.94, p = .057, d = 0.26) or the CS− (M DIFF = −6.98, t (61) = −1.56, p = .125, d = −0.20) after correcting the familywise error rate.
Intolerance of uncertainty as a predictor of reinstatement effects
Simple linear regressions were conducted to assess intolerance of uncertainty as a predictor of reinstated threat expectancy ratings. IUS scores were a significant predictor of differential threat reinstatement, explaining 11.6% of the variance in differential reinstatement, β = −.340, R
2
= .116, F (1, 60) = 7.86, p = .007, BF10 = 6.37. Further linear regressions indicate that this effect is strongest for the CS+, β = −.338, R
2
= .114, F (1, 60) = 7.73, p = .007, BF10 = 6.06, relative to the CS− which did not survive a Sidak correction for multiple comparisons, β = −.250, R
2
= .062, F (1, 60) = 3.99, p = .050, BF10 = 1.36. A post-hoc multiple regression analysis with the final two CS+ extinction learning trials averaged and entered alongside IUS scores significantly predicted ratings to the first CS+ trial of the reinstatement phase, R
2
= .287, F (2, 59) = 11.89, p < .001. Here, threat expectancy ratings at the end of extinction learning (β = .520, t = 4.66, p < .001) and IUS scores (β = −.254, t = −2.27, p = .027) were both significant predictors in the model. These results suggest IU has a negative association with threat expectancy ratings following reinstatement (see Figure 2), and that this effect is driven by responding to the CS+, but not the CS−. Scatterplots of the correlations between the IUS total score and (a) the reinstatement index of differential responding, operationalised as the difference between the CS+ and CS− on the final two trials of the extinction phase subtracted from the CS+/− difference on the first trial of the reinstatement phase, (b) reinstatement to the CS+, operationalised as the final two CS+ trials of the extinction phase subtracted from the first CS+ trial of the reinstatement phase, and (c) reinstatement to the CS−, operationalised as the final two CS− trials of the extinction phase subtracted from the first CS- trial of the reinstatement phase. IUS: Intolerance of uncertainty scale.
Post-hoc analyses
To explore the negative relationship between IU and reinstatement effects, and to further assess the likelihood that these effects may be driven by reduced (or incomplete) extinction in participants with high IU, we split the sample into quartiles based on IUS scores and compared the lower (n = 17) and upper quartiles (n = 15) on pre- and post-reinstatement threat expectancy ratings (see Figure 3). Here, a 2 (group: lower quartile IU, upper quartile IU) × 2 (CS: CS+, CS−) × 2 (time: extinction trials 11 and 12, reinstatement trial 1) mixed models ANOVA was conducted. The ANOVA revealed a significant main effect of CS, F (1, 30) = 19.23, p < .001, ηp2 = .391, BF10 = 0.336, with greater threat expectancy ratings to the CS+ compared to the CS−. There was also a significant group × time interaction, F (1, 30) = 9.39, p = .005, ηp2 = .238, BFincl = 29.27. Sidak-corrected tests of simple main effects revealed that the lower quartile IU group showed a significant non-differential CS+/− increase in threat expectancy ratings from pre-to post-reinstatement (M
DIFF
= −17.95, t (16) = −2.22, p = .034, d = −0.62), whilst the upper quartile IU group showed a significant non-differential CS+/− decrease in threat expectancy ratings from pre-to post-reinstatement (M
DIFF
= 18.18, t (14) = 2.12, p = .043, d = 0.63). Further tests of simple main effects revealed significant differential CS+/− expectancy at the end of the extinction learning phase for both the lower quartile IU group (M
DIFF
= 38.47, t (16) = 3.61, p = .002, d = 0.91) and the upper quartile IU group (M
DIFF
= 29.96, t (14) = 2.47, p = .016, d = 0.64) suggesting that neither group experienced complete extinction. Finally, there was no significant CS × time interaction, F (1, 30) = 2.65, p = .114, ηp2 = .081, BFincl = 0.378, group × CS × time interaction, F (1, 30) = 0.03, p = .859, ηp2 = .001, BFincl = 0.334, or group main effect, F (1, 30) = 1.52, p = .228, d = 0.44, BF10 = 3.277e-6. Interaction showing the change in threat expectancy ratings from pre-to post-reinstatement for participants classified as low and high IU. Here, the low IU group showed a significant non-differential CS+/− increase in threat expectancy ratings, whilst the high IU group showed a significant non-differential CS+/− decrease in expectancy ratings. IU: intolerance of uncertainty; CS: conditional stimulus; EXT: extinction learning phase; Reinstatement: reinstatement phase.
Discussion
In the current study we investigated intolerance of uncertainty (IU) as a predictor of the reinstatement of threat perception following unsignaled US exposure. Our hypothesis, that IU would be associated with significantly greater reinstatement of threat expectancy, was not supported. Rather, whilst we found that IU was a significant predictor of differential threat reinstatement, the effect was in the reverse direction, such that elevated IU was a predictor of reduced reinstatement of threat expectancy ratings to the CS+, but not the CS−.
In the current study, we found initial evidence for differential reinstatement of threat expectancy ratings. Simple effects analyses – and visual inspection of the results – show that differential CS+/− responding was smaller following reinstatement compared to the end of the extinction learning phase. These findings could be argued to provide support for previous research showing differential reinstatement (Norrholm et al., 2006), although this support is limited due to the lack of significant change independently for the CS+ and CS−. That is, further tests of simple main effects showed that there was no significant change in responding for either the CS+ or CS− independently. Significantly smaller differential CS+/− responding at reinstatement suggests that the unsignaled US produced significant uncertainty by challenging prior understanding of the CS+/US contingency (Haaker et al., 2014). Notably, this uncertainty may result in a general increase in reactivity, as well as enhanced reactivity to the conditioned context (Haaker et al., 2013; Lonsdorf et al., 2014). Consistent with this explanation, we expected that reinstatement effects would be predicted by increased IU, as found by Lucas et al. (2018), however, this was not the case.
Regression analyses on the full sample (minus exclusions) found a significant, but negative, relationship between IU and differential threat reinstatement, which was driven by responses to the CS+, rather than the CS−. In further examining effects specifically at high and low levels of IU, exploratory analyses showed that higher IU was associated with reduced threat expectancy (i.e. certainty in receiving a shock) from the end of extinction to the start of the reinstatement phase across both CS+ and CS−, whilst lower IU was associated with increased threat expectancy from the end of extinction to the start of the reinstatement phase across both CS+ and CS−. Interestingly, the low IU group showed larger differential CS+/− responding at the end of extinction learning compared to the high IU group, with moderate to large effect sizes in both groups. Whilst the high IU group showed higher threat expectancy to the CS+ and CS− at the end of extinction, differential responding here is smaller. The differences in the findings may be due to statistical power as the regression analyses were based on the full sample, whereas the group-based analyses were based on a sub-sample. Despite the findings being slightly different, both results suggest IU-related effects from extinction to reinstatement. Notably, these findings were at odds with our initial hypothesis that higher IU would be associated with greater threat expectancy ratings to the CS+ versus CS− during reinstatement. There are two potential explanations for these results. Firstly, it has been frequently identified that high IU participants show significantly less extinction than low IU participants (Morriss, 2019; Morriss et al., 2016), and it is therefore plausible that these findings were driven by a reduced level of extinction, rather than reduced reinstatement per se. That is the reinstatement of threat perception is predicated on a prior actual extinction of threat responses, and if those high in IU did not previously extinguish learned threat contingencies then they may be unlikely to show reinstatement effects operationalised as a further increase in threat expectancy. Indeed, further exploratory simple effects suggested that both the low and high IU groups were experiencing significant ongoing differential CS+/− responding at the end of the extinction learning phase. These findings suggest that an unsignaled US experience may cause an increase or decrease in differential responding as a function of both (a) individual differences in IU, and (b) the level of prior extinction learning. Secondly, it is possible that these surprising findings are related to the wording of the threat expectancy ratings (e.g. certainty of receiving a shock). Individuals with high IU, relative to low IU, may have felt more uncertain about the shift in contingencies at the end of extinction to the start of reinstatement (e.g. were providing ratings in the middle of the scale for both CS+ and CS− during reinstatement).
In the present study, the mean and range of IU scores observed were comparable to that of undergraduate and community samples. A number of individuals classed as high IU scored >40 on the IUS-12, which is similar to that of clinically anxious and depressed samples (Carleton et al., 2012; Khawaja & Yu, 2010). Thus, the sample from the current study reflects a mixture of young adult healthy and clinical populations. However, further examination of the role of IU in reinstatement is required in both healthy and clinical populations.
There are several limitations in the current study. Firstly, the current study did not include psychophysiological indices of CS responding. Previous research finding non-differential, non-selective, and differential evidence for reinstatement has done so with skin conductance responding (Kull et al., 2012; Lucas et al., 2018) and fear-potentiated startle (Norrholm et al., 2006). Whilst previous research has also found support for reinstatement with self-reported threat expectancy ratings (Dirikx et al., 2009; Hermans et al., 2005; Zuj et al., 2018), including psychophysiological measures of arousal in the current study would have produced a more comprehensive investigation of IU and reinstatement. Secondly, there were a large number of exclusions (n = 24) due to poor contingency awareness, where these participants reported greater threat expectancy to the CS− compared to the CS+ at the end of the threat conditioning phase. This was unexpected and is not thought to be due to the partial-continuous reinforcement schedule used during acquisition, which has been found to produce stronger, more extinction-resistant learning (Grady et al., 2016). Rather, the current wording of the threat expectancy scale (i.e. ‘Please indicate how certain you are that you will receive a shock after this circle?’) likely did not tap into threat expectancy per se, but certainty of experiencing the US. Whilst this is not uncommon in conditioning and extinction research (e.g. Lommen et al., 2013; Zuj et al., 2018, 2020), future research focussing specifically on expectancy of the US may reveal different effects. Finally, the current study did not include a control group of participants that did not receive the reinstatement stimuli. This is an important procedure to allow for further clarification of reinstatement effects (Haaker et al., 2014) as previous research has found reinstatement effects in control groups that did not receive reinstatement stimuli (Kull et al., 2012).
The current study used the partial-continuous reinforcement schedule from Grady et al. (2016), which was argued to produce extinction-resistant conditioned associations. As a result, this may have impacted upon the subsequent reinstatement effects in the current study and future research would benefit from using a different reinforcement schedule. Indeed, a systematic investigation of the effect of reinforcement rate during threat acquisition on threat reinstatement would be of significant benefit to reinstatement research moving forward. Future reinstatement research may also benefit from a variable number of trials during the extinction learning phase such that reinstatement does not begin until CS+ responding is extinguished to levels equivalent to CS− responses (Sjouwerman & Lonsdorf, 2020). Although there is evidence that individuals high in IU show greater differential conditioned responses late in extinction learning (Morriss et al., 2016), there is also evidence of greater retention of safety associations in high IU participants when extended extinction sessions are used (Morriss et al., 2020).
In conclusion, we found inconclusive evidence for differential reinstatement of threat expectancy ratings following unsignaled (reinstatement) delivery of an aversive US. Whilst we hypothesised that higher IU would be associated with greater differential threat reinstatement, this was not supported. Instead, we found that higher IU was associated with reduced threat expectancy (i.e. certainty in receiving a shock) to the CS+ from the end of extinction to the start of the reinstatement phase. Notably, when examining individuals with extreme low and high IU scores, those with high IU scores showed reduced threat expectancy (i.e. certainty of receiving a shock) from extinction to reinstatement overall. Such results suggest that individual differences in IU likely play an important role in reinstatement, although further research is required to understand how IU differentially impacts threat and safety learning mechanisms during reinstatement.
Supplemental Material
Supplemental Material – Intolerance of uncertainty predicts altered threat expectancy ratings during reinstatement
Supplemental Material for Intolerance of uncertainty predicts altered threat expectancy ratings during reinstatement by Daniel V. Zuj, Simon Dymond, Stuart B. Murray, Shaima Thomas and Jayne Morriss in Journal of Experimental Psychopathology
Footnotes
Author contributions
All authors contributed to the study conception and design. Material preparation and data collection were performed by Daniel Zuj and Shaima Thomas. Data analyses were performed by Daniel Zuj and Jayne Morriss. The first draft of the manuscript was written by Daniel Zuj and all authors commented on subsequent versions of the manuscript. All authors read and approved the final manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Data statement
For access to the data please contact Daniel Zuj:
Supplemental Material
Supplemental material for this article is available online.
Note
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.