
Abstract
Background:
In this study, we aimed to investigate the effect of iodide intake adjustment, 1,25(OH)2D3 supplementation, or both, on the thyroid gland of rat offspring.
Methods:
The offspring of female rats administered 100 times the normal dose of iodide (100 HI; 750 μg/d) during pregnancy and lactation were divided into four different treatment groups. They were either having their iodide intake adjusted from 100 HI to normal iodide intake (7.5 μg/day) or supplemented with 25-hydroxy vitamin D3 [1,25(OH)2D3; 5 μg·kg−1·day−1], or both, for 4 weeks. Thyroid sodium pertechnetate (Na99mTcO4) uptake percentages were measured using single-photon emission computed tomography, while serum levels of free triiodothyronine (FT3), free thyroxine (FT4), thyroglobulin antibody (TgAb), thyroid peroxidase antibody (TPOAb), and vitamin D3 (VD3) were monitored using enzyme-linked immunosorbent assay. The messenger ribonucleic acid expression of interleukin (IL)-17A, interferon gamma (IFN-γ), and IL-10 in the thyroid gland was measured using quantitative real-time polymerase chain reaction, while the protein expression of thyroid-hormone-receptor α1 (TRα1) and thyroid-hormone-receptor β1 (TRβ1) in the thyroid gland was detected using Western blotting. Haematoxylin and eosin (H & E) and immunofluorescence staining were also used to assess thyroid follicular structure and lymphocytic infiltration in the thyroid glands.
Results:
The immunofluorescence staining showed CD4+ co-localized with TRβ1 or the vitamin D receptor in thyroid gland cells of rats that were continuously treated with 100 HI. Following iodide adjustment, 1,25(OH)2D3 supplementation, or both, an increase in serum levels of FT3, free thyroxine, and VD3, protein expression of TRα1 and TRβ1 in the thyroid gland cells, and Na99mTcO4 thyroid uptake percentages was observed. The mRNA expression levels of IL-17A and IFN-γ, decreased, while the mRNA expression levels of IL-10 increased in the thyroid cells of each treatment group, except the group with continuous 100 HI intake.
Conclusion:
Iodide adjustment, 1,25(OH)2D3 supplementation, or both may increase the serum levels of FT3, FT4, and VD3, as well as the protein expression levels of TRα1 and TRβ1, in thyroid cells. In addition, iodide adjustment, 1,25(OH)2D3 supplementation, or both, may potentially reverse the imbalance in pro-inflammatory and anti-inflammatory cytokines (IL-17A, IFN-γ, and IL-10) caused by 100 HI, which may be beneficial in improving Na99mTcO4 thyroid uptake percentages.
Introduction
Epidemiological studies have indicated that excess iodine consumption may lead to hypothyroidism, 1 hyperthyroidism, 2 and autoimmune thyroid diseases. 3 Adequate iodine consumption during pregnancy and lactation guarantees the maintenance of normal maternal and fetal thyroid function. Serrano-Nascimento et al. investigated the effects of distilled water supplemented with five times higher-than-normal iodide concentration (sodium iodide, NaI) during the pregnancy and lactation period of female rats. The results showed decreased circulating levels of free triiodothyronine
(FT3) and free thyroxine (FT4) in offspring. 4 Previously, our group investigated the effects of 100 times higher-than-normal iodide intake (potassium iodide, KI) in female rats during the pregnancy and lactation period. This resulted in decreased FT3, FT4 and increased thyroid antibodies (TPOAb and TgAb) serum levels and T-cell lymphocytic infiltration in the thyroid of rat offspring that were continuously fed 100 HI (100 times the normal iodide dose) from weaning (postnatal day 21, PN21) until postnatal day 180 (PN180). 5 Therefore, high iodide supplementation during pregnancy and lactation periods can alter thyroid function in female rats and their offspring. However, the effect of iodide adjustment and with 25-hydroxy vitamin D3 [1,25(OH)2D3] supplementation on rat offspring, thyroid functions remains unclear.
The involvement of vitamin D in the regulation of the immune system has been emphasized in recent years, 6 as the well-established function of vitamin D is to regulate calcium homeostasis. Epidemiological and animal-model studies of human diseases show evidence that vitamin D deficiency is a predisposing condition for autoimmune diseases. 7 Some studies have also demonstrated the inhibitory effect of 1,25(OH)2D3 supplementation on the development of autoimmune diseases, such as inflammatory bowel disease, experimental autoimmune encephalomyelitis, and experimental autoimmune uveitis.8–10 Although Chen et al. reported that 1,25(OH)2D3 supplementation is effective for thyroglobulin (Tg)-induced autoimmune thyroiditis, 11 the effect of 1,25(OH)2D3 supplementation on high-iodide-induced thyroid diseases remains elusive. Therefore, in this study we aimed to investigate the effect of iodide intake adjustment, 1,25(OH)2D3 supplementation, or both, on the structure and function of the thyroid gland in the offspring of rats exposed to 100 HI.
Methods and materials
Animals and administration
Healthy, adult (6-weeks old) Wistar rats were obtained from the Experimental Animal Center of the Military Medical Science Academy of China. The animals were housed in clean polypropylene cages and maintained at a temperature of 22 ± 1°C in the specific pathogen-free level of the Experimental Animal Center of Tianjin Medical University. A constant light:dark cycle (12:12 h) was maintained throughout the study period. After 1 week of adaptation, female rats were mated with male rats (1:1). Gestation was confirmed by a positive vaginal plug or the presence of sperm in the vaginal smear of the female rats. The pregnant rats were randomly assigned to two groups: NI (normal iodide intake, n = 6) and 100 HI (100 times higher-than-normal iodide intake, n = 12). The rats in the NI group received dietary feed containing iodide (7.5 μg/day), in addition to orally administered deionized water. Rats in the 100 HI group received deionized water containing KI (24,750 μg/l) and dietary iodide; therefore, the intake of iodide was 750 μg/day. 12
The offspring were continuously administered KI from weaning (PN21) to PN90. After PN90, the NI rats were held as the control group (group 1), and the rats with 100 HI were randomly divided into four treatment groups: adjustment from 100 HI to NI administration (group 2), adjustment from 100 HI to NI administration + 1,25(OH)2D3 supplementation (group 3), continued 100 HI administration + 1,25(OH)2D3 supplementation (group 4), continued 100 HI administration (group 5). The rats in groups 3 and 4 were supplemented with 1,25(OH)2D3 (Medchem Express, Monmouth Junction, USA) by gavage (5 μg·kg−1·day−1). The treatment was carried out for 4 weeks (Figure 1). The vitamin D supplementation guidelines recommend doses ranging between 400 IU/day and 2000 IU/day to prevent or correct vitamin D deficiency depending on the age, body weight, ethnic origin, presence of certain diseases, and pharmaceutical consumption. 13 Based on the normalization method of body surface area, the conversion factor from the dosage of humans to rats was 6.17, 14 and the recommended doses ranging between 400 IU/day and 2000 IU/day are equivalent to 1.6–8.1 µg/day in rats. A dose of 5 µg·kg−1·day−1 of 1,25(OH)2D3 was used in our experiments, which is within the normalized range.

Experimental design.
All procedures were approved by the Institutional Animal Care and Use Committee of Tianjin Medical University (no. TMUaMEC 2016054), were in accordance with the guidelines of the Committee for Humane Animal Treatment, and complied with the relevant legislation.
Measurement of urinary iodine concentration
Urine samples were collected 24 h before the rats were sacrificed. Urinary iodine concentration
Single-photon emission computed tomography
The rats were given a tail vein injection of 500 μCi sodium pertechnetate (Na99mTcO4) and were anesthetized after 20 min using 10% chloral hydrate (0.3 ml/100 g of body weight). The anesthetized rats were maintained in a prone position on an animal bed. Animal imaging was performed using a single-photon emission computed tomography (SPECT) system (GE NM Infinia VC HE4, Milwaukee, WI, USA). For quantitative evaluation, irregular regions of interest were drawn over the thyroid and armpit area to calculate the uptake of Na99mTcO4 in the thyroid gland:

Thyroid function measurements
At the end of treatment, blood samples were collected from the orbital sinus and centrifuged for 10 min at 1500 rpm to obtain the serum. The levels of FT3, FT4, and vitamin D3 (VD3) (Meilian Biological Technology, Shanghai, China), as well as levels of TgAb and TPOAb (Mybiosourc, San Diego, CA, USA), were determined using rat-specific enzyme-linked immunosorbent assay kits.
Haematoxylin and eosin (H & E) and immunofluorescence staining
Some thyroid gland sections were stained with haematoxylin and eosin (H & E), while the remaining sections were incubated with the primary anti-thyroid-hormone-receptor β1 (TRβ1) antibody (1:200) or anti-vitamin D receptor (VDR) antibody (1:200; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), and anti-CD4 antibody (1:200; Abcam, Cambridge, MA, USA), at 4°C overnight. The following day, the sections were incubated with secondary antibodies in the dark for 30 min at 37°C. The nuclei were stained with 100 μl Hoechst 33258, and the sections were visualized using a Zeiss LSM 510 laser confocal microscope (Carl Zeiss Microscopy GmbH, Germany) for immunofluorescence analysis.
Western blotting
Protein samples were separated using 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto a polyvinylidene fluoride (PVDF) membrane. The blots were incubated with primary antibodies against thyroid-hormone-receptor α1 (TRα1) or TRβ1 (1:1000; Abcam, Cambridge, MA, USA) at 4°C overnight. The PVDF membrane was also incubated with secondary antibodies for 1 h. The proteins were visualized using chemiluminescence.
RNA extraction and qRT-PCR
Total ribonucleic acid (RNA) was extracted from the thyroid gland using TRIzol reagent (Life Technologies, California, USA). The complementary DNA (cDNA) was synthesized using a reverse-transcription kit (CWBio, Peking, China). The relative messenger ribonucleic acid (mRNA) levels were normalized to the internal control glyceraldehyde 3-phosphate dehydrogenase (GAPDH) using the 2−ΔΔCt method and SYBR Green (CWBIO). The following polymerase chain reaction (PCR) primer strands were used:
Interleukin (IL)-17A forward primer: 5′CGCCGAGGCCAATAACTTTC 3′
IL-17A reverse primer: 5′GGTTGAGGTAGTCTGAGGGC 3′
Interferon (IFN)-γ forward primer: 5′CGTCTTGGTTTTGCAGCTCT 3′
IFN-γ reverse primer: 5′CGTCCTTTTGCCAGTTCCTC 3′
IL-10 forward primer: 5′CCTGGTAGAAGTGATGCCCC 3′
IL-10 reverse primer: 5′TGCCGGGTGGTTCAATTTTT 3′
GAPDH forward primer: 5′CATGGCCTTCCGTGTTCCTA 3′
GAPDH reverse primer: 5′ATGCCTGCTTCACCACCTTCT 3′
Statistical analysis
Groups 1 and 5 were compared using the independent-samples t test. Groups 2, 3, 4, and 5 were compared using two-way analysis of variance. To control for distribution skewedness, the median was used to describe the central tendency of the UIC. Differences among groups were evaluated using the non-parametric Kruskal–Wallis test, while the individual groups were compared with the control group using the Nemenyi post hoc test. All other quantitative data are expressed as the mean ± standard deviation. Differences in values were considered statistically significant at p < 0.05.
Results
Body weight and median UIC alteration following iodide adjustment and/or 1,25(OH)2D3 supplementation for 4 weeks
Before the rats were sacrificed, their body weight per group were as follows: 349 ± 73.58 g (group 1, n = 6), 309.64 ± 67.87 g (group 2, n = 6), 313.34 ± 66.48 g (group 3, n = 6), 324.35 ± 94.47 g (group 4, n = 6), 311.13 ± 77.61 g (group 5, n = 6). Furthermore, there was no significant difference in body weight among the five groups (p > 0.05). The median UIC in group 5 (continued 100 HI administration) was approximately 98 times higher than that of group 1 (control; p < 0.05). The median UIC was approximately 11 and 12 times lower in group 2 (adjustment from 100 HI to NI administration) and group 3 [adjustment from 100 HI to NI administration + 1,25(OH)2D3 supplementation] (p < 0.05), respectively, than in group 5 (continued 100 HI administration), while there was no significant difference when compared with group 4 [continued 100 HI administration + 1,25(OH)2D3 supplementation] median UIC (p > 0.05; Table 1).
Median urinary iodine concentrations (UIC) in different treatment groups.

1,25(OH)2D3, 25-hydroxy vitamin D3; HI, 100 times the normal dose of iodide; NI, normal iodide intake.
Iodide adjustment and/or 1,25(OH)2D3 supplementation improved thyroid uptake percentages in SPECT
Compared with group 1 (control), the thyroid uptake percentages in group 5 (continued 100 HI administration) were decreased significantly (p < 0.05). Compared with group 5 (continued 100 HI administration), the thyroid uptake percentages were increased significantly in group 2 (adjustment from 100 HI to NI administration), group 3 [adjustment from 100 HI to NI administration + 1,25(OH)2D3 supplementation], and group 4 [continued 100 HI administration + 1,25(OH)2D3 supplementation] (p < 0.05) after 4 weeks of treatment (Figure 2).

SPECT imaging Na99mTcO4 thyroid uptake percentages in different treatment groups.
Effect of iodide adjustment and/or 1,25(OH)2D3 supplementation on thyroid function and VD3 levels
Compared with group 1 (control), the FT3, FT4, and VD3 levels were decreased, and the TPOAb and TgAb levels were significantly increased (p < 0.05) in group 5 (continued 100 HI administration). Compared with group 5 (continued 100 HI administration), the FT3, FT4, and VD3 levels were improved significantly in group 2 (adjustment from 100 HI to NI administration), group 3 [adjustment from 100 HI to NI administration + 1,25(OH)2D3 supplementation], and group 4 [continued 100 HI administration + 1,25(OH)2D3 supplementation] (p < 0.05; Table 2).
Thyroid hormone, autoantibody and VD3 levels in different treatment groups.
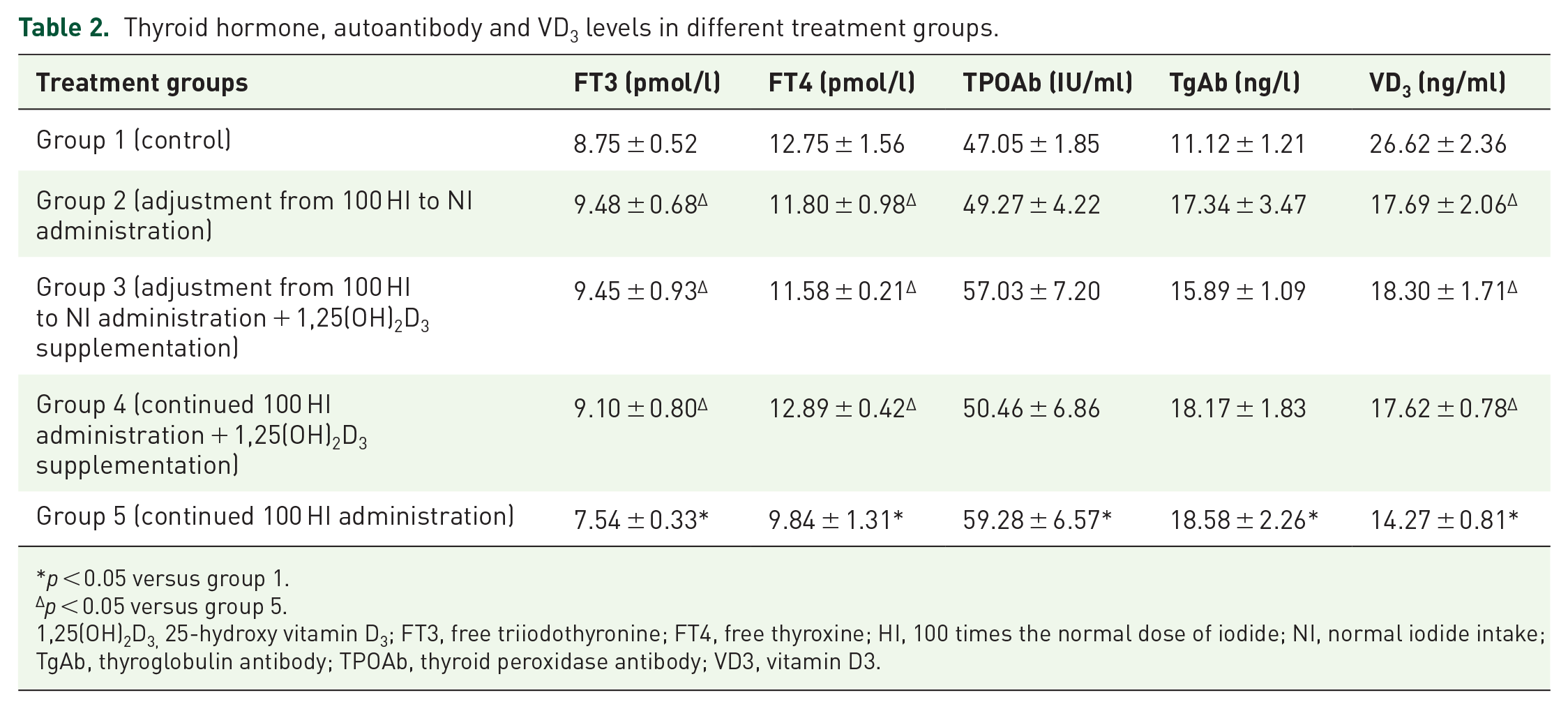
p < 0.05 versus group 1.
p < 0.05 versus group 5.
1,25(OH)2D3, 25-hydroxy vitamin D3; FT3, free triiodothyronine; FT4, free thyroxine; HI, 100 times the normal dose of iodide; NI, normal iodide intake; TgAb, thyroglobulin antibody; TPOAb, thyroid peroxidase antibody; VD3, vitamin D3.
Continued 100 HI administration resulted in lymphocytic infiltration in the thyroid gland, and CD4+ was co-localized with TRβ1 or VDR in the infiltrated cells
HE and immunofluorescence staining showed a relatively intact structure of thyroid follicles in group 1 (control). In group 5 (continued 100 HI administration), lymphocytic infiltration was observed in the thyroid follicular cavity and around the follicles; thyroid follicular epithelial cells were characterized by an enlarged shape, and the nuclei were hyperchromatic and had prominent nucleoli. Immunofluorescence staining showed positive staining for CD4+ (red) co-localized with TRβ1 or VDR (green) and Hoechst (blue) in the infiltrated cells in group 5 (continued 100 HI administration) (Figure 3).

Histological analysis of the thyroid gland using haematoxylin and eosin (H & E) and immunofluorescence staining.
Iodide adjustment and/or 1,25(OH)2D3 supplementation improved the expression of TRα1 and TRβ1 in thyroid cells
The expression of TRα1 and TRβ1 was decreased in group 5 (continued 100 HI administration), compared to the expression observed in group 1 (control) (p < 0.05). Compared with group 5 (continued 100 HI administration), the expression of TRα1 and TRβ1 was increased significantly in group 2 (adjustment from 100 HI to NI administration), group 3 [adjustment from 100 HI to NI administration + 1,25(OH)2D3 supplementation], and group 4 [continued 100 HI administration + 1,25(OH)2D3 supplementation] (p < 0.05; Figure 4).

The protein levels of TRα1 and TRβ1 in the thyroid analyzed by Western blotting (n = 6 for each group).
Iodide adjustment and/or 1,25(OH)2D3 supplementation decreased the mRNA expression of IL-17A and IFN-γ, while IL-10 increased in thyroid cells
Compared with group 1 (control), the mRNA expression of IL-17A and IFN-γ was increased significantly, while the mRNA expression of IL-10 was decreased significantly in group 5 (continued 100 HI administration; p < 0.05). Compared with group 5 (continued 100 HI administration), the mRNA expression of IL-17A and IFN-γ were decreased significantly, while IL-10 mRNA expression was increased significantly in group 2 (adjustment from 100 HI to NI administration), group 3 [adjustment from 100 HI to NI administration + 1,25(OH)2D3 supplementation], and group 4 [continued 100 HI administration + 1,25(OH)2D3 supplementation] (p < 0.05; Figure 5).

Effects of different treatments on the mRNA expression of IL-17A, IFN-γ, and IL-10 in the thyroid gland measured by qRT-PCR.
Discussion
Following SPECT analysis, we showed that the uptake of Na99mTcO4 by thyroid cells decreased significantly in group 5 (continued 100 HI administration). 15 After 4 weeks of adjusted iodide intake, 1,25(OH)2D3 supplementation, or both, the results showed significant improvement in Na99mTcO4 thyroid uptake percentages. Franken et al. studied 6–8-week-old mice injected intraperitoneally with approximately 250–300 mBq of Na99mTcO4 and 2 mg NaI after 20 min, which resulted in a decrease in thyroid uptake percentages. 16 The US Institute of Medicine, World Health Organization, United Nations Children’s Fund, and the International Council for the Control of Iodine Deficiency Disorders recommend a daily iodine intake of 150 μg in adults. 17 The administration of 100 HI in a rat is the equivalent of 16.2 times the normal iodine intake recommended for a human being. Epidemiological studies indicate that the mean iodine intake from drinking water in some counties (cities, districts) within China is 1073.5 μg/day, 18 which is equivalent to seven times that of the normal human iodine intake. In populations that consume seaweed, such as the Japanese, the mean iodine intake is 1.5 mg/day, 19 which is equivalent to 10 times that of the normal human iodide daily intake. During the metabolism of amiodarone, a benzofuran derivative used for long-term treatment of cardiac arrhythmias, approximately 9 mg of iodine is released. This is equivalent to 60 times that of the normal human iodide daily intake. 20
This study demonstrated that the mRNA expression of IL-17A and IFN-γ increased, while the mRNA expression of IL-10 decreased in the thyroid gland cells of group 5 rats (continued 100 HI administration). IL-17 and IFN-γ are mainly secreted by T-helper 1 (Th1) or Th17 cells, which play key roles in autoimmune diseases, such as multiple sclerosis and ulcerative colitis. 21 Regulatory T cells are a major source of IL-10 and have immunosuppressive and anti-inflammatory properties. 22
Moreover, the FT3 level and the expression of TRα1 and TRβ1 in the thyroid decreased in the 100 HI continuous administration group of the rat offspring. Following the iodide intake adjustment, 1,25(OH)2D3 supplementation, or both, we observed an improvement in the expression levels of these molecules. Many thyroid hormone (T3) functions are mediated by TRs. The binding of T3 to its receptor plays key physiological roles in the regulation of development, growth, and metabolism. 23 In addition, positive staining of CD4+ co-localized with TRβ1 in the infiltrated cells was also observed. TRβ1 is a T3-dependent transcription factor; therefore, the decreased FT3 induced by 100 HI administration may interact with TRβ1 in CD4+ T cells, thus affecting the function of CD4+ T cells.
A significant decrease in serum level of VD3 was observed following 100 HI administration. 1,25(OH)2D3 supplementation helps reverse the changes in mRNA expression of IL-17A, IFN-γ, and IL-10 induced by 100 HI administration. 1,25(OH)2D3 is the active form of vitamin D, which exerts its actions by binding to VDRs. 24 As demonstrated by immunofluorescence staining, CD4+ was co-localized with VDRs in the thyroid-gland-infiltrated lymphocytes. Consistent with our results, Chang et al. reported that 1,25(OH)2D3 can protect against myelin oligodendrocyte-glycoprotein-induced experimental autoimmune encephalomyelitis through VDR signaling, by suppressing the expression of IL-17, while enhancing the expression of IL-10. 25 It has been demonstrated that 50% of the candidate risk gene orthologs changed their expression in CD4+ T cells upon vitamin D supplementation. 6 Vitamin D can also reduce the concentration of enzymes involved in maintaining deoxyribonucleic acid methylation, 6 the activation of nuclear factor κB, and the release of inflammatory cytokines. 26
The present study demonstrates that treatment with 1,25(OH)2D3 preserves thyroid function by increasing HI-induced low levels of FT3. In animal studies, doses from 50 ng/day (2 IU/day) to 30 µg·kg−1·day−1 (1200 IU·kg−1·day−1) have been reported, all of which were effective (Table 3).25–32 Alrefaie et al. reported that following 10 weeks of oral vitamin D3 supplementation (500 IU·kg−1·day−1) in diabetic adult male rats, the levels of FT3 and FT4 returned towards normal levels, and this effect was not observed in untreated diabetic rats. 33 In addition, although 1,25(OH)2D3 treatment did not significantly change the levels of thyroid autoantibodies, a slight decrease was observed in TPOAb and TgAb levels. Zhang et al. measured the serum levels of 1,25(OH)2D3 and thyroid autoantibodies in 1424 healthy Chinese adults without a history of thyroid disease. The results showed no correlation between vitamin D status and the presence of thyroid autoantibodies after controlling for influential factors such as age, sex, body mass index, and smoking status. 34 Goswami et al. measured the serum levels of TPOAb and 1,25(OH)2D3 in 642 healthy subjects in India. The results indicated a weak inverse correlation between the serum 1,25(OH)2D3 values and TPOAb titres (r = −0.08; p = 0.04). 35
The dosages of vitamin D supplementation in experimental autoimmune diseases.
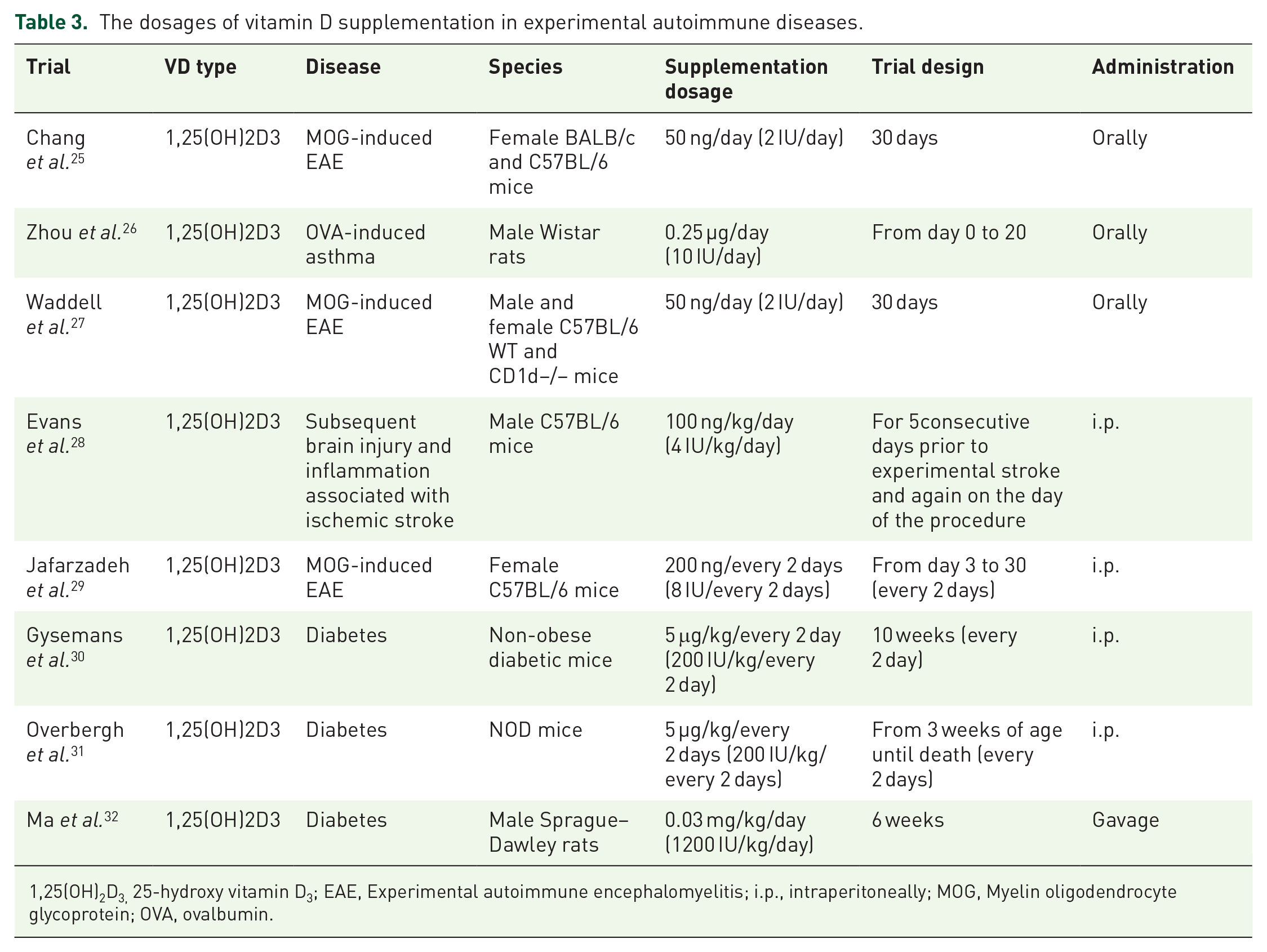
1,25(OH)2D3, 25-hydroxy vitamin D3; EAE, Experimental autoimmune encephalomyelitis; i.p., intraperitoneally; MOG, Myelin oligodendrocyte glycoprotein; OVA, ovalbumin.
In conclusion, the present study demonstrates that iodide intake adjustment, 1,25(OH)2D3 supplementation, or both, may improve the levels of serum FT3, FT4, and VD3, which may exert their actions on the infiltrated CD4+ cells in the thyroid gland by binding to the TR or VDR, respectively. Iodide intake adjustment or 1,25(OH)2D3 supplementation, or both can inhibit the expression of IL-17A and IFN-γ, which are pro-inflammatory cytokines, while enhancing the expression of IL-10, an anti-inflammatory cytokine. This protective effect may contribute to the improvement of Na99mTcO4 thyroid uptake percentages (Figure 6).
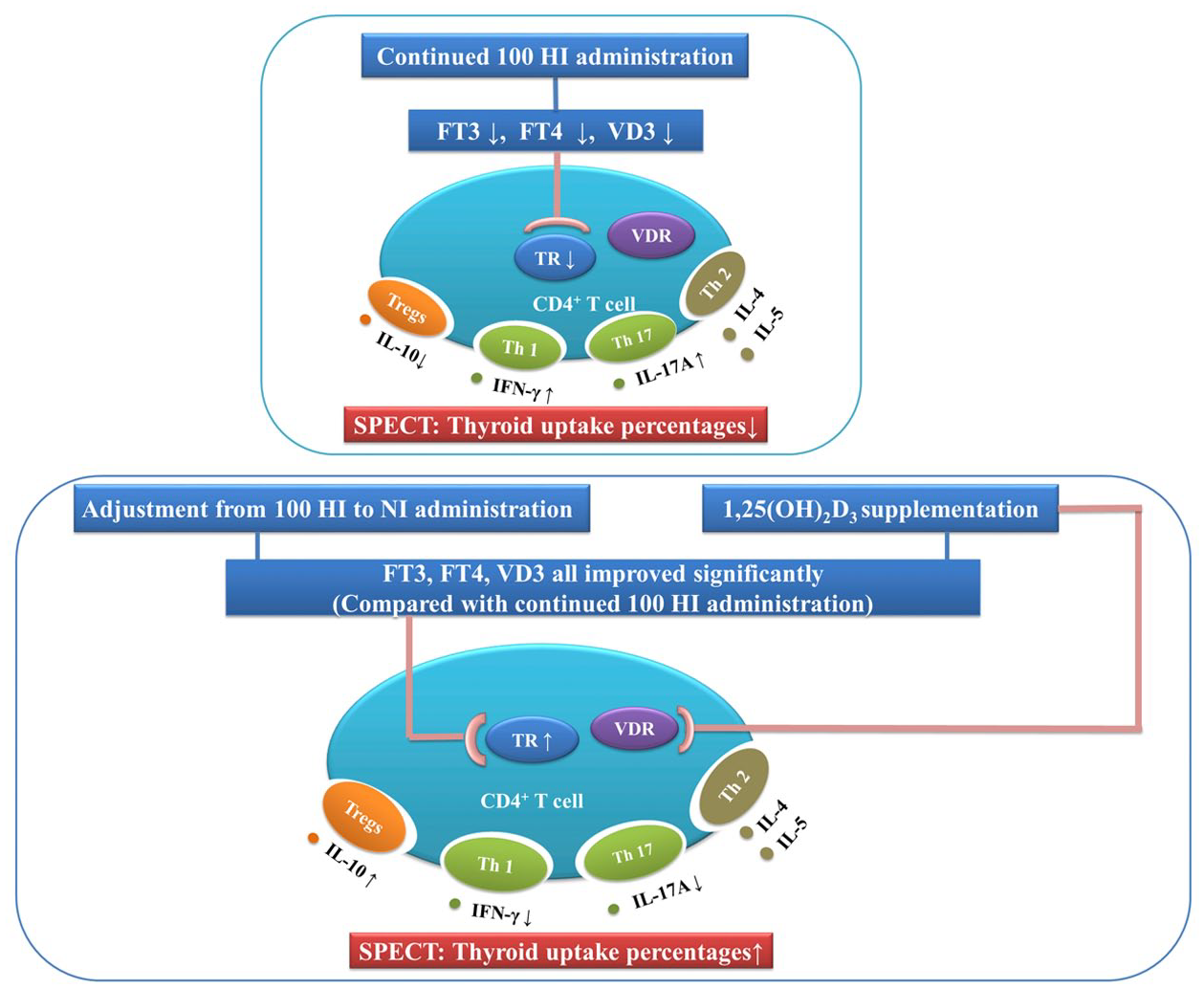
Proposed mechanisms in offspring rats. Improvement in FT3 and VD3 levels, expression of TRα1 and TRβ1, mRNA expression of IL-17A, IFN-γ, IL-10, and thyroid uptake percentages following iodide adjustment and/or 1,25(OH)2D3 supplementation.
Footnotes
Author contribution(s)
Conflict of interest statement
The authors declare that there is no conflict of interest.
Funding
The authors disclose receipt of the following financial support for the research, authorship, and/or publication of this article: This work is supported by the National Natural Science Foundation of China (nos. 81874257 and 81273009) as well as the Principal Investigator, Xiaomei Yao.